
Nuclear-Localized Fluorescent Proteins Enable Visualization of Nuclear Behavior in the Basidiomycete Schizophyllum commune Early Mating Interactions


Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture Conditions, Strains, Cloning and Mating Interactions
2.2. Spinning Disc Confocal Microscop
2.3. Recording of Nuclear Numbers with Different Colors
3. Results
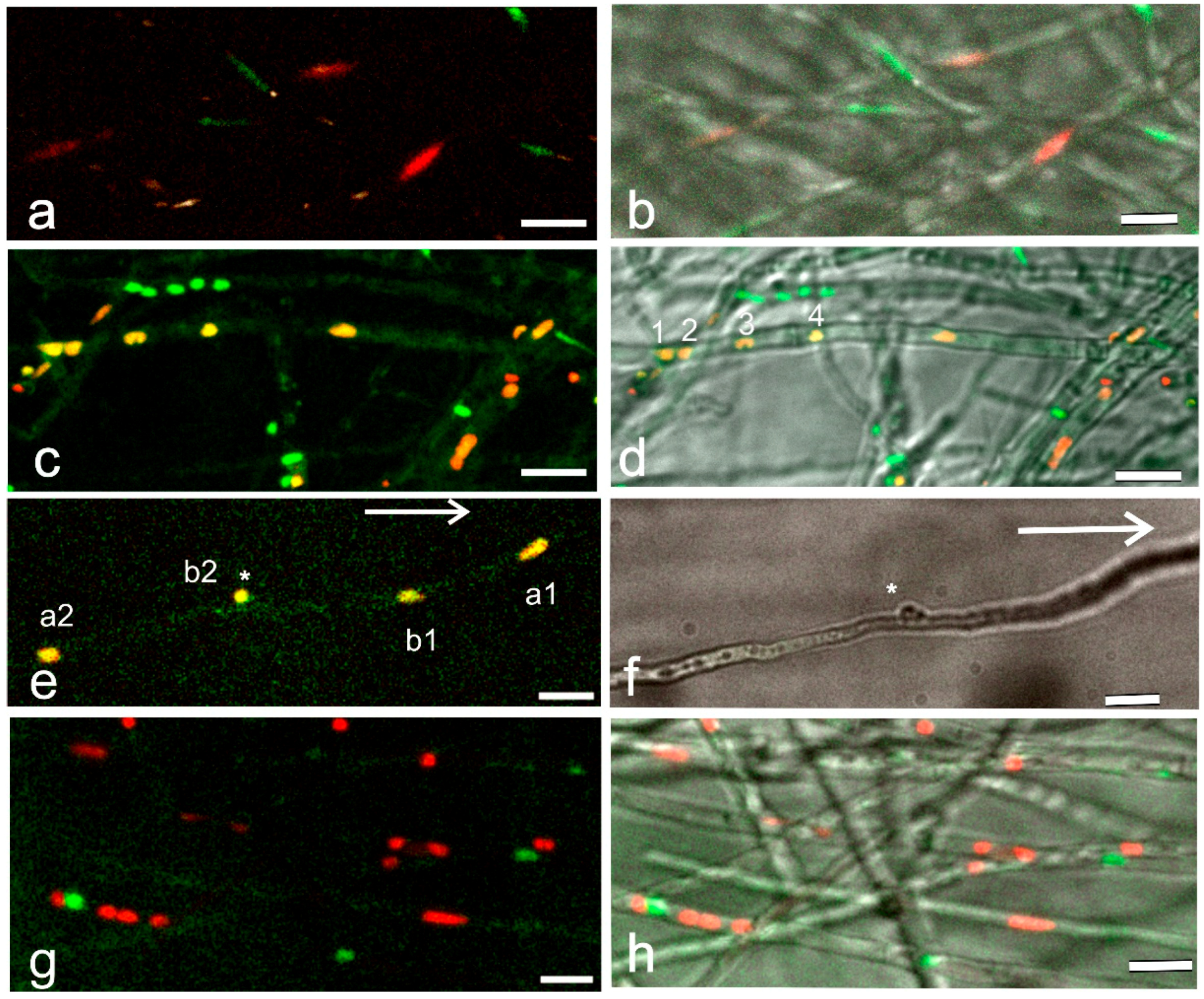
3.1. Spinning Disc Confocal Projections of Mating Interactions with Different-Colored Nuclei—An Overview
3.2. Details of Mating Interactions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mating Interactions | Number of Nuclei with Different Colors | Total Number of Nuclei | ||
|---|---|---|---|---|
| Green | Red | Yellow | ||
| Incompatible A = B = | 45 | 29 | 1 | 75 * |
| Compatible A ≠ B ≠ | 135 | 25 | 72 | 232 * |
| Hemicompatible A = B ≠ | 34 | 59 | 1 | 94 ** |
3.3. Development of Dikaryotic Hyphae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kües, U.; Casselton, L.A. Homeodomains and regulation of sexual development in basidiomycetes. Trends Genet. 1992, 8, 154–155. [Google Scholar] [CrossRef] [PubMed]
- Raper, J. Genetics of Sexuality in Higher Fungi; Ronald Press: New York, NY, USA, 1966. [Google Scholar]
- Casselton, L.A.; Olesnicky, N.S. Molecular genetics of mating recognition in basidiomycete fungi. Microbiol. Mol. Biol. Rev. 1998, 62, 55–70. [Google Scholar] [CrossRef] [PubMed]
- Kamada, T. Molecular genetics of sexual development in the mushroom Coprinus cinereus. BioEssays 2002, 24, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Raudaskoski, M.; Kothe, E. Basidiomycete mating type genes and pheromone signaling. Eukaryot. Cell 2010, 9, 847–859. [Google Scholar] [CrossRef]
- Raudaskoski, M. Mating-type genes and hyphal fusion in filamentous basidiomycetes. Fungal Biol. Rev. 2015, 29, 179–193. [Google Scholar] [CrossRef]
- Girbardt, M. Ultrastructure and dynamics of the moving nucleus. In Aspects of Cell Motility (23rd Symposium of the Society for Experimental Biology); Miller, P.L., Ed.; University Press: Cambridge, UK, 1968; Volume 22, pp. 249–259. [Google Scholar]
- Raudaskoski, M.; Rupes, J.I.; Timonen, S. lmmunofluorescence microscopy of the cytoskeleton in filamentous fungi after quick-freezing and low-temperature fixation. Exp. Mycol. 1991, 15, 167–173. [Google Scholar] [CrossRef]
- Jung, E.-M.; Kothe, E.; Raudaskoski, M. The making of a mushroom: Mitosis, nuclear migration and the actin network. Fungal Genet. Biol. 2018, 111, 85–91. [Google Scholar] [CrossRef]
- Raudaskoski, M. The central role of septa in the basidiomycete Schizophyllum commune hyphal morphogenesis. Fungal Biol. 2019, 123, 638–649. [Google Scholar] [CrossRef]
- Vaillancourt, L.J.; Raper, C.A. Pheromones and pheromone receptors as mating-type determinants in basidiomycetes. In Genetic Engineering; Setlow, J.K., Ed.; Plenum Press: New York, NY, USA, 1996; Volume 18, pp. 219–247. [Google Scholar]
- Kämper, J.; Friedrich, M.W.; Kahmann, R. Creating novel specificities in a fungal nonself recognition system by single step homologous recombination events. New Phytol. 2020, 228, 1001–1010. [Google Scholar] [CrossRef]
- Coelho, M.A.; Bakkeren, G.; Sun, S.; Hood, M.E.; Giraud, T. Fungal sex: The basidiomycota. Microbiol. Spectr. 2017, 5, 147–175. [Google Scholar] [CrossRef]
- Wallen, R.M.; Perlin, M.H. An overview of the function and maintenance of sexual reproduction in dikaryotic fungi. Front. Microbiol. 2018, 9, 503. [Google Scholar] [CrossRef] [PubMed]
- Koltin, Y.; Flexer, A.S. Alterartion of nuclear distribution in B-mutant strain of Schizophyllum commune. J. Cell Sci. 1969, 4, 739–749. [Google Scholar] [CrossRef]
- Raudaskoski, M. The relationship between B-mating-type genes and nuclear migration in Schizophyllum commune. Fungal Genet. Biol. 1998, 24, 207–227. [Google Scholar] [CrossRef]
- Raper, J.; Raudaskoski, M. Secondary mutations at the Bβ incompatibility locus of Schizophyllum. Heredity 1968, 23, 109–117. [Google Scholar] [CrossRef]
- Weber, M.; Salo, V.; Uuskallio, M.; Raudaskoski, M. Ectopic expression of a constitutively active Cdc42 small GTPase alters the morphology of haploid and dikaryotic hyphae in the filamentous homobasidiomycete Schizophyllum commune. Fungal Genet. Biol. 2005, 42, 624–637. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji—An opensource platform for biological image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Kurat, C.F.; Recht, J.; Radovani, E.; Durbic, T.; Andrews, B.; Fillingham, J. Regulation of histone gene transcription in yeast. Cell. Mol. Life Sci. 2014, 71, 599–613. [Google Scholar] [CrossRef]
- Zhang, W.; Feng, J.; Li, Q. The replisome guides nucleosome assembly during DNA replication. Cell Biosci. 2020, 10, 37. [Google Scholar] [CrossRef]
- Hsu, C.-L.; Chong, S.Y.; Lin, C.-Y.; Kao, C.-F. Histone dynamics during DNA replication stress. J. Biomed. Sci. 2021, 28, 48. [Google Scholar] [CrossRef]
- Guglielmi, B.; La Rochelle, N.; Tjian, R. Gene-specific transcriptional mechanisms at the histone gene cluster revealed by single-cell imaging. Mol. Cell 2013, 51, 480–492. [Google Scholar] [CrossRef]
- Bernardes, N.E.; Chook, Y.M. Nuclear import of histones. Biochem. Soc. Trans. 2020, 48, 2753–2767. [Google Scholar] [CrossRef] [PubMed]
- Giesy, R.M.; Day, P.R. The septal pores of Coprinus lagopus in relation to nuclear migration. Amer. J. Bot. 1965, 52, 287–294. [Google Scholar] [CrossRef]
- Zarnack, K.; Eichhorn, H.; Kahmann, R.; Feldbrugge, M. Pheromone-regulated target genes respond differentially to MAPK phosphorylation of transcription factor Prf1. Mol. Microbiol. 2008, 69, 1041–1053. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, L.; Schwier, M.A.; Heimela, K. The Unfolded Protein Response Regulates Pathogenic Development of Ustilago maydis by Rok1-Dependent Inhibition of Mating-Type Signaling. MBio 2019, 10, e02756-19. [Google Scholar] [CrossRef] [PubMed]
- Fowler, T.J.S.; Mitton, M.F.; Vaillancourt, L.J.; Raper, C.A. Changes in mate recognition through alterations of pheromones and receptors in the multisexual mushroom fungus Schizophyllum commune. Genetics 2001, 158, 1491–1503. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Shen, G.-P.; Park, D.C.; Novotny, C.P.; Ullrich, R.C. The Aα mating-type transcripts of Schizophyllum commune. Exp. Mycol. 1995, 19, 16–25. [Google Scholar] [CrossRef]
- Shen, G.-P.; Park, D.C.; Ullrich, R.C.; Novotny, C.P. Cloning and characterization of a Schizophyllum gene with Aβ6 mating-type activity. Curr. Genet. 1996, 29, 136–142. [Google Scholar] [CrossRef]
- Robertson, I.C.; McMahon Kende, A.; Toenjes, K.; Novotny, C.P.; Ullrich, R.C. Evidence for interaction of Schizophyllum commune Y mating-type proteins in vivo. Genetics 2002, 160, 1461–1467. [Google Scholar] [CrossRef]
- Yi, R.; Tachikawa, T.; Ishikawa, M.; Mukaiyama, H.; Bao, D.; Aimi, T. Genomic structure of the A mating-type locus in a bipolar basidiomycete, Pholiota nameko. Mycol. Res. 2009, 113, 240–248. [Google Scholar] [CrossRef]
- Magae, Y.; Novotny, C.; Ullrich, R. Interaction of the A alpha Y mating-type and Z mating-type homeodomain proteins of Schizophyllum commune detected by the two-hybrid system. Biochiem. Biophys. Res. Commun. 1995, 211, 1071–1076. [Google Scholar] [CrossRef]
- Banham, A.H.; Asante-Owusu, R.N.; Göttgens, B.; Thompson, S.A.J.; Kingsnorth, C.S.; Mellor, E.J.C.; Casselton, L.A. An N-Terminal dimerization domain permits homeodomain proteins to choose compatible partners and initiate sexual development in the mushroom Coprinus cinereus. Plant Cell 1995, 7, 773–783. [Google Scholar] [PubMed]
- Yue, C.; Osier, M.; Novotny, C.P.; Ullrich, C.R. The specificity determinant of the Y mating-Type proteins of Schizophyllum commune is also essential for Y-Z protein binding. Genetics 1997, 145, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Spit, A.; Hyland, R.; Mellor, E.J.C.; Casselton, L.A. Heterodimerization targets a homeodomain protein complex to the nucleus. Proc. Natl. Acad. Sci. USA 1998, 95, 6228–6233. [Google Scholar] [CrossRef] [PubMed]
- Raudaskoski, M. Light and electron microscope study of unilateral mating between secondary mutant and a wild-type strain of Schizophyllum commune. Protoplasma 1973, 76, 35–48. [Google Scholar] [CrossRef]
- Yadav, V.; Sun, S.; Heitman, J. Uniparental nuclear inheritance following bisexual mating in fungi. eLife 2021, 10, e66234. [Google Scholar] [CrossRef]






| Strains | Genotype | Interaction | Literature |
|---|---|---|---|
| I-II H2B::EGFP | A26Bα4-β1 ura-h2B::egfp phleom+ | [9,10] | |
| F16 H2B::EGFP | A43Bα4-β1 h2B::egfp phleom+ | [9,10] | |
| 4 H2B::EGFP | A43Bα3-β6 h2B::egfp phleom+ | [9,10] | |
| T14-3-7 H2B::mCherry | A43BBα3-β6 h2B::mCherry phloem+ | This study | |
| Mating interactions | |||
| 4 H2B::EGFP × T14-3-7 H2B::mCherry | A43Bα3-β6 h2B::egfp × A43BBα3-β6 h2B::mCherry | Incompatible A = B = | This study |
| I-II H2B::EGFP × T14-3-7 H2B::mCherry | A26Bα4-β1 ura-h2B::egfp × A43Bα3-β6 h2B::mCherry | Compatible A ≠ B ≠ | This study |
| F16 H2B::EGFP × T14-3-7 H2B::mCherry | A43Bα4-β1 h2B::egfp × A43BBα3-β6 h2B::mCherry | Hemicompatible A = B ≠ | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raudaskoski, M.; Butler-Hallissey, C. Nuclear-Localized Fluorescent Proteins Enable Visualization of Nuclear Behavior in the Basidiomycete Schizophyllum commune Early Mating Interactions. J. Fungi 2023, 9, 1043. https://doi.org/10.3390/jof9111043
Raudaskoski M, Butler-Hallissey C. Nuclear-Localized Fluorescent Proteins Enable Visualization of Nuclear Behavior in the Basidiomycete Schizophyllum commune Early Mating Interactions. Journal of Fungi. 2023; 9(11):1043. https://doi.org/10.3390/jof9111043
Chicago/Turabian StyleRaudaskoski, Marjatta, and Ciarán Butler-Hallissey. 2023. "Nuclear-Localized Fluorescent Proteins Enable Visualization of Nuclear Behavior in the Basidiomycete Schizophyllum commune Early Mating Interactions" Journal of Fungi 9, no. 11: 1043. https://doi.org/10.3390/jof9111043
APA StyleRaudaskoski, M., & Butler-Hallissey, C. (2023). Nuclear-Localized Fluorescent Proteins Enable Visualization of Nuclear Behavior in the Basidiomycete Schizophyllum commune Early Mating Interactions. Journal of Fungi, 9(11), 1043. https://doi.org/10.3390/jof9111043