
O-Linked Glycans of Candida albicans Interact with Specific GPCRs in the Coronary Endothelium and Inhibit the Cardiac Response to Agonists

Abstract
:1. Introduction
2. Materials and Methods
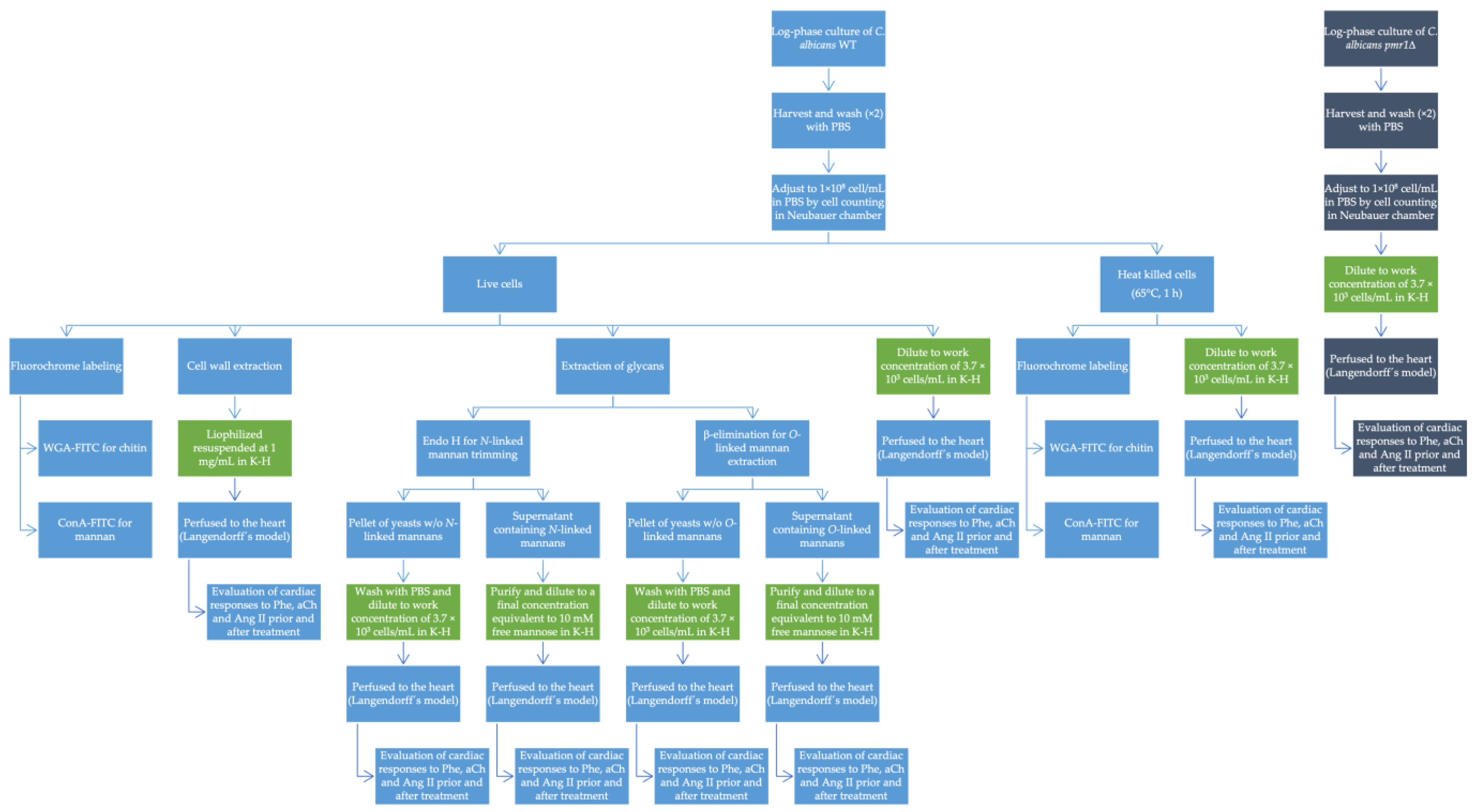
2.1. Strains and Culture Methods
2.2. Fluorochrome Labeling
2.3. Cell Wall Extraction
2.4. Extraction and Purification of Glycans
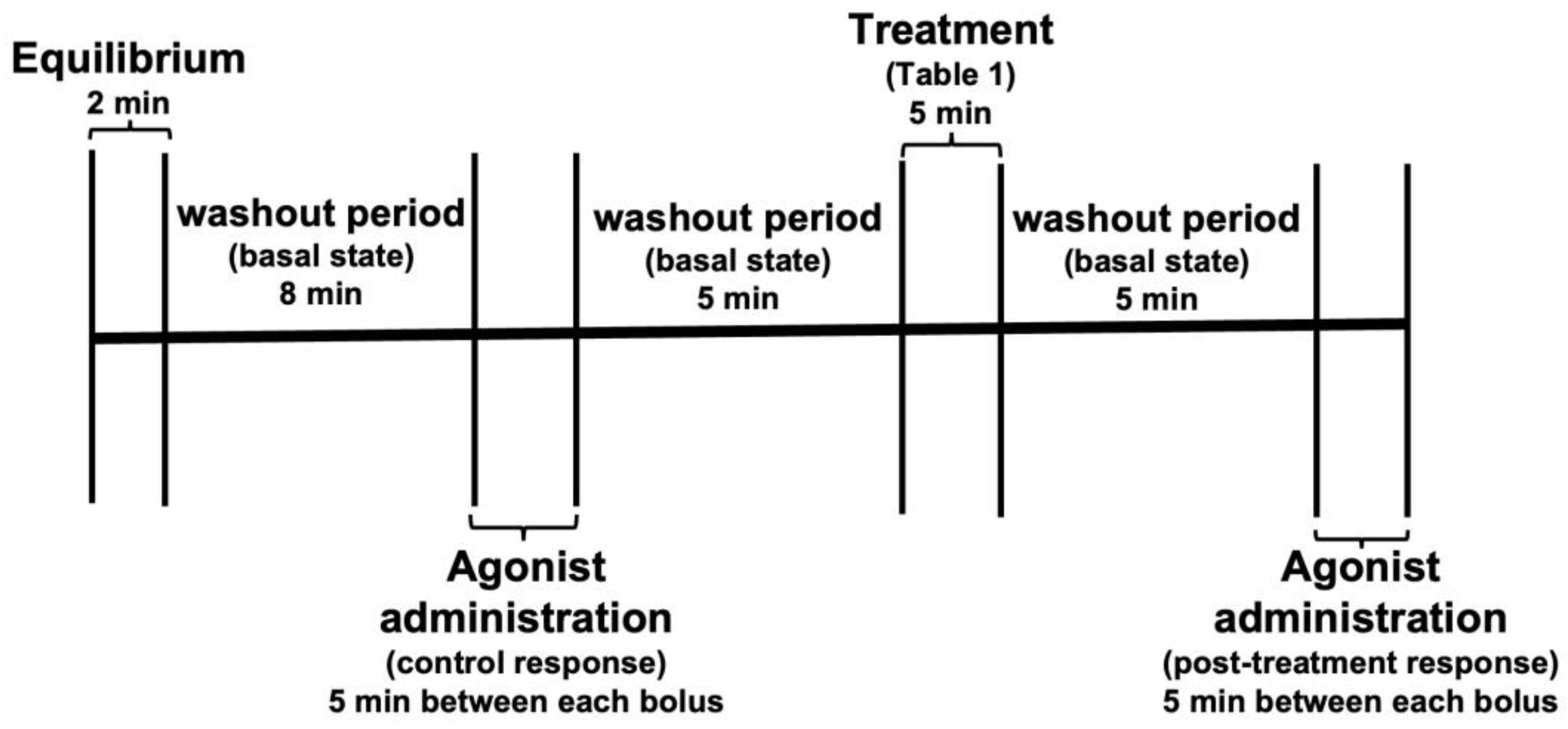
2.5. Isolated Heart Model
2.6. Coronary Perfusion Measurement
2.7. Measurement of Left Ventricular Contraction
2.8. Evaluation of Vascular and Inotropic Responses Induced by Intracoronary Administration of Acetylcholine, Phenylephrine and Angiotensin II
2.9. Statistical Analysis
3. Results
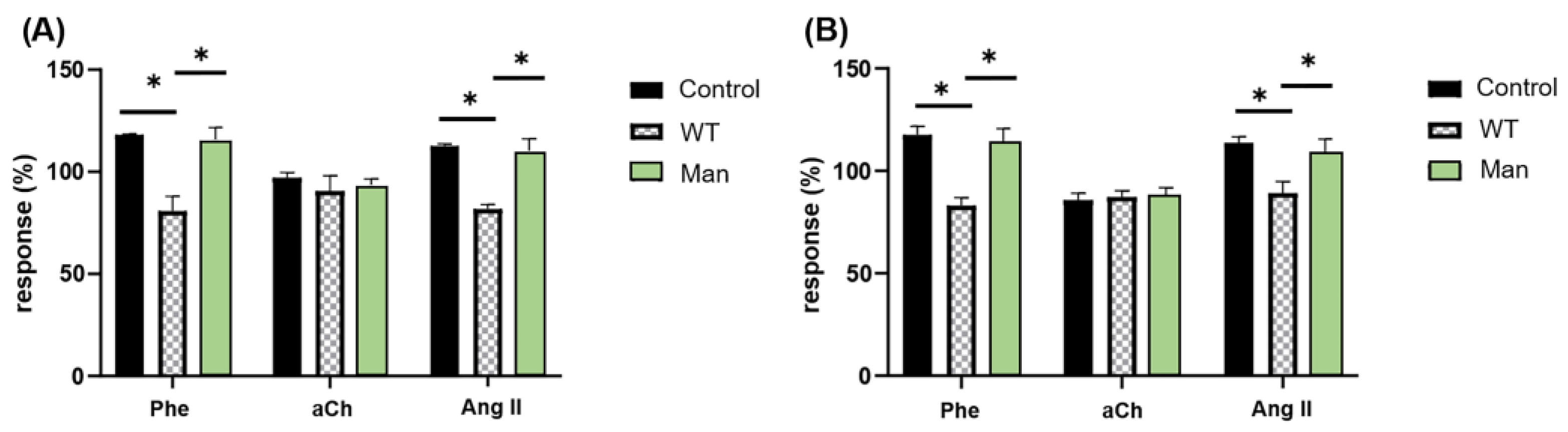
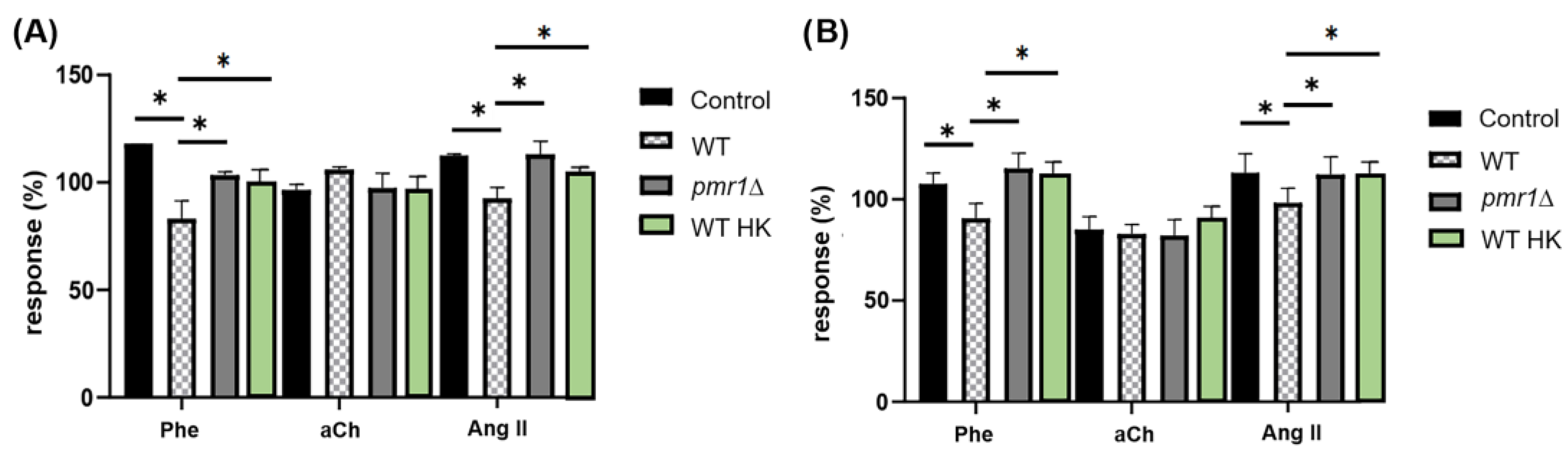
3.1. C. albicans Inhibits the Cardiac Response to Phe and Ang II but Not to aCh in a Mannose-Dependent Way
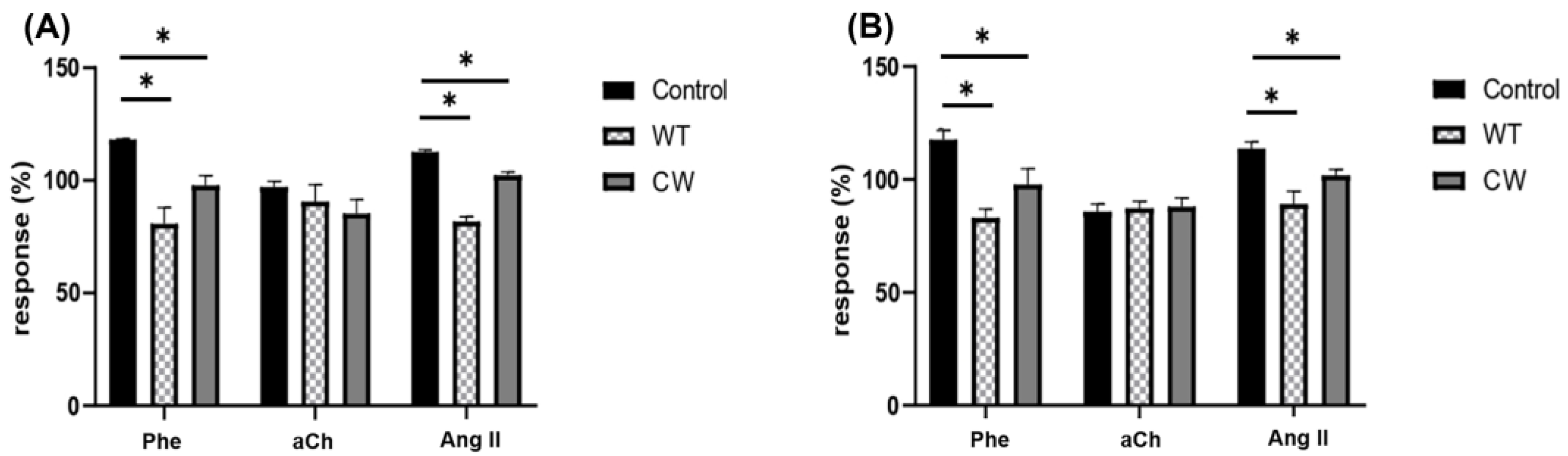
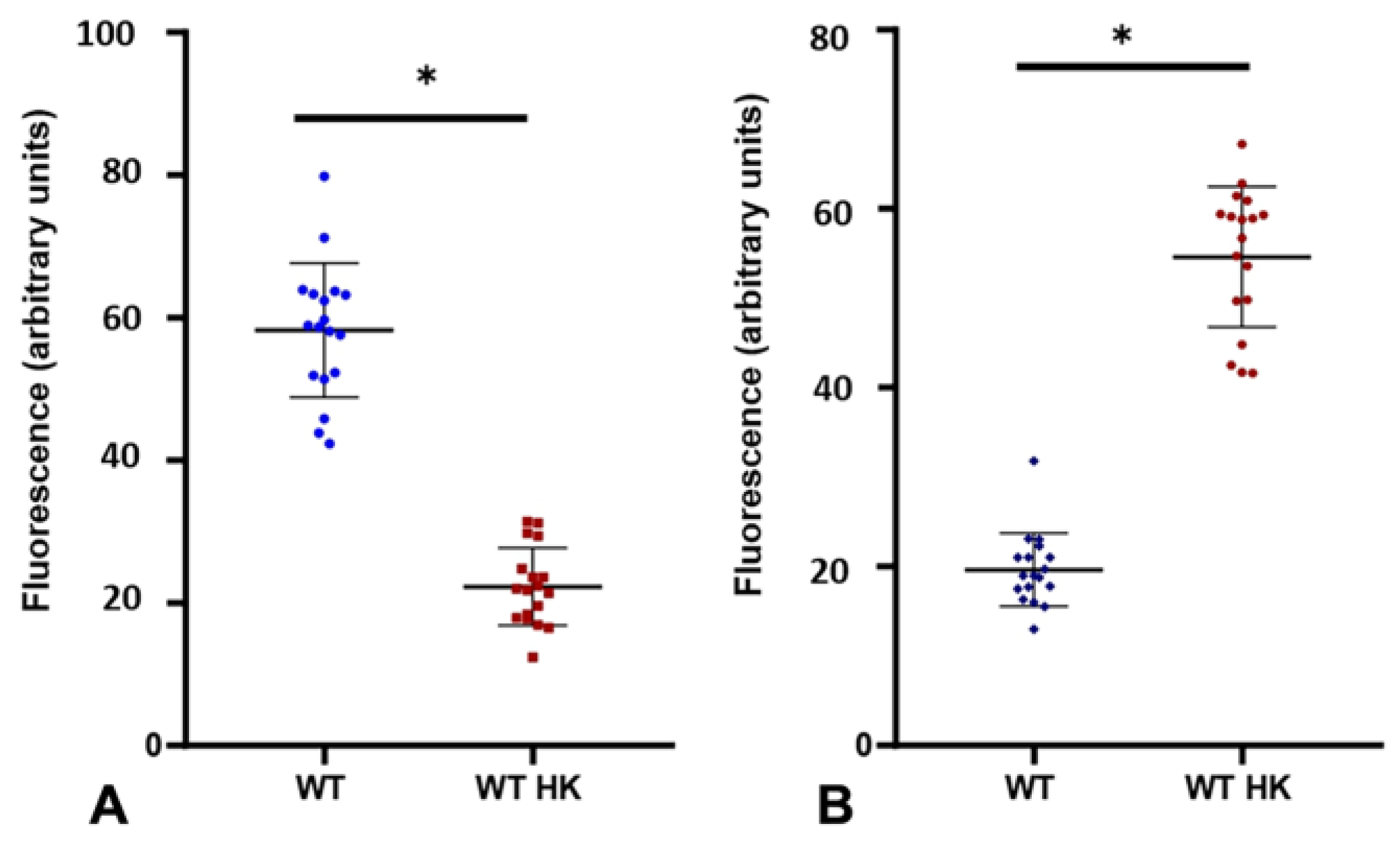
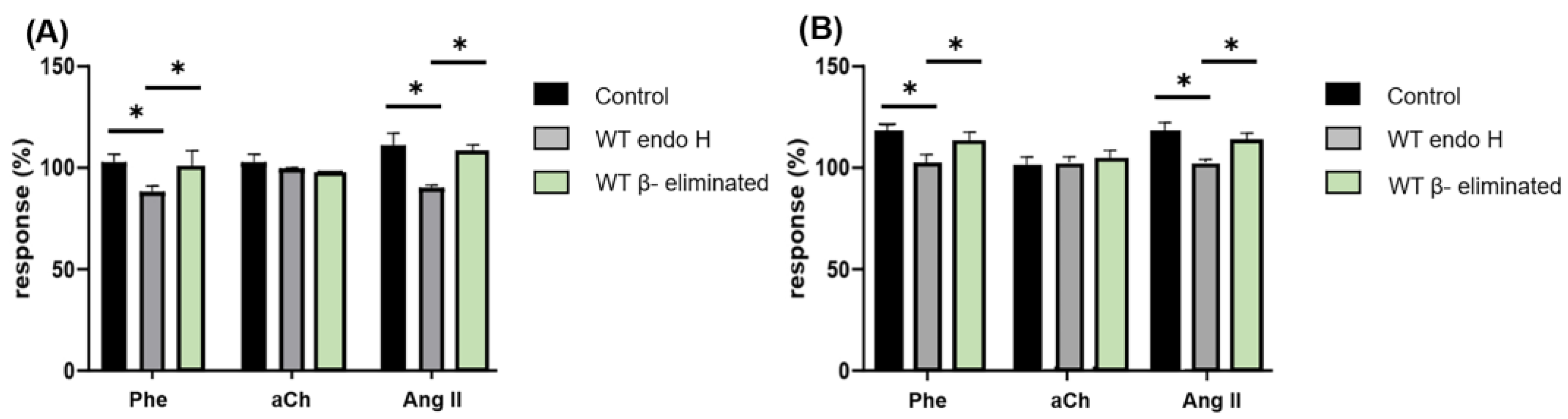
3.2. The Interaction of C. albicans with Specific GPCRs Is Due to the Cell Wall Structure and Not to Secreted Components
3.3. Proper C. albicans Glycosylation Is Required for Its Interaction with Specific GPCRs of the Coronary Endothelium
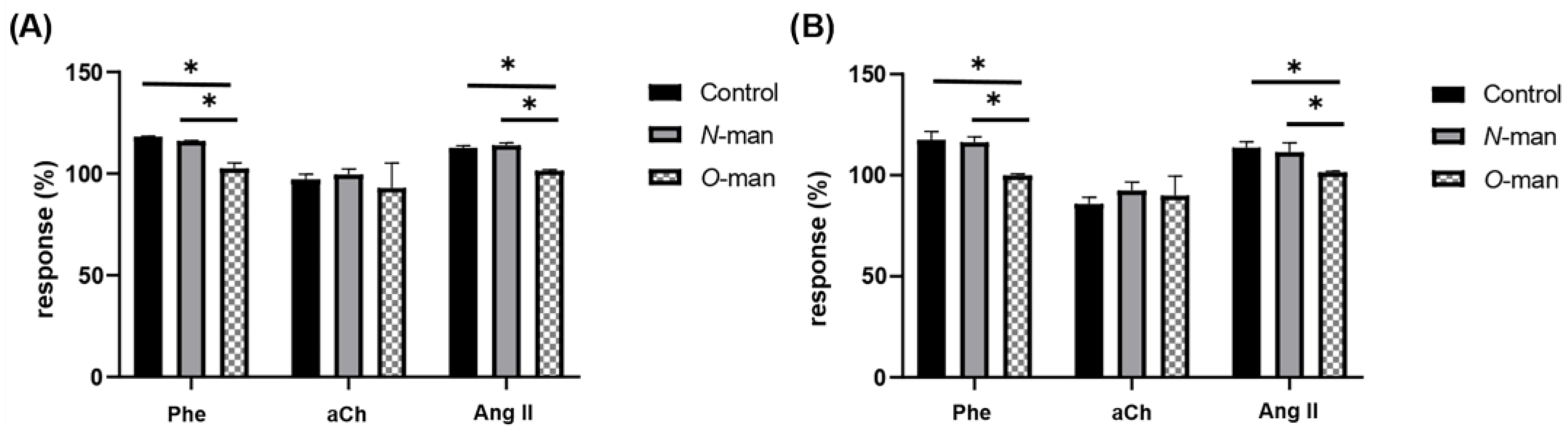
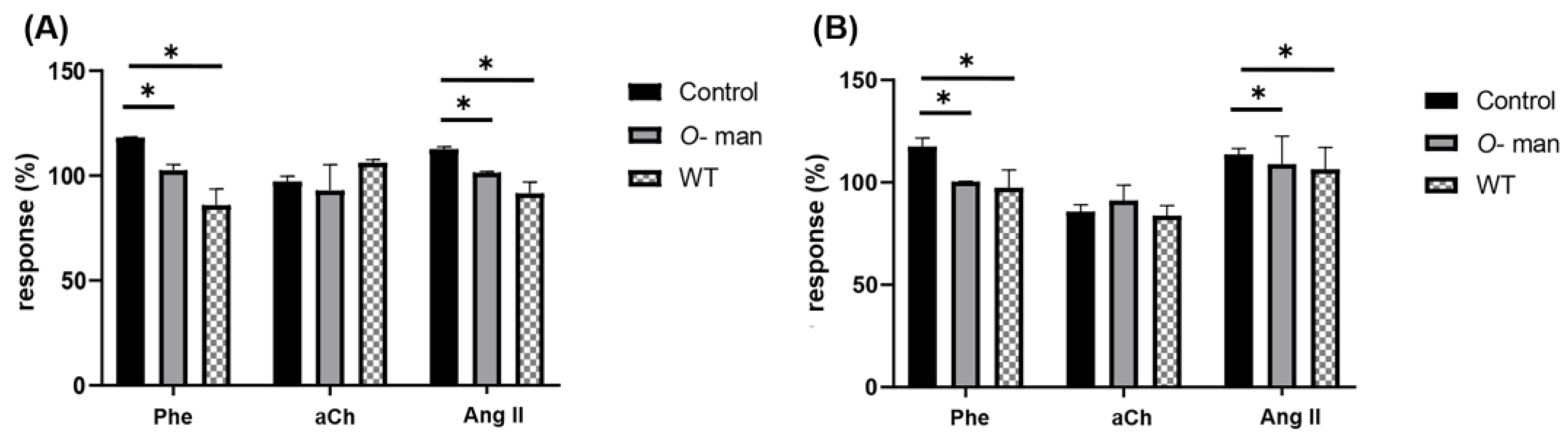
3.4. O-Linked Mannans from C. albicans Cell Wall Inhibit the Agonist-Mediated Response of GPCRs in the Coronary Endothelium
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Magill, S.S.; Edwards, J.R.; Bamberg, W.; Beldavs, Z.G.; Dumyati, G.; Kainer, M.A.; Lynfield, R.; Maloney, M.; McAllister-Hollod, L.; Nadle, J.; et al. Multistate Point-Prevalence Survey of Health Care-Associated Infections. N. Engl. J. Med. 2014, 370, 1198–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dongari-Bagtzoglou, A.; Dwivedi, P.; Ioannidou, E.; Shaqman, M.; Hull, D.; Burleson, J. Oral Candida Infection and Colonization in Solid Organ Transplant Recipients. Oral Microbiol. Immunol. 2009, 24, 249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaller, M.A.; Diekema, D.J. Epidemiology of Invasive Candidiasis: A Persistent Public Health Problem. Clin. Microbiol. Rev. 2007, 20, 133–163. [Google Scholar] [CrossRef] [Green Version]
- Bongomin, F.; Gago, S.; Oladele, R.O.; Denning, D.W. Global and Multi-National Prevalence of Fungal Diseases—Estimate Precision. J. Fungi 2017, 3, 57. [Google Scholar] [CrossRef]
- Pappas, P.G.; Lionakis, M.S.; Arendrup, M.C.; Ostrosky-Zeichner, L.; Kullberg, B.J. Invasive Candidiasis. Nat. Rev. Dis. Prim. 2018, 4, 18026. [Google Scholar] [CrossRef]
- Gow, N.A.R.; van de Veerdonk, F.L.; Brown, A.J.P.; Netea, M.G. Candida Albicans Morphogenesis and Host Defence: Discriminating Invasion from Colonization. Nat. Rev. Microbiol. 2012, 10, 112–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aspesberro, F.; Beghetti, M.; Oberhänsli, I.; Friedli, B. Fungal Endocarditis in Critically Ill Children. Eur. J. Pediatr. 1999, 158, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.T.; Hsieh, K.S.; Chen, Y.S.; Huang, I.F.; Cheng, M.F. Infective Endocarditis in Children without Underlying Heart Disease. J. Microbiol. Immunol. Infect. 2013, 46, 121–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marom, D.; Ashkenazi, S.; Samra, Z.; Birk, E. Infective Endocarditis in Previously Healthy Children with Structurally Normal Hearts. Pediatr. Cardiol. 2013, 34, 1415–1421. [Google Scholar] [CrossRef] [PubMed]
- Mullick, A.; Leon, Z.; Min-Oo, G.; Berghout, J.; Lo, R.; Daniels, E.; Gros, P. Cardiac Failure in C5-Deficient A/J Mice after Candida Albicans Infection. Infect. Immun. 2006, 74, 4439. [Google Scholar] [CrossRef]
- McCarty, T.P.; Pappas, P.G. Invasive Candidiasis. Infect. Dis. Clin. N. Am. 2016, 30, 103–124. [Google Scholar] [CrossRef] [PubMed]
- Kullberg, B.J.; Arendrup, M.C. Invasive Candidiasis. N. Engl. J. Med. 2015, 373, 1445–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gulati, M.; Nobile, C.J. Candida Albicans Biofilms: Development, Regulation, and Molecular Mechanisms. Microbes Infect. 2016, 18, 310. [Google Scholar] [CrossRef] [Green Version]
- Badiee, P.; Amirghofran, A.A.; Ghazi Nour, M.; Shafa, M.; Nemati, M.H. Incidence and Outcome of Documented Fungal Endocarditis. Int. Cardiovasc. Res. J. 2014, 8, 152–155. [Google Scholar] [PubMed]
- Mullick, A.; Elias, M.; Picard, S.; Bourget, L.; Jovcevski, O.; Gauthier, S.; Tuite, A.; Harakidas, P.; Bihun, C.; Massie, B.; et al. Dysregulated Inflammatory Response to Candida Albicans in a C5-Deficient Mouse Strain. Infect. Immun. 2004, 72, 5868–5876. [Google Scholar] [CrossRef] [Green Version]
- Yuan, S.M. Fungal Endocarditis. Braz. J. Cardiovasc. Surg. 2016, 31, 252–255. [Google Scholar] [CrossRef]
- Mora-Montes, H.M.; Ponce-Noyola, P.; Villagómez-Castro, J.C.; Gow, N.A.R.; Flores-Carreón, A.; López-Romero, E. Protein Glycosylation in Candida. Future Microbiol. 2009, 4, 1167–1183. [Google Scholar] [CrossRef]
- Grubb, S.E.W.; Murdoch, C.; Sudbery, P.E.; Saville, S.P.; Lopez-Ribot, J.L.; Thornhill, M.H. Adhesion of Candida Albicans to Endothelial Cells under Physiological Conditions of Flow. Infect. Immun. 2009, 77, 3872–3878. [Google Scholar] [CrossRef] [Green Version]
- Gow, N.A.R.; Latge, J.-P.; Munro, C.A. The Fungal Cell Wall: Structure, Biosynthesis, and Function. Microbiol. Spectr. 2017, 5, 28513415. [Google Scholar] [CrossRef] [Green Version]
- Hoyer, L.L.; Cota, E. Candida Albicans Agglutinin-like Sequence (Als) Family Vignettes: A Review of Als Protein Structure and Function. Front. Microbiol. 2016, 7, 280. [Google Scholar] [CrossRef]
- Sheppard, D.C.; Yeaman, M.R.; Welch, W.H.; Phan, Q.T.; Fu, Y.; Ibrahim, A.S.; Filler, S.G.; Zhang, M.; Waring, A.J.; Edwards, J.E. Functional and Structural Diversity in the Als Protein Family of Candida Albicans. J. Biol. Chem. 2004, 279, 30480–30489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klotz, S.A. Fungal Adherence to the Vascular Compartment: A Critical Step in the Pathogenesis of Disseminated Candidiasis. Clin. Infect. Dis. 1992, 14, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Barajas-Espinosa, A.; Ramiro-Diaz, J.; Briones-Cerecero, E.; Chi-Ahumada, E.; de La Rosa, A.B.; Arroyo-Flores, B.; Rubio, R. Involvement of Endothelial Man and Gal-Binding Lectins in Sensing the Flow in Coronary Arteries. Front. Biosci. 2008, 13, 5421–5431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Aguilar, S.; Torres-Tirado, D.; Martell-Gallegos, G.; Velarde-Salcedo, J.; Paulina Barba-de la Rosa, A.; Knabb, M.; Rubio, R. G Protein-Coupled Receptors Mediate Coronary Flow-and Agonist-Induced Responses via Lectin-Oligosaccharide Interactions. Am. J. Physiol. Heart Circ. Physiol. 2014, 306, 699–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubio-Gayosso, I.; Barajas-Espinosa, A.; Castillo-Hernandez, J.; Ramiro-Diaz, J.; Ceballos, G.; Rubio, R. Enzymatic Hydrolysis of Luminal Coronary Glycosidic Structures Uncovers Their Role in Sensing Coronary Flow. Front. Biosci. 2005, 10, 1050–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramiro-Diaz, J.; Barajas-Espinosa, A.; Chi-Ahumada, E.; Perez-Aguilar, S.; Torres-Tirado, D.; Castillo-Hernandez, J.; Knabb, M.; de La Rosa, A.B.; Rubio, R. Luminal Endothelial Lectins with Affinity for N-Acetylglucosamine Determine Flow-Induced Cardiac and Vascular Paracrine-Dependent Responses. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, 743–751. [Google Scholar] [CrossRef] [Green Version]
- Bates, S.; MacCallum, D.M.; Bertram, G.; Munro, C.A.; Hughes, H.B.; Buurman, E.T.; Brown, A.J.P.; Odds, F.C.; Gow, N.A.R. Candida Albicans Pmr1p, a Secretory Pathway P-Type Ca2+/Mn2+-ATPase, Is Required for Glycosylation and Virulence. J. Biol. Chem. 2005, 280, 23408–23415. [Google Scholar] [CrossRef] [Green Version]
- Brand, A.; MacCallum, D.M.; Brown, A.J.P.; Gow, N.A.R.; Odds, F.C. Ectopic Expression of URA3 Can Influence the Virulence Phenotypes and Proteome of Candida Albicans but Can Be Overcome by Targeted Reintegration of URA3 at the RPS10 Locus. Eukaryot. Cell 2004, 3, 900–909. [Google Scholar] [CrossRef] [Green Version]
- Pérez-García, L.A.; Csonka, K.; Flores-Carreón, A.; Estrada-Mata, E.; Mellado-Mojica, E.; Németh, T.; López-Ramírez, L.A.; Toth, R.; López, M.G.; Vizler, C.; et al. Role of Protein Glycosylation in Candida Parapsilosis Cell Wall Integrity and Host Interaction. Front. Microbiol. 2016, 7, 306. [Google Scholar] [CrossRef] [Green Version]
- Mora-Montes, H.M.; Bates, S.; Netea, M.G.; Díaz-Jiménez, D.F.; López-Romero, E.; Zinker, S.; Ponce-Noyola, P.; Kullberg, B.J.; Brown, A.J.P.; Odds, F.C.; et al. Endoplasmic Reticulum α-Glycosidases of Candida Albicans Are Required for N Glycosylation, Cell Wall Integrity, and Normal Host-Fungus Interaction. Eukaryot. Cell 2007, 6, 2184–2193. [Google Scholar] [CrossRef]
- Mora-Montes, H.M.; McKenzie, C.; Bain, J.M.; Lewis, L.E.; Erwig, L.P.; Gow, N.A.R. Interactions Between Macrophages and Cell Wall Oligosaccharides of Candida albicans. In Host-Fungus Interactions; Methods and Protocols; Brand, A.C., MacCallum, D.M., Eds.; Humana Press: Totowa, NJ, USA, 2012; pp. 247–260. ISBN 9781617795398. [Google Scholar]
- Díaz-Jiménez, D.F.; Mora-Montes, H.M.; Hernández-Cervantes, A.; Luna-Arias, J.P.; Gow, N.A.R.; Flores-Carreón, A. Biochemical Characterization of Recombinant Candida Albicans Mannosyltransferases Mnt1, Mnt2 and Mnt5 Reveals New Functions in O- and N-Mannan Biosynthesis. Biochem. Biophys. Res. Commun. 2012, 419, 77–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kocourek, J.; Ballou, C.E. Method for Fingerprinting Yeast Cell Wall Mannans. J. Bacteriol. 1969, 100, 1175–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, R.M.; Mocanu, M.M.; Yellon, D.M. Retrograde Heart Perfusion: The Langendorff Technique of Isolated Heart Perfusion. J. Mol. Cell. Cardiol. 2011, 50, 940–950. [Google Scholar] [CrossRef] [PubMed]
- Liao, R.; Podesser, B.K.; Lim, C.C. The Continuing Evolution of the Langendorff and Ejecting Murine Heart: New Advances in Cardiac Phenotyping. Am. J. Physiol. Heart Circ. Physiol. 2012, 303, 156–167. [Google Scholar] [CrossRef]
- NOM-062-ZOO-1999; Especificaciones Técnicas Para la Producción, Cuidado y Uso de Los Animales de Laboratorio. Diario Oficial de la Federación Norma Oficial Mexicana: Mexico City, Mexico, 1999.
- Torres-Tirado, D.; Knabb, M.; Castaño, I.; Patrón-Soberano, A.; De Las Peñas, A.; Rubio, R. Candida Glabrata Binds to Glycosylated and Lectinic Receptors on the Coronary Endothelial Luminal Membrane and Inhibits Flow Sense and Cardiac Responses to Agonists. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 310, R24–R32. [Google Scholar] [CrossRef] [Green Version]
- Bojar, D.; Meche, L.; Meng, G.; Eng, W.; Smith, D.F.; Cummings, R.D.; Mahal, L.K. A Useful Guide to Lectin Binding: Machine-Learning Directed Annotation of 57 Unique Lectin Specificities. ACS Chem. Biol. 2021, 17, 2993–3012. [Google Scholar] [CrossRef]
- Gow, N.A.R.; Yadav, B. Microbe Profile: Candida Albicans: A Shape-Changing, Opportunistic Pathogenic Fungus of Humans. Microbiology 2017, 163, 1145–1147. [Google Scholar] [CrossRef]
- Kojic, E.M.; Darouiche, R.O. Candida Infections of Medical Devices. Clin. Microbiol. Rev. 2004, 17, 255. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.J.; Liu, W.L.; Lin, H.L.; Lai, C.C. Clinical Manifestations and Prognostic Factors of Central Line-Associated Candidemia. Am. J. Infect. Control 2014, 42, 1238–1240. [Google Scholar] [CrossRef]
- Sheppard, D.C.; Filler, S.G. Host Cell Invasion by Medically Important Fungi. Cold Spring Harb. Perspect. Med. 2015, 5, a019687. [Google Scholar] [CrossRef]
- Bojang, E.; Ghuman, H.; Kumwenda, P.; Hall, R.A. Immune Sensing of Candida Albicans. J. Fungi 2021, 7, 119. [Google Scholar] [CrossRef]
- Lin, B.; Qing, X.; Liao, J.; Zhuo, K. Role of Protein Glycosylation in Host-Pathogen Interaction. Cells 2020, 9, 1022. [Google Scholar] [CrossRef] [PubMed]
- Scheld, W.M.; Strunk, R.W.; Balian, G.; Calderone, R.A. Microbial Adhesion to Fibronectin in Vitro Correlates with Production of Endocarditis in Rabbits. Proc. Soc. Exp. Biol. Med. 1985, 180, 474–482. [Google Scholar] [CrossRef] [PubMed]
- Klotz, S.A.; Smith, R.L.; Stewart, B.W. Effect of an Arginine-Glycine-Aspartic Acid-Containing Peptide on Hematogenous Candidal Infections in Rabbits. Antimicrob. Agents Chemother. 1992, 36, 132–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukazawa, Y.; Kagaya, K. Molecular Bases of Adhesion of Candida Albicans. J. Med. Vet. Mycol. 1997, 35, 87–99. [Google Scholar] [CrossRef] [Green Version]
- Munro, C.A.; Bates, S.; Buurman, E.T.; Hughes, H.B.; MacCallum, D.M.; Bertram, G.; Atrih, A.; Ferguson, M.A.J.; Bain, J.M.; Brand, A.; et al. Mnt1p and Mnt2p of Candida Albicans Are Partially Redundant α-1,2-Mannosyltransferases That Participate in O-Linked Mannosylation and Are Required for Adhesion and Virulence. J. Biol. Chem. 2005, 280, 1051–1060. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Description | Reference |
|---|---|---|
| C. albicans WT (NGY-152) (3.7 × 103 cel/mL) | ura3Δ::imm434/ura3Δ::imm434, RPS1/rps1Δ::CIp10. It behaves like the wild-type strain | [28] |
| C. albicans WT HK (3.7 × 103 cel/mL) | ura3Δ::imm434/ura3Δ::imm434, RPS1/rps1Δ::CIp10. It behaves like the wild-type strain and it was heat-inactivated | [28] |
| C. albicans pmr1Δ (3.7 × 103 cel/mL) | ura3Δ::imm434/ura3Δ::imm434; pmr1Δ::hisG/pmr1Δ::hisG, RPS1/rps1Δ::CIp10. It has truncated both N-linked and O-linked mannans. | [27] |
| C. albicans without N-linked glycans | C. albicans WT (NGY-152) treated with endo H | [31] |
| C. albicans without O-linked glycans | C. albicans WT (NGY-152) β-eliminated | [32] |
| N-linked glycan (10 mM) | Glycans recovered and purified from C. albicans WT trimming with Endo H | [31,33] |
| O-linked glycan (10 mM) | Glycans recovered and purified from β-elimination of C. albicans WT | [32,33] |
| Cell wall from C. albicans WT (1 mg/mL) | Purified yeast cell walls of C. albicans WT (NGY-152) | [30] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ocaña-Ortega, A.; Pérez-Flores, G.; Torres-Tirado, D.; Pérez-García, L.A. O-Linked Glycans of Candida albicans Interact with Specific GPCRs in the Coronary Endothelium and Inhibit the Cardiac Response to Agonists. J. Fungi 2023, 9, 141. https://doi.org/10.3390/jof9020141
Ocaña-Ortega A, Pérez-Flores G, Torres-Tirado D, Pérez-García LA. O-Linked Glycans of Candida albicans Interact with Specific GPCRs in the Coronary Endothelium and Inhibit the Cardiac Response to Agonists. Journal of Fungi. 2023; 9(2):141. https://doi.org/10.3390/jof9020141
Chicago/Turabian StyleOcaña-Ortega, Alejandro, Gabriela Pérez-Flores, David Torres-Tirado, and Luis A. Pérez-García. 2023. "O-Linked Glycans of Candida albicans Interact with Specific GPCRs in the Coronary Endothelium and Inhibit the Cardiac Response to Agonists" Journal of Fungi 9, no. 2: 141. https://doi.org/10.3390/jof9020141
APA StyleOcaña-Ortega, A., Pérez-Flores, G., Torres-Tirado, D., & Pérez-García, L. A. (2023). O-Linked Glycans of Candida albicans Interact with Specific GPCRs in the Coronary Endothelium and Inhibit the Cardiac Response to Agonists. Journal of Fungi, 9(2), 141. https://doi.org/10.3390/jof9020141