
Advances in Metabolic Engineering of Pichia pastoris Strains as Powerful Cell Factories
Abstract
:1. Introduction
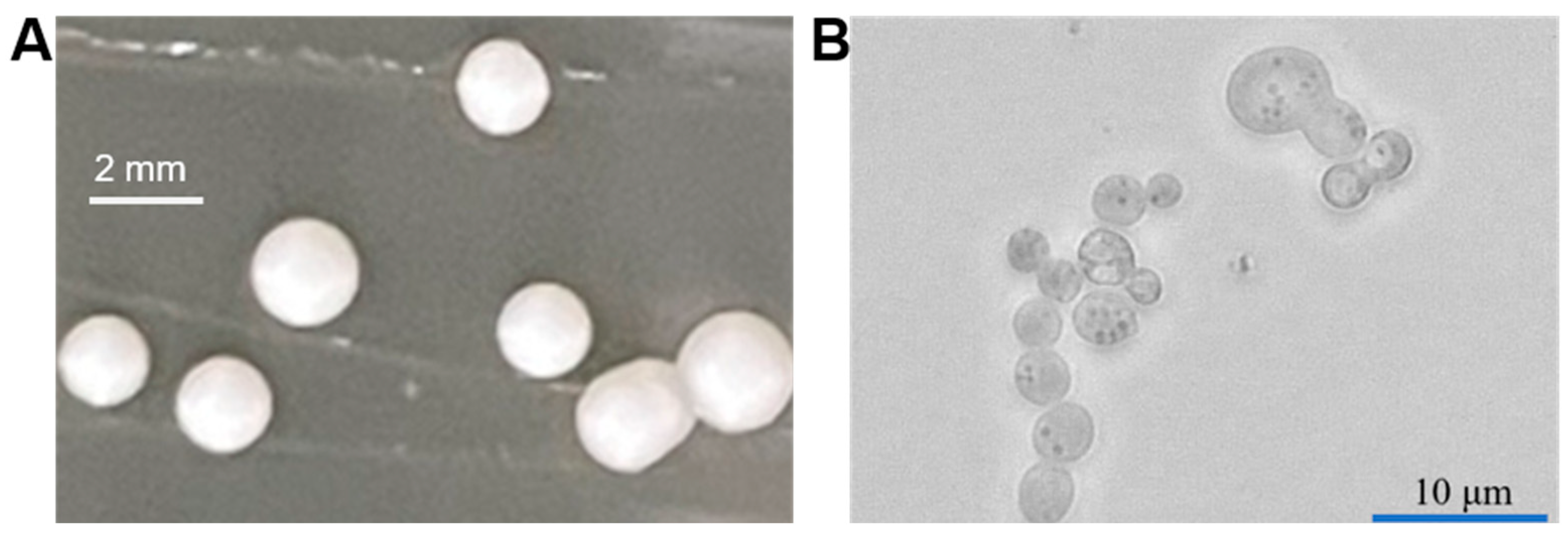
2. Physiological Characteristics of Pichia pastoris
3. The Pichia pastoris Expression System
3.1. Strains
3.2. Expression Vectors
3.3. Promoters
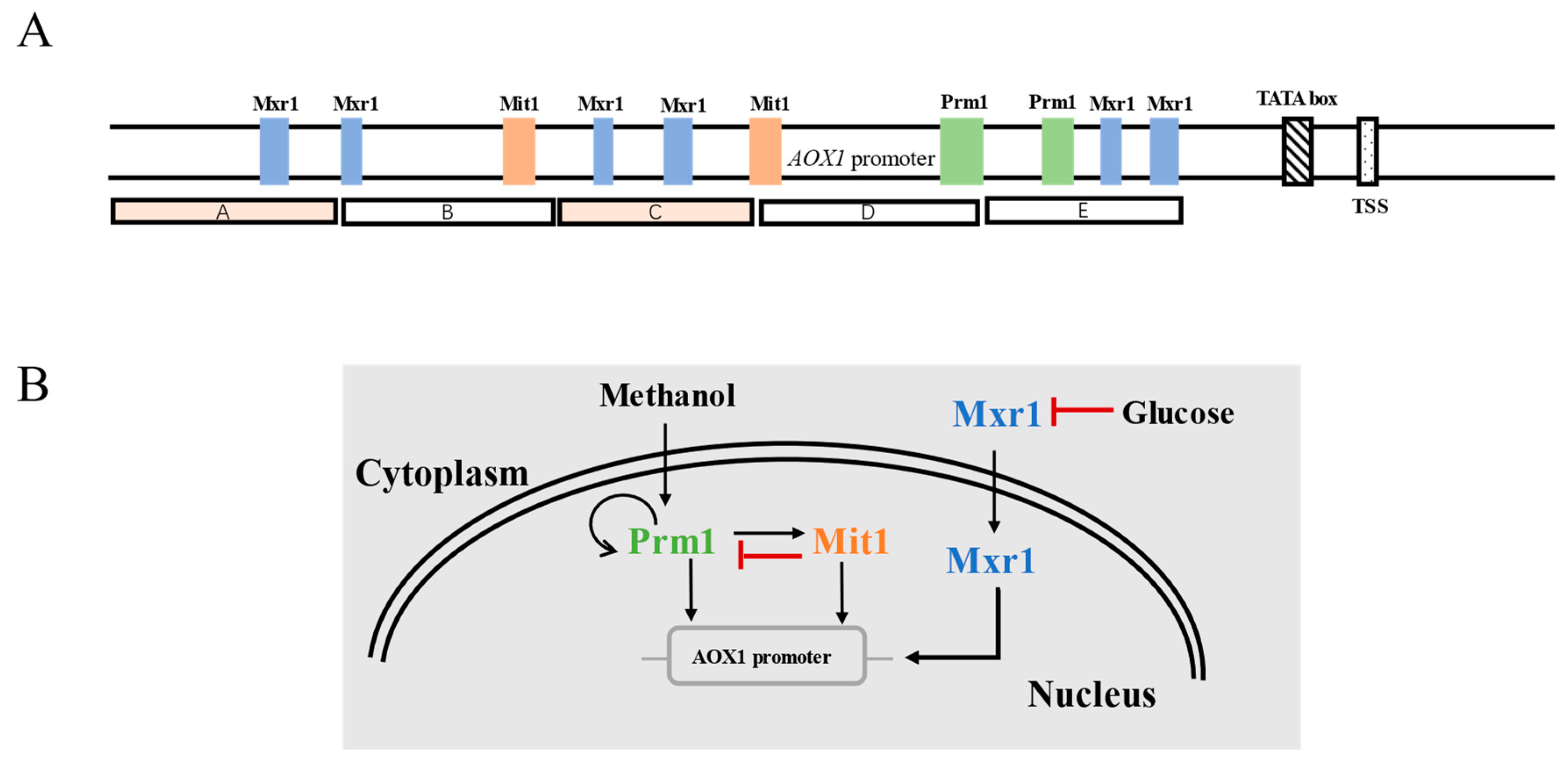
3.3.1. AOX1 Promoter
3.3.2. GAP Promoter
3.3.3. Other Promoters
3.3.4. Synthetic Core Promoter Engineering
3.4. Signal Peptides
3.5. CRISPR/Cas9 Genome Editing in Pichia pastoris
4. Adaptive Laboratory Evolution of Pichia pastoris
5. Practical Applications of Pichia pastoris as a Cell Factory
5.1. Recombinant Proteins
5.1.1. Nanobodies
5.1.2. Human Proteins
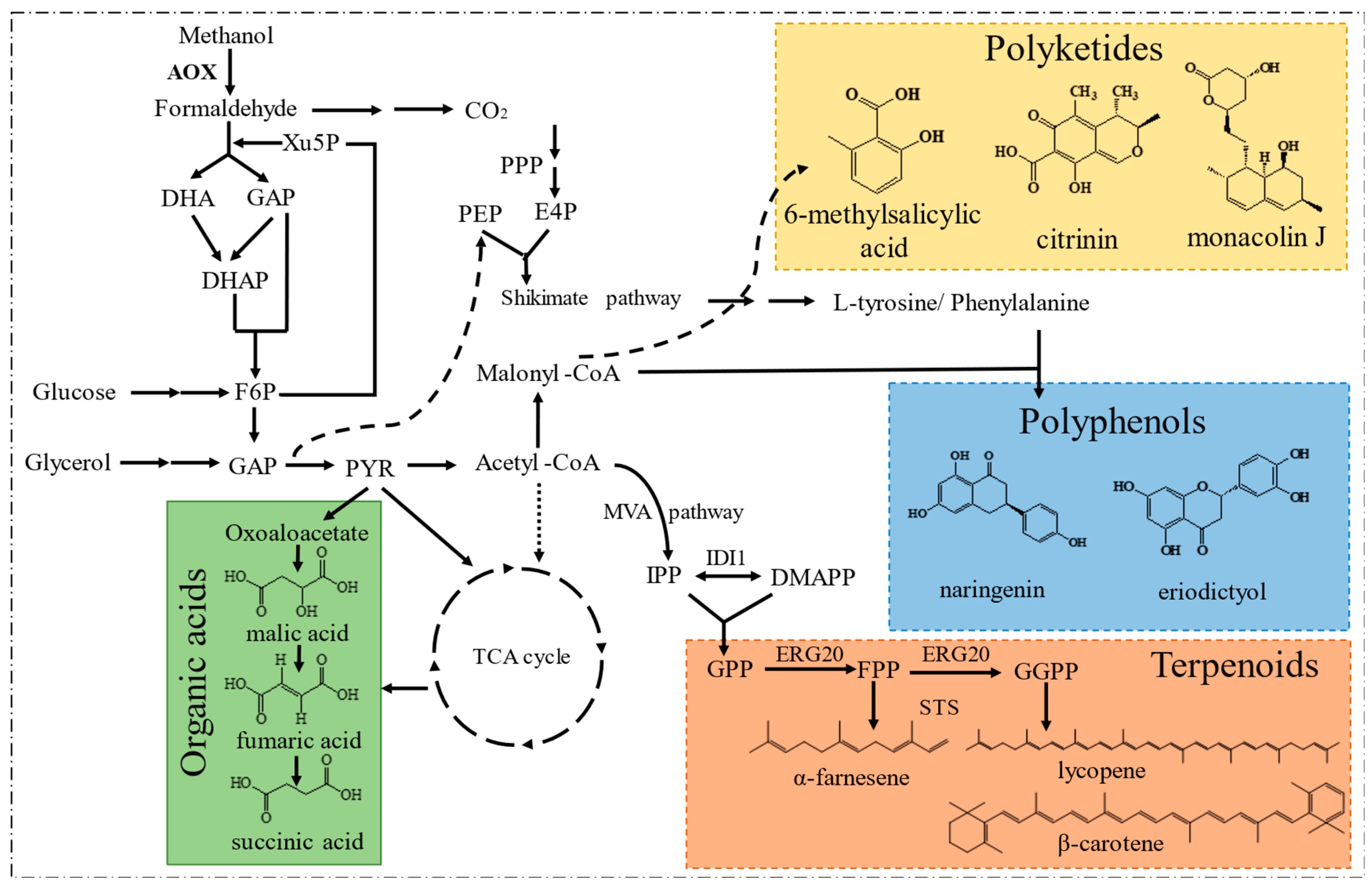
5.2. Value-Added Compounds
5.2.1. Terpenoids
5.2.2. Polysaccharides
5.2.3. Polyketides
5.2.4. Other Compounds
6. Conclusions and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kurtzman, C.P. Biotechnological strains of Komagataella (Pichia) pastoris are Komagataella phaffii as determined from multigene sequence analysis. J. Ind. Microbiol. Biotechnol. 2009, 36, 1435–1438. [Google Scholar] [CrossRef] [PubMed]
- Kulagina, N.; Besseau, S.; Godon, C.; Goldman, G.H.; Papon, N.; Courdavault, V. Yeasts as biopharmaceutical production platforms. Front. Fungal Biol. 2021, 2, 733492. [Google Scholar] [CrossRef]
- Cereghino, G.P.L.; Cereghino, J.L.; Ilgen, C.; Cregg, J.M. Production of recombinant proteins in fermenter cultures of the yeast Pichia pastoris. Curr. Opin. Biotechnol. 2002, 13, 329–332. [Google Scholar] [CrossRef] [PubMed]
- Cereghino, J.L.; Cregg, J.M. Heterologous protein expression in the methylotrophic yeast Pichia pastoris. FEMS Microbiol. Rev. 2000, 24, 45–66. [Google Scholar] [CrossRef] [PubMed]
- Hasslacher, M.; Schall, M.; Hayn, M.; Bona, R.; Rumbold, K.; Lückl, J.; Griengl, H.; Kohlwein, S.D.; Schwab, H. High-level intracellular expression of hydroxynitrile lyase from the tropical rubber tree Hevea brasiliensis in microbial hosts. Protein Expr. Purif. 1997, 11, 61–71. [Google Scholar] [CrossRef]
- Karbalaei, M.; Rezaee, S.A.; Farsiani, H. Pichia pastoris: A highly successful expression system for optimal synthesis of heterologous proteins. J. Cell Physiol. 2020, 235, 5867–5881. [Google Scholar] [CrossRef]
- Pan, Y.; Yang, J.; Wu, J.; Yang, L.; Fang, H. Current advances of Pichia pastoris as cell factories for production of recombinant proteins. Front. Microbiol. 2022, 13, 1059777. [Google Scholar] [CrossRef]
- Gao, J.; Jiang, L.; Lian, J. Development of synthetic biology tools to engineer Pichia pastoris as a chassis for the production of natural products. Synth. Syst. Biotechnol. 2021, 6, 110–119. [Google Scholar] [CrossRef]
- Grinna, L.S.; Tschopp, J.F. Size distribution and general structural features of N-linked oligosaccharides from the methylotrophic yeast, Pichia pastoris. Yeast 1989, 5, 107–115. [Google Scholar] [CrossRef]
- Zhu, T.; Zhao, T.; Bankefa, O.E.; Li, Y. Engineering unnatural methylotrophic cell factories for methanol-based biomanufacturing: Challenges and opportunities. Biotechnol. Adv. 2020, 39, 107467. [Google Scholar] [CrossRef]
- Cai, P.; Duan, X.; Wu, X.; Gao, L.; Ye, M.; Zhou, Y.J. Recombination machinery engineering facilitates metabolic engineering of the industrial yeast Pichia pastoris. Nucleic Acids Res. 2021, 49, 7791–7805. [Google Scholar] [CrossRef] [PubMed]
- Schwarzhans, J.-P.; Luttermann, T.; Geier, M.; Kalinowski, J.; Friehs, K. Towards systems metabolic engineering in Pichia pastoris. Biotechnol. Adv. 2017, 35, 681–710. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, A.; Pal, M.; Sharma, R.K. Pichia as yeast cell factory for production of industrially important bio-products: Current trends, challenges, and future prospects. J. Bioresour. Bioprod. 2023, 8, 108–124. [Google Scholar] [CrossRef]
- Batt, C.A. Pichia pastoris. In Encyclopedia of Food Microbiology; Academic Press: London, UK, 2014; pp. 42–46. [Google Scholar] [CrossRef]
- Heistinger, L.; Moser, J.; Tatto, N.E.; Valli, M.; Gasser, B.; Mattanovich, D. Identification and characterization of the Komagataella phaffii mating pheromone genes. FEMS Yeast Res. 2018, 18, foy051. [Google Scholar] [CrossRef] [PubMed]
- Dragosits, M.; Stadlmann, J.; Albiol, J.; Baumann, K.; Maurer, M.; Gasser, B.; Sauer, M.; Altmann, F.; Ferrer, P.; Mattanovich, D. The effect of temperature on the proteome of recombinant Pichia pastoris. J. Proteome Res. 2009, 8, 1380–1392. [Google Scholar] [CrossRef] [PubMed]
- Rosenbergová, Z.; Kántorová, K.; Šimkovič, M.; Breier, A.; Rebroš, M. Optimisation of recombinant myrosinase production in Pichia pastoris. Int. J. Mol. Sci. 2021, 22, 3677. [Google Scholar] [CrossRef] [PubMed]
- Ata, Ö.; Rebnegger, C.; Tatto, N.E.; Valli, M.; Mairinger, T.; Hann, S.; Steiger, M.G.; Çalık, P.; Mattanovich, D. A single Gal4-like transcription factor activates the Crabtree effect in Komagataella phaffii. Nat. Commun. 2018, 9, 4911. [Google Scholar] [CrossRef]
- Zhan, C.; Wang, S.; Sun, Y.; Dai, X.; Liu, X.; Harvey, L.; McNeil, B.; Yang, Y.; Bai, Z. The Pichia pastoris transmembrane protein GT1 is a glycerol transporter and relieves the repression of glycerol on AOX1 expression. FEMS Yeast Res. 2016, 16, fow033. [Google Scholar] [CrossRef]
- Liu, B.; Zhang, Y.; Zhang, X.; Yan, C.; Zhang, Y.; Xu, X.; Zhang, W. Discovery of a rhamnose utilization pathway and rhamnose-inducible promoters in Pichia pastoris. Sci. Rep. 2016, 6, 27352. [Google Scholar] [CrossRef]
- Liu, B.; Li, H.; Zhou, H.; Zhang, J. Enhancing xylanase expression by Komagataella phaffii by formate as carbon source and inducer. Appl. Microbiol. Biotechnol. 2022, 106, 7819–7829. [Google Scholar] [CrossRef]
- Gabaldón, T. Peroxisome diversity and evolution. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 765–773. [Google Scholar] [CrossRef] [PubMed]
- Vogl, T.; Glieder, A. Regulation of Pichia pastoris promoters and its consequences for protein production. New Biotechnol. 2013, 30, 385–404. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.-W.; Lu, X.; Zhao, L.-S.; Xu, Y. Impact of NH4+ nitrogen source on the production of Rhizopus oryzae lipase in Pichia pastoris. Process Biochem. 2013, 48, 1462–1468. [Google Scholar] [CrossRef]
- Rumjantsev, A.M.; Bondareva, O.V.; Padkina, M.V.; Sambuk, E.V. Effect of nitrogen source and inorganic phosphate concentration on methanol utilization and PEX genes expression in Pichia pastoris. Sci. World J. 2014, 2014, 743615. [Google Scholar] [CrossRef]
- Ayed, A.; Rabhi, I.; Dellagi, K.; Kallel, H. High level production and purification of human interferon α2b in high cell density culture of Pichia pastoris. Enzym. Microb. Technol. 2008, 42, 173–180. [Google Scholar] [CrossRef]
- Ahmad, M.; Hirz, M.; Pichler, H.; Schwab, H. Protein expression in Pichia pastoris: Recent achievements and perspectives for heterologous protein production. Appl. Microbiol. Biotechnol. 2014, 98, 5301–5317. [Google Scholar] [CrossRef]
- Barone, G.D.; Emmerstorfer-Augustin, A.; Biundo, A.; Pisano, I.; Coccetti, P.; Mapelli, V.; Camattari, A. Industrial production of proteins with Pichia pastoris-Komagataella phaffii. Biomolecules 2023, 13, 441. [Google Scholar] [CrossRef]
- Gleeson, M.A.; White, C.E.; Meininger, D.P.; Komives, E.A. Generation of protease-deficient strains and their use in heterologous protein expression. Methods Mol. Biol. 1998, 103, 81–94. [Google Scholar] [CrossRef]
- Brady, J.R.; Whittaker, C.A.; Tan, M.C.; Kristensen, D.L.; Ma, D.; Dalvie, N.C.; Love, K.R.; Love, J.C. Comparative genome-scale analysis of Pichia pastoris variants informs selection of an optimal base strain. Biotechnol. Bioeng. 2020, 117, 543–555. [Google Scholar] [CrossRef]
- Vijayakumar, V.E.; Venkataraman, K. A Systematic review of the potential of Pichia pastoris (Komagataella phaffii) as an alternative host for biologics production. Mol. Biotechnol. 2023, 1–19. [Google Scholar] [CrossRef]
- Nakamura, Y.; Nishi, T.; Noguchi, R.; Ito, Y.; Watanabe, T.; Nishiyama, T.; Aikawa, S.; Hasunuma, T.; Ishii, J.; Okubo, Y.; et al. A Stable, Autonomously Replicating Plasmid Vector C containing Pichia pastoris centromeric DNA. Appl. Envion. Microbiol. 2018, 84, e02882-17. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Gao, J.; Cao, M.; Dong, C.; Lian, J.; Huang, L.; Cai, J.; Xu, Z. Construction of a series of episomal plasmids and their application in the development of an efficient CRISPR/Cas9 system in Pichia pastoris. World J. Microbiol. Biotechnol. 2019, 35, 79. [Google Scholar] [CrossRef]
- Inan, M. Studies on the Alcohol Oxidase (AOX1) Promoter of Pichia pastoris; The University of Nebraska: Lincoln, NE, USA, 2000. [Google Scholar]
- Wang, X.; Wang, Q.; Wang, J.; Bai, P.; Shi, L.; Shen, W.; Zhou, M.; Zhou, X.; Zhang, Y.; Cai, M. Mit1 transcription factor mediates methanol signaling and regulates the Alcohol Oxidase 1 (AOX1) Promoter in Pichia pastoris. J. Biol. Chem. 2016, 291, 6245–6261. [Google Scholar] [CrossRef]
- Haghighi Poodeh, S.; Ranaei Siadat, S.O.; Arjmand, S.; Khalifeh Soltani, M. Improving AOX1 promoter efficiency by overexpression of Mit1 transcription factor. Mol. Biol. Rep. 2022, 49, 9379–9386. [Google Scholar] [CrossRef] [PubMed]
- Sahu, U.; Krishna Rao, K.; Rangarajan, P.N. Trm1p, a Zn(II)2Cys6-type transcription factor, is essential for the transcriptional activation of genes of methanol utilization pathway, in Pichia pastoris. Biochem. Biophys. Res. Commun. 2014, 451, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Waterham, H.R.; Digan, M.E.; Koutz, P.J.; Lair, S.V.; Cregg, J.M. Isolation of the Pichia pastoris glyceraldehyde-3-phosphate dehydrogenase gene and regulation and use of its promoter. Gene 1997, 186, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Garrigós-Martínez, J.; Vuoristo, K.; Nieto-Taype, M.A.; Tähtiharju, J.; Uusitalo, J.; Tukiainen, P.; Schmid, C.; Tolstorukov, I.; Madden, K.; Penttilä, M.; et al. Bioprocess performance analysis of novel methanol-independent promoters for recombinant protein production with Pichia pastoris. Microb. Cell Factories 2021, 20, 74. [Google Scholar] [CrossRef]
- Baghban, R.; Farajnia, S.; Ghasemi, Y.; Mortazavi, M.; Zarghami, N.; Samadi, N. New developments in Pichia pastoris expression system, review and update. Curr. Pharm. Biotechnol. 2018, 19, 451–467. [Google Scholar] [CrossRef]
- Döring, F.; Klapper, M.; Theis, S.; Daniel, H. Use of the glyceraldehyde-3-phosphate dehydrogenase promoter for production of functional mammalian membrane transport proteins in the yeast Pichia pastoris. Biochem. Biophys. Res. Commun. 1998, 250, 531–535. [Google Scholar] [CrossRef]
- Ata, Ö.; Prielhofer, R.; Gasser, B.; Mattanovich, D.; Çalık, P. Transcriptional engineering of the glyceraldehyde-3-phosphate dehydrogenase promoter for improved heterologous protein production in Pichia pastoris. Biotechnol. Bioeng. 2017, 114, 2319–2327. [Google Scholar] [CrossRef]
- Jiang, B.; Argyros, R.; Bukowski, J.; Nelson, S.; Sharkey, N.; Kim, S.; Copeland, V.; Davidson, R.C.; Chen, R.; Zhuang, J.; et al. Inactivation of a GAL4-like transcription factor improves cell fitness and product yield in glycoengineered Pichia pastoris strains. Appl. Envion. Microbiol. 2015, 81, 260–271. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, X.; Shi, L.; Qi, F.; Zhang, P.; Zhang, Y.; Zhou, X.; Song, Z.; Cai, M. Methanol-independent protein expression by AOX1 Promoter with trans-acting elements engineering and glucose-glycerol-shift induction in Pichia pastoris. Sci. Rep. 2017, 7, 41850. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-H.; Hsiung, H.-A.; Hong, K.-L.; Huang, C.-T. Enhancing the efficiency of the Pichia pastoris AOX1 promoter via the synthetic positive feedback circuit of transcription factor Mxr1. BMC Biotechnol. 2018, 18, 81. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Kong, C.; Xue, Y.; Liu, Y.; Cai, M.; Zhang, Y.; Jiang, T.; Zhou, X.; Zhou, M. Kinase screening in Pichia pastoris identified promising targets involved in cell growth and Alcohol Oxidase 1 promoter (PAOX1) regulation. PLoS ONE 2016, 11, e0167766. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Xue, Y.; Liu, Y.; Kong, C.; Wang, X.; Huang, M.; Cai, M.; Zhou, X.; Zhang, Y.; Zhou, M. A novel methanol-free Pichia pastoris system for recombinant protein expression. Microb. Cell Factories 2016, 15, 178. [Google Scholar] [CrossRef] [PubMed]
- Dou, W.; Zhu, Q.; Zhang, M.; Jia, Z.; Guan, W. Screening and evaluation of the strong endogenous promoters in Pichia pastoris. Microb. Cell Factories 2021, 20, 156. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, S.; Lu, L.; Zhang, C.; Cai, F.; Lin, Y.; Huang, Y. Isolation and evaluation of strong endogenous promoters for the heterologous expression of proteins in Pichia pastoris. World J. Microbiol. Biotechnol. 2022, 38, 226. [Google Scholar] [CrossRef]
- Erden-Karaoğlan, F.; Karaoğlan, M. Applicability of the heterologous yeast promoters for recombinant protein production in Pichia pastoris. Appl. Microbiol. Biotechnol. 2022, 106, 7073–7083. [Google Scholar] [CrossRef]
- Erden-Karaoğlan, F.; Karaoğlan, M.; Yılmaz, G.; Yılmaz, S.; İnan, M. Deletion analysis of Pichia pastoris alcohol dehydrogenase 2 (ADH2) promoter and development of synthetic promoters. Biotechnol. J. 2022, 17, e2100332. [Google Scholar] [CrossRef]
- Nong, L.; Zhang, Y.; Duan, Y.; Hu, S.; Lin, Y.; Liang, S. Engineering the regulatory site of the catalase promoter for improved heterologous protein production in Pichia pastoris. Biotechnol. Lett. 2020, 42, 2703–2709. [Google Scholar] [CrossRef]
- Karaoglan, M.; Karaoglan, F.E.; Inan, M. Comparison of ADH3 promoter with commonly used promoters for recombinant protein production in Pichia pastoris. Protein Expr. Purif. 2016, 121, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Resina, D.; Cos, O.; Ferrer, P.; Valero, F. Developing high cell density fed-batch cultivation strategies for heterologous protein production in Pichia pastoris using the nitrogen source-regulated FLD1 Promoter. Biotechnol. Bioeng. 2005, 91, 760–767. [Google Scholar] [CrossRef] [PubMed]
- Kranthi, B.V.; Kumar, H.R.V.; Rangarajan, P.N. Identification of Mxr1p-binding sites in the promoters of genes encoding dihydroxyacetone synthase and peroxin 8 of the methylotrophic yeast Pichia pastoris. Yeast 2010, 27, 705–711. [Google Scholar] [CrossRef] [PubMed]
- Prielhofer, R.; Reichinger, M.; Wagner, N.; Claes, K.; Kiziak, C.; Gasser, B.; Mattanovich, D. Superior protein titers in half the fermentation time: Promoter and process engineering for the glucose-regulated GTH1 promoter of Pichia pastoris. Biotechnol. Bioeng. 2018, 115, 2479–2488. [Google Scholar] [CrossRef] [PubMed]
- Tschopp, J.F.; Brust, P.F.; Cregg, J.M.; Stillman, C.A.; Gingeras, T.R. Expression of the lacZ gene from two methanol-regulated promoters in Pichia pastoris. Nucleic Acids Res. 1987, 15, 3859–3876. [Google Scholar] [CrossRef]
- Liang, S.; Zou, C.; Lin, Y.; Zhang, X.; Ye, Y. Identification and characterization of PGCW14: A novel, strong constitutive promoter of Pichia pastoris. Biotechnol. Lett. 2013, 35, 1865–1871. [Google Scholar] [CrossRef]
- Vogl, T.; Ruth, C.; Pitzer, J.; Kickenweiz, T.; Glieder, A. Synthetic core promoters for Pichia pastoris. ACS Synth. Biol. 2014, 3, 188–191. [Google Scholar] [CrossRef]
- Portela, R.M.C.; Vogl, T.; Kniely, C.; Fischer, J.E.; Oliveira, R.; Glieder, A. Synthetic core promoters as universal parts for fine-tuning expression in different yeast species. ACS Synth. Biol. 2017, 6, 471–484. [Google Scholar] [CrossRef]
- Damasceno, L.M.; Anderson, K.A.; Ritter, G.; Cregg, J.M.; Old, L.J.; Batt, C.A. Cooverexpression of chaperones for enhanced secretion of a single-chain antibody fragment in Pichia pastoris. Appl. Microbiol. Biotechnol. 2007, 74, 381–389. [Google Scholar] [CrossRef]
- Owji, H.; Nezafat, N.; Negahdaripour, M.; Hajiebrahimi, A.; Ghasemi, Y. A comprehensive review of signal peptides: Structure, roles, and applications. Eur. J. Cell Biol. 2018, 97, 422–441. [Google Scholar] [CrossRef]
- Ahn, J.; Jang, M.J.; Ang, K.S.; Lee, H.; Choi, E.S.; Lee, D.Y. Codon optimization of Saccharomyces cerevisiae mating factor alpha prepro-leader to improve recombinant protein production in Pichia pastoris. Biotechnol. Lett. 2016, 38, 2137–2143. [Google Scholar] [CrossRef] [PubMed]
- Barrero, J.J.; Casler, J.C.; Valero, F.; Ferrer, P.; Glick, B.S. An improved secretion signal enhances the secretion of model proteins from Pichia pastoris. Microb. Cell Factories 2018, 17, 161. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Ishigami, M.; Hashiba, N.; Nakamura, Y.; Terai, G.; Hasunuma, T.; Ishii, J.; Kondo, A. Avoiding entry into intracellular protein degradation pathways by signal mutations increases protein secretion in Pichia pastoris. Microb. Biotechnol. 2022, 15, 2364–2378. [Google Scholar] [CrossRef] [PubMed]
- Duan, G.; Ding, L.; Wei, D.; Zhou, H.; Chu, J.; Zhang, S.; Qian, J. Screening endogenous signal peptides and protein folding factors to promote the secretory expression of heterologous proteins in Pichia pastoris. J. Biotechnol. 2019, 306, 193–202. [Google Scholar] [CrossRef]
- Xiong, R.; Chen, J.; Chen, J. Secreted expression of human lysozyme in the yeast Pichia pastoris under the direction of the signal peptide from human serum albumin. Biotechnol. Appl. Biochem. 2008, 51, 129–134. [Google Scholar] [CrossRef]
- Vadhana, A.K.P.; Samuel, P.; Berin, R.M.; Krishna, J.; Kamatchi, K.; Meenakshisundaram, S. Improved secretion of Candida antarctica lipase B with its native signal peptide in Pichia pastoris. Enzym. Microb. Technol. 2013, 52, 177–183. [Google Scholar] [CrossRef]
- Peña, D.A.; Gasser, B.; Zanghellini, J.; Steiger, M.G.; Mattanovich, D. Metabolic engineering of Pichia pastoris. Metab. Eng. 2018, 50, 2–15. [Google Scholar] [CrossRef]
- Wu, X.; Cai, P.; Yao, L.; Zhou, Y.J. Genetic tools for metabolic engineering of Pichia pastoris. Eng. Microbiol. 2023, 3, 100094. [Google Scholar] [CrossRef]
- Weninger, A.; Hatzl, A.-M.; Schmid, C.; Vogl, T.; Glieder, A. Combinatorial optimization of CRISPR/Cas9 expression enables precision genome engineering in the methylotrophic yeast Pichia pastoris. J. Biotechnol. 2016, 235, 139–149. [Google Scholar] [CrossRef]
- Weninger, A.; Fischer, J.E.; Raschmanová, H.; Kniely, C.; Vogl, T.; Glieder, A. Expanding the CRISPR/Cas9 toolkit for Pichia pastoris with efficient donor integration and alternative resistance markers. J. Cell Biochem. 2018, 119, 3183–3198. [Google Scholar] [CrossRef]
- Liu, Q.; Shi, X.; Song, L.; Liu, H.; Zhou, X.; Wang, Q.; Zhang, Y.; Cai, M. CRISPR-Cas9-mediated genomic multiloci integration in Pichia pastoris. Microb. Cell Factories 2019, 18, 144. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Xu, J.; Zuo, Y.; Ye, C.; Jiang, L.; Feng, L.; Huang, L.; Xu, Z.; Lian, J. Synthetic biology toolkit for marker-less integration of multigene pathways into Pichia pastoris via CRISPR/Cas9. ACS Synth. Biol. 2022, 11, 623–633. [Google Scholar] [CrossRef]
- Gao, J.; Ye, C.; Cheng, J.; Jiang, L.; Yuan, X.; Lian, J. Enhancing homologous recombination efficiency in Pichia pastoris for multiplex genome integration using short homology arms. ACS Synth. Biol. 2022, 11, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Sandberg, T.E.; Salazar, M.J.; Weng, L.L.; Palsson, B.O.; Feist, A.M. The emergence of adaptive laboratory evolution as an efficient tool for biological discovery and industrial biotechnology. Metab. Eng. 2019, 56, 1–16. [Google Scholar] [CrossRef]
- Hirasawa, T.; Maeda, T. Adaptive laboratory evolution of microorganisms: Methodology and application for bioproduction. Microorganisms 2022, 11, 92. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kim, P. Current status and applications of adaptive laboratory evolution in industrial microorganisms. J. Microbiol. Biotechnol. 2020, 30, 793–803. [Google Scholar] [CrossRef]
- Moser, J.W.; Prielhofer, R.; Gerner, S.M.; Graf, A.B.; Wilson, I.B.H.; Mattanovich, D.; Dragosits, M. Implications of evolutionary engineering for growth and recombinant protein production in methanol-based growth media in the yeast Pichia pastoris. Microb. Cell Factories 2017, 16, 49. [Google Scholar] [CrossRef] [PubMed]
- Bankefa, O.E.; Samuel-Osamoka, F.C.; Oladeji, S.J. Improved enzyme production on corncob hydrolysate by a xylose-evolved Pichia pastoris cell factory. J. Food Sci. Technol. 2022, 59, 1280–1287. [Google Scholar] [CrossRef]
- Gassler, T.; Sauer, M.; Gasser, B.; Egermeier, M.; Troyer, C.; Causon, T.; Hann, S.; Mattanovich, D.; Steiger, M.G. The industrial yeast Pichia pastoris is converted from a heterotroph into an autotroph capable of growth on CO2. Nat. Biotechnol. 2020, 38, 210–216. [Google Scholar] [CrossRef]
- Gassler, T.; Baumschabl, M.; Sallaberger, J.; Egermeier, M.; Mattanovich, D. Adaptive laboratory evolution and reverse engineering enhances autotrophic growth in Pichia pastoris. Metab. Eng. 2022, 69, 112–121. [Google Scholar] [CrossRef]
- Higgins, D.R.; Cregg, J.M. Introduction to Pichia pastoris. Methods Mol. Biol. 1998, 103, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Damasceno, L.M.; Huang, C., Jr.; Batt, C.A. Protein secretion in Pichia pastoris and advances in protein production. Appl. Microbiol. Biotechnol. 2012, 93, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Y.; Xiao, F.; Gao, J.; Ye, C.; Jiang, L.; Dong, C.; Lian, J. Establishing Komagataella phaffii as a cell factory for efficient production of sesquiterpenoid α-Santalene. J. Agric. Food Chem. 2022, 70, 8024–8031. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Zuo, Y.; Xiao, F.; Wang, Y.; Li, D.; Xu, J.; Ye, C.; Feng, L.; Jiang, L.; Liu, T.; et al. Biosynthesis of catharanthine in engineered Pichia pastoris. Nat. Synth. 2023, 2, 231–242. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, D.; Duan, Y.; Zheng, X.; Lin, Y.; Liang, S. Production of lycopene by metabolically engineered Pichia pastoris. Biosci. Biotechnol. Biochem. 2020, 84, 463–470. [Google Scholar] [CrossRef]
- Chen, S.-L.; Liu, T.-S.; Zhang, W.-G.; Xu, J.-Z. Cofactor engineering for efficient production of α-Farnesene by rational modification of NADPH and ATP regeneration pathway in Pichia pastoris. Int. J. Mol. Sci. 2023, 24, 1767. [Google Scholar] [CrossRef]
- Jeong, E.; Shim, W.Y.; Kim, J.H. Metabolic engineering of Pichia pastoris for production of hyaluronic acid with high molecular weight. J. Biotechnol. 2014, 185, 28–36. [Google Scholar] [CrossRef]
- Jin, X.; Zhang, W.; Wang, Y.; Sheng, J.; Xu, R.; Li, J.; Du, G.; Kang, Z. Biosynthesis of non-animal chondroitin sulfate from methanol using genetically engineered Pichia pastoris. Green Chem. 2021, 23, 4365–4374. [Google Scholar] [CrossRef]
- Qian, D.; Zhang, C.; Deng, C.; Zhou, M.; Fan, L.; Zhao, L. De novo biosynthesis of 2′-fucosyllactose in engineered Pichia pastoris. Biotechnol. Lett. 2023, 45, 521–536. [Google Scholar] [CrossRef]
- Gao, L.; Cai, M.; Shen, W.; Xiao, S.; Zhou, X.; Zhang, Y. Engineered fungal polyketide biosynthesis in Pichia pastoris: A potential excellent host for polyketide production. Microb. Cell Factories 2013, 12, 77. [Google Scholar] [CrossRef]
- Xue, Y.; Kong, C.; Shen, W.; Bai, C.; Ren, Y.; Zhou, X.; Zhang, Y.; Cai, M. Methylotrophic yeast Pichia pastoris as a chassis organism for polyketide synthesis via the full citrinin biosynthetic pathway. J. Biotechnol. 2017, 242, 64–72. [Google Scholar] [CrossRef]
- Liu, Y.; Tu, X.; Xu, Q.; Bai, C.; Kong, C.; Liu, Q.; Yu, J.; Peng, Q.; Zhou, X.; Zhang, Y.; et al. Engineered monoculture and co-culture of methylotrophic yeast for de novo production of monacolin J and lovastatin from methanol. Metab. Eng. 2018, 45, 189–199. [Google Scholar] [CrossRef]
- Qian, Z.; Yu, J.; Chen, X.; Kang, Y.; Ren, Y.; Liu, Q.; Lu, J.; Zhao, Q.; Cai, M. De novo production of plant 4′-Deoxyflavones baicalein and oroxylin A from ethanol in Crabtree-negative yeast. ACS Synth. Biol. 2022, 11, 1600–1612. [Google Scholar] [CrossRef] [PubMed]
- Kumokita, R.; Bamba, T.; Inokuma, K.; Yoshida, T.; Ito, Y.; Kondo, A.; Hasunuma, T. Construction of an L-Tyrosine chassis in Pichia pastoris enhances aromatic secondary metabolite production from glycerol. ACS Synth. Biol. 2022, 11, 2098–2107. [Google Scholar] [CrossRef] [PubMed]
- Bumrungtham, P.; Promdonkoy, P.; Prabmark, K.; Bunterngsook, B.; Boonyapakron, K.; Tanapongpipat, S.; Champreda, V.; Runguphan, W. Engineered production of isobutanol from sugarcane trash hydrolysates in Pichia pastoris. J. Fungi 2022, 8, 767. [Google Scholar] [CrossRef] [PubMed]
- Siripong, W.; Angela, C.; Tanapongpipat, S.; Runguphan, W. Metabolic engineering of Pichia pastoris for production of isopentanol (3-Methyl-1-butanol). Enzym. Microb. Technol. 2020, 138, 109557. [Google Scholar] [CrossRef]
- Guo, F.; Dai, Z.; Peng, W.; Zhang, S.; Zhou, J.; Ma, J.; Dong, W.; Xin, F.; Zhang, W.; Jiang, M. Metabolic engineering of Pichia pastoris for malic acid production from methanol. Biotechnol. Bioeng. 2021, 118, 357–371. [Google Scholar] [CrossRef]
- Hamers-Casterman, C.; Atarhouch, T.; Muyldermans, S.; Robinson, G.; Hamers, C.; Songa, E.B.; Bendahman, N.; Hamers, R. Naturally occurring antibodies devoid of light chains. Nature 1993, 363, 446–448. [Google Scholar] [CrossRef]
- Greenberg, A.S.; Avila, D.; Hughes, M.; Hughes, A.; McKinney, E.C.; Flajnik, M.F. A new antigen receptor gene family that undergoes rearrangement and extensive somatic diversification in sharks. Nature 1995, 374, 168–173. [Google Scholar] [CrossRef]
- Hassanzadeh-Ghassabeh, G.; Devoogdt, N.; De Pauw, P.; Vincke, C.; Muyldermans, S. Nanobodies and their potential applications. Nanomedicine 2013, 8, 1013–1026. [Google Scholar] [CrossRef]
- Huo, J.; Mikolajek, H.; Le Bas, A.; Clark, J.J.; Sharma, P.; Kipar, A.; Dormon, J.; Norman, C.; Weckener, M.; Clare, D.K.; et al. A potent SARS-CoV-2 neutralising nanobody shows therapeutic efficacy in the Syrian golden hamster model of COVID-19. Nat. Commun. 2021, 12, 5469. [Google Scholar] [CrossRef] [PubMed]
- Muyldermans, S. A guide to: Generation and design of nanobodies. FEBS J. 2021, 288, 2084–2102. [Google Scholar] [CrossRef]
- Baghban, R.; Gargari, S.L.M.; Rajabibazl, M.; Nazarian, S.; Bakherad, H. Camelid-derived heavy-chain nanobody against Clostridium botulinum neurotoxin E in Pichia pastoris. Biotechnol. Appl. Biochem. 2016, 63, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Xian, Z.; Ma, L.; Zhu, M.; Li, G.; Gai, J.; Chang, Q.; Huang, Y.; Ju, D.; Wan, Y. Blocking the PD-1-PD-L1 axis by a novel PD-1 specific nanobody expressed in yeast as a potential therapeutic for immunotherapy. Biochem. Biophys. Res. Commun. 2019, 519, 267–273. [Google Scholar] [CrossRef]
- Chen, Q.; Zhou, Y.; Yu, J.; Liu, W.; Li, F.; Xian, M.; Nian, R.; Song, H.; Feng, D. An efficient constitutive expression system for Anti-CEACAM5 nanobody production in the yeast Pichia pastoris. Protein Expr. Purif. 2019, 155, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Qiu, C.; Wen, B.; Wang, S.; Zhu, L.; Zhao, L.; Li, H. A bispecific nanobody targeting the dimerization interface of epidermal growth factor receptor: Evidence for tumor suppressive actions in vitro and in vivo. Biochem. Biophys. Res. Commun. 2021, 548, 78–83. [Google Scholar] [CrossRef]
- Zhu, W.; Gong, G.; Pan, J.; Han, S.; Zhang, W.; Hu, Y.; Xie, L. High level expression and purification of recombinant human serum albumin in Pichia pastoris. Protein Expr. Purif. 2018, 147, 61–68. [Google Scholar] [CrossRef]
- Peng, B.; Xue, G.; Xu, D.; Feng, Z.; Chen, J.; Huang, M.; Lu, H.; Gong, L. Expression and purification of recombinant serine protease domain of human coagulation factor XII in Pichia pastoris. Biosci. Biotechnol. Biochem. 2019, 83, 1815–1821. [Google Scholar] [CrossRef]
- Cai, H.; Yao, H.; Li, T.; Tang, Y.; Li, D. High-level heterologous expression of the human transmembrane sterol Δ8,Δ7-isomerase in Pichia pastoris. Protein Expr. Purif. 2019, 164, 105463. [Google Scholar] [CrossRef]
- Gurramkonda, C.; Polez, S.; Skoko, N.; Adnan, A.; Gäbel, T.; Chugh, D.; Swaminathan, S.; Khanna, N.; Tisminetzky, S.; Rinas, U. Application of simple fed-batch technique to high-level secretory production of insulin precursor using Pichia pastoris with subsequent purification and conversion to human insulin. Microb. Cell Factories 2010, 9, 31. [Google Scholar] [CrossRef]
- Polez, S.; Origi, D.; Zahariev, S.; Guarnaccia, C.; Tisminetzky, S.G.; Skoko, N.; Baralle, M. A simplified and efficient process for insulin production in Pichia pastoris. PLoS ONE 2016, 11, e0167207. [Google Scholar] [CrossRef] [PubMed]
- Katla, S.; Karmakar, B.; Tadi, S.R.R.; Mohan, N.; Anand, B.; Pal, U.; Sivaprakasam, S. High level extracellular production of recombinant human interferon alpha 2b in glycoengineered Pichia pastoris: Culture medium optimization, high cell density cultivation and biological characterization. J. Appl. Microbiol. 2019, 126, 1438–1453. [Google Scholar] [CrossRef] [PubMed]
- Tholl, D. Biosynthesis and biological functions of terpenoids in plants. Adv. Biochem. Eng. Biotechnol. 2015, 148, 63–106. [Google Scholar] [CrossRef] [PubMed]
- Akazawa, H.; Kohno, H.; Tokuda, H.; Suzuki, N.; Yasukawa, K.; Kimura, Y.; Manosroi, A.; Manosroi, J.; Akihisa, T. Anti-inflammatory and anti-tumor-promoting effects of 5-deprenyllupulonol C and other compounds from Hop (Humulus lupulus L.). Chem. Biodivers. 2012, 9, 1045–1054. [Google Scholar] [CrossRef]
- Zhang, Y.; Nielsen, J.; Liu, Z. Engineering yeast metabolism for production of terpenoids for use as perfume ingredients, pharmaceuticals and biofuels. FEMS Yeast Res. 2017, 17, fox080. [Google Scholar] [CrossRef]
- Bhataya, A.; Schmidt-Dannert, C.; Lee, P.C. Metabolic engineering of Pichia pastoris X-33 for lycopene production. Process. Biochem. 2009, 44, 1095–1102. [Google Scholar] [CrossRef]
- Liu, H.; Chen, S.-L.; Xu, J.-Z.; Zhang, W.-G. Dual regulation of cytoplasm and peroxisomes for improved α-Farnesene production in recombinant Pichia pastoris. ACS Synth. Biol. 2021, 10, 1563–1573. [Google Scholar] [CrossRef]
- Prateeksha; Sharma, V.K.; Liu, X.; Oyarzún, D.A.; Abdel-Azeem, A.M.; Atanasov, A.G.; Hesham, A.E.-L.; Barik, S.K.; Gupta, V.K.; Singh, B.N. Microbial polysaccharides: An emerging family of natural biomaterials for cancer therapy and diagnostics. Semin. Cancer Biol. 2022, 86, 706–731. [Google Scholar] [CrossRef]
- Liu, L.; Liu, Y.; Li, J.; Du, G.; Chen, J. Microbial production of hyaluronic acid: Current state, challenges, and perspectives. Microb. Cell Factories 2011, 10, 99. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Promoters | Regulation | Encoding Gene | Expression Level | Refs. |
|---|---|---|---|---|
| PAOX1 | Induced by methanol | Alcohol oxidase | Strong | [36] |
| PADH2 | Induced by ethanol | Alcohol dehydrogenase | Stronger than PGAP | [51] |
| PSNT5 | Induced by ethanol | synthetic promoter | Strong | [51] |
| PCAT1 | Induced by methanol and oleic acid | Catalase | Similar to PAOX1 | [52] |
| PLAR3 | Induced by L-Rhamnose | L-rhamnose dehydratase | Similar to PGAP | [20] |
| PLAR4 | Induced by L-Rhamnose | L-KDR aldolase | Weak | [20] |
| PADH3 | Induced by ethanol and methanol | Alcohol dehydrogenase | Strong | [53] |
| PFLD1 | Induced by methanol and methylamine | Formaldehyde dehydrogenase | Similar to PGAP | [54] |
| PPEX8 | Induced by methanol | Peroxisomal matrix protein | Weak | [55] |
| PGTH1 | Induced by glucose | High-affinity glucose transporter | Similar to PAOX1 | [56] |
| PDAS1 | Induced by methanol | Dihydroxyacetone phosphate | Strong | [57] |
| PFDH1 | Induced by methanol | Formate dehydrogenase 1 | Strong | [49] |
| PICL1 | Induced by ethanol | Isocitrate lyase | ~10% of PAOX1 | [50] |
| PCUPI | Induced by glucose | Copper-binding metallothionein protein | ~10% of PAOX1 | [50] |
| PGAP | Constitutive | Glyceraldehyde-3-phosphate dehydrogenase | Strong | [38] |
| PGCW14 | Constitutive | Glycosylphosphatidylinositol | Strong | [58] |
| PTPI | Constitutive | Triose phosphate isomerase | ~4% of PGAP | [50] |
| PPMA | Constitutive | Plasma membrane (H+) ATPases | ~81% of PGAP | [50] |
| Classification | Products | Titer | Synthetic Strategies | Culture Conditions | Ref. |
|---|---|---|---|---|---|
| Terpenoids | α-Santalene | 21.5 g/L | Overexpression of tHMG1 and IDI1, increase in SAS copy number | Glucose/Fermenter | [85] |
| Catharanthine | 2.57 mg/L | Optimization of pathway genes and integration sites | Glucose/Fermenter | [86] | |
| Lycopene | 6.146 mg/g | Promoter selection, increase in precursor supply and GGPP accumulation | Methanol/Shake flask | [87] | |
| α-Farnesene | 3.09 g/L | Pathway reconstruction for the synthesis of NADPH and ATP | Glucose/Shake flask | [88] | |
| Polysaccharides | Hyaluronic acid | 0.8–1.7 g/L | Induction by PAOX2, low-temperature cultivation | Glucose/Fermenter | [89] |
| Chondroitin sulfate | 2.1 g/L | Codon optimization, endogenous promoter analysis, enhancement of the PAPS synthesis pathway | Methanol/Fermenter | [90] | |
| 2′-Fucosyllactose | 0.276 g/L | GAP promoter control | Glucose/Fermenter | [91] | |
| Polyketides | 6-Methylsalicylic acid | 2.2 g/L | Heterologous expression of atX from Aspergillus terreus and npgA from Aspergillus nidulans | Methanol/Fermenter | [92] |
| Citrinin | 0.6 mg/L | Heterologous expression of pksCT from Monascus purpureus and npgA from Aspergillus nidulans, introduction of mpl6 and mpl7 from Monascus purpureus | Glucose/Shake flask | [93] | |
| Monacolin J | 593.9 mg/L | Optimization of pathway gene expression, co-culture | Methanol/Fermenter | [94] | |
| Flavonoids | Baicalein | 401.9 mg/L | Ethanol induction, modularization of metabolic pathway | Ethanol/Shake flask | [95] |
| Oroxylin A | 339.5 mg/L | Ethanol induction, modularization of metabolic pathway | Ethanol/Shake flask | [95] | |
| Naringenin | 1067 mg/L | Overexpression of ARO4K229L and ARO7G141S for efficient production of L-tyrosine | Glycerol/Fermenter | [96] | |
| Alcohols | Isobutanol | 48.2 mg/L | Overexpression of heterologous xylose isomerase and endogenous xylulokinase, introduction of the isobutanol pathway | Glucose/Microfermenter | [97] |
| Isopentanol | 191.0 mg/L | Overexpression of the endogenous valine/leucine biosynthetic pathways, and artificial keto-acid degradation pathway | Glucose/Microfermenter | [98] | |
| Organic acids | Malic acid | 2.79 g/L | Knockout of gpi optimized methanol assimilation, optimization of nitrogen sources | Methanol/Shake flask | [99] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zha, J.; Liu, D.; Ren, J.; Liu, Z.; Wu, X. Advances in Metabolic Engineering of Pichia pastoris Strains as Powerful Cell Factories. J. Fungi 2023, 9, 1027. https://doi.org/10.3390/jof9101027
Zha J, Liu D, Ren J, Liu Z, Wu X. Advances in Metabolic Engineering of Pichia pastoris Strains as Powerful Cell Factories. Journal of Fungi. 2023; 9(10):1027. https://doi.org/10.3390/jof9101027
Chicago/Turabian StyleZha, Jian, Dan Liu, Juan Ren, Zhijun Liu, and Xia Wu. 2023. "Advances in Metabolic Engineering of Pichia pastoris Strains as Powerful Cell Factories" Journal of Fungi 9, no. 10: 1027. https://doi.org/10.3390/jof9101027
APA StyleZha, J., Liu, D., Ren, J., Liu, Z., & Wu, X. (2023). Advances in Metabolic Engineering of Pichia pastoris Strains as Powerful Cell Factories. Journal of Fungi, 9(10), 1027. https://doi.org/10.3390/jof9101027