
Applications of Diacylglycerol Acyltransferase for Triacylglycerol Production in Mortierella alpina

Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Plasmids
2.2. Sequence Alignment, Cloning, and Construction of Homologous Overexpression Vector for MaDGATs of M. alpina
2.3. Culture Conditions of M. alpina
2.4. A. Tumefaciens-Mediated Transformation
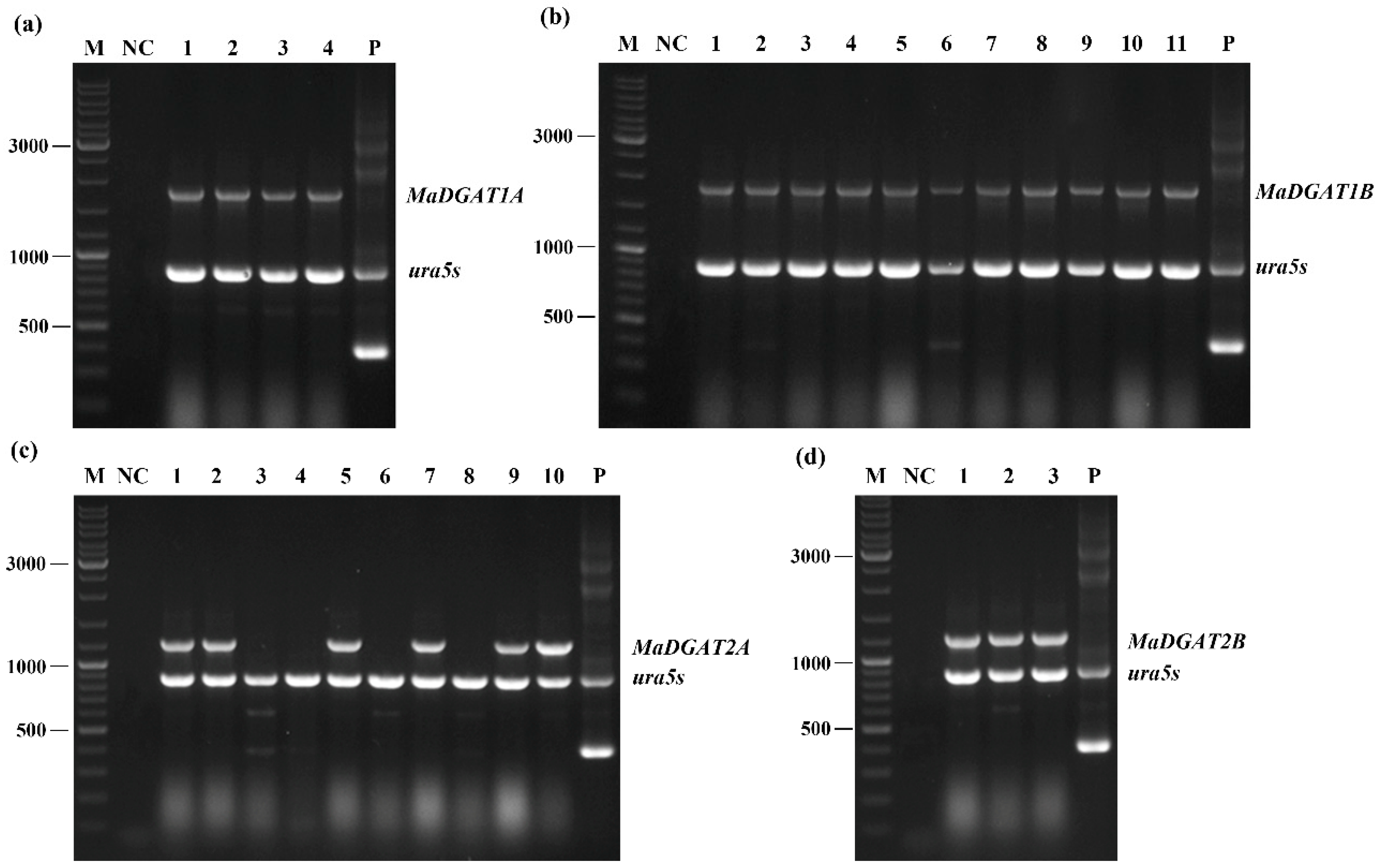
2.5. Extraction of Genomic DNA and Identification of Transformants
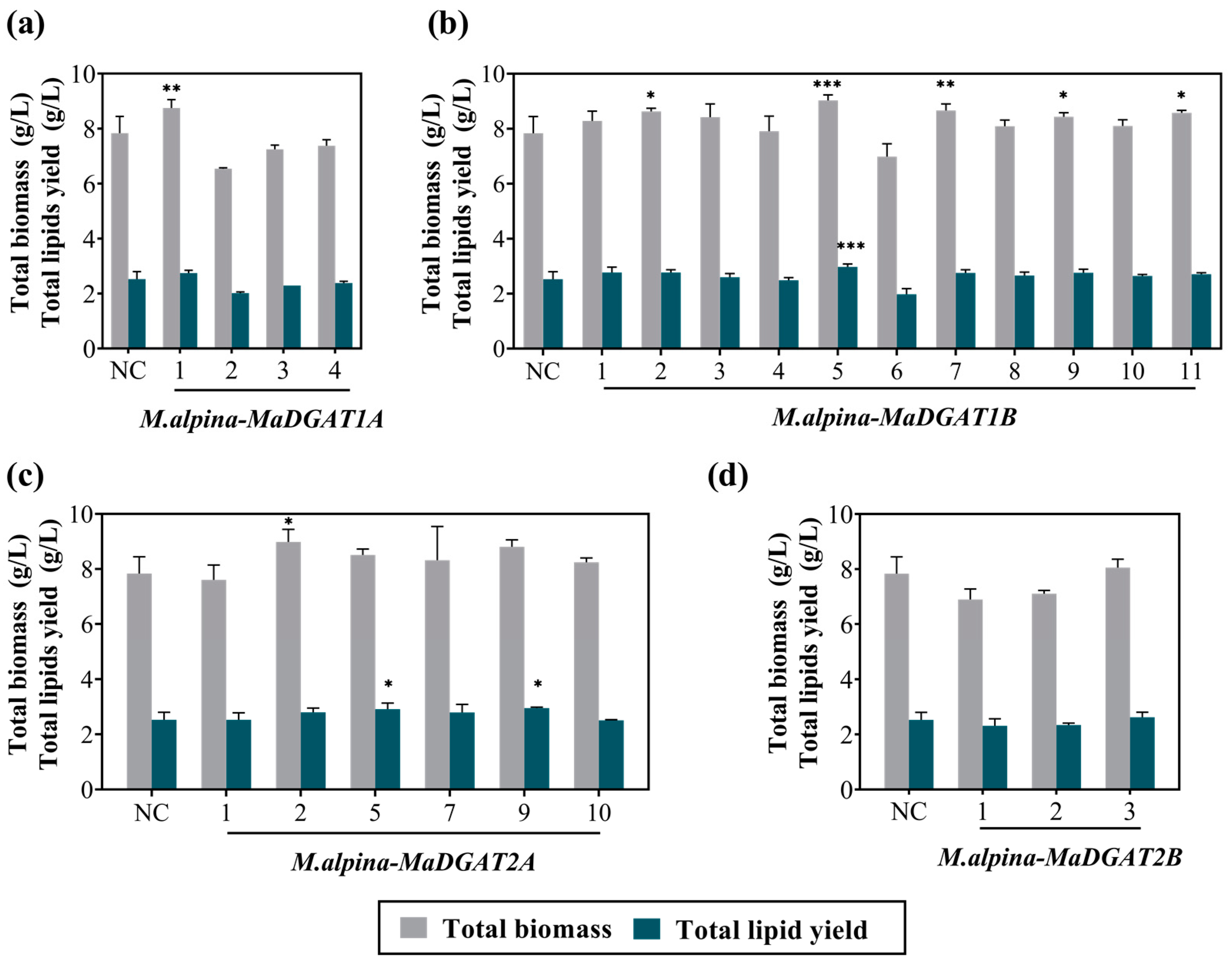
2.6. Biomass Analysis, Lipid Extraction, and Targeted Analysis of FA Profiles
2.7. Untargeted Lipidomics Analysis
2.8. Statistical Analysis
3. Results
3.1. Identification and Screening of the M. alpina-MaDGAT1A/1B/2A/2B Recombinant Strains
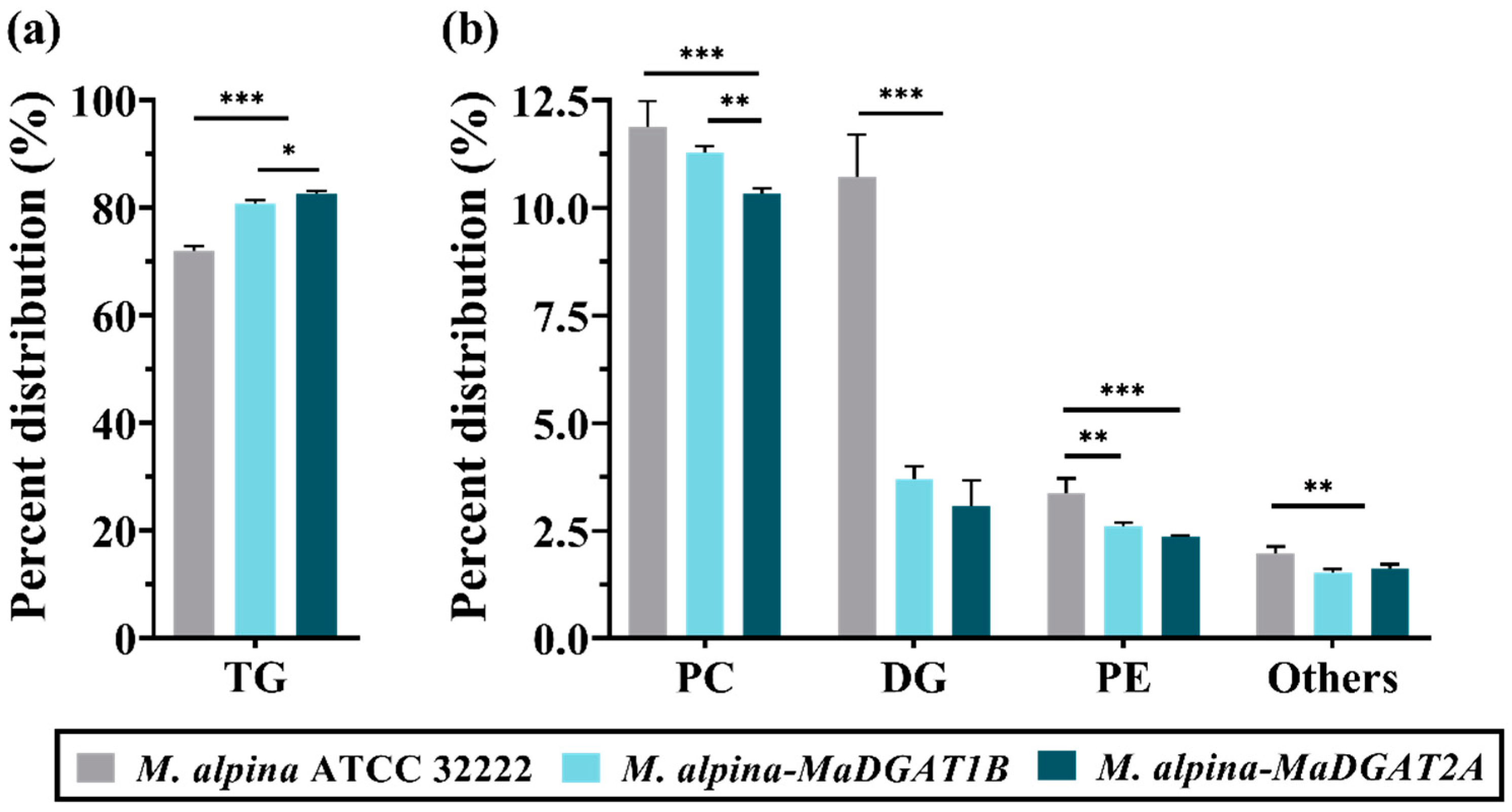
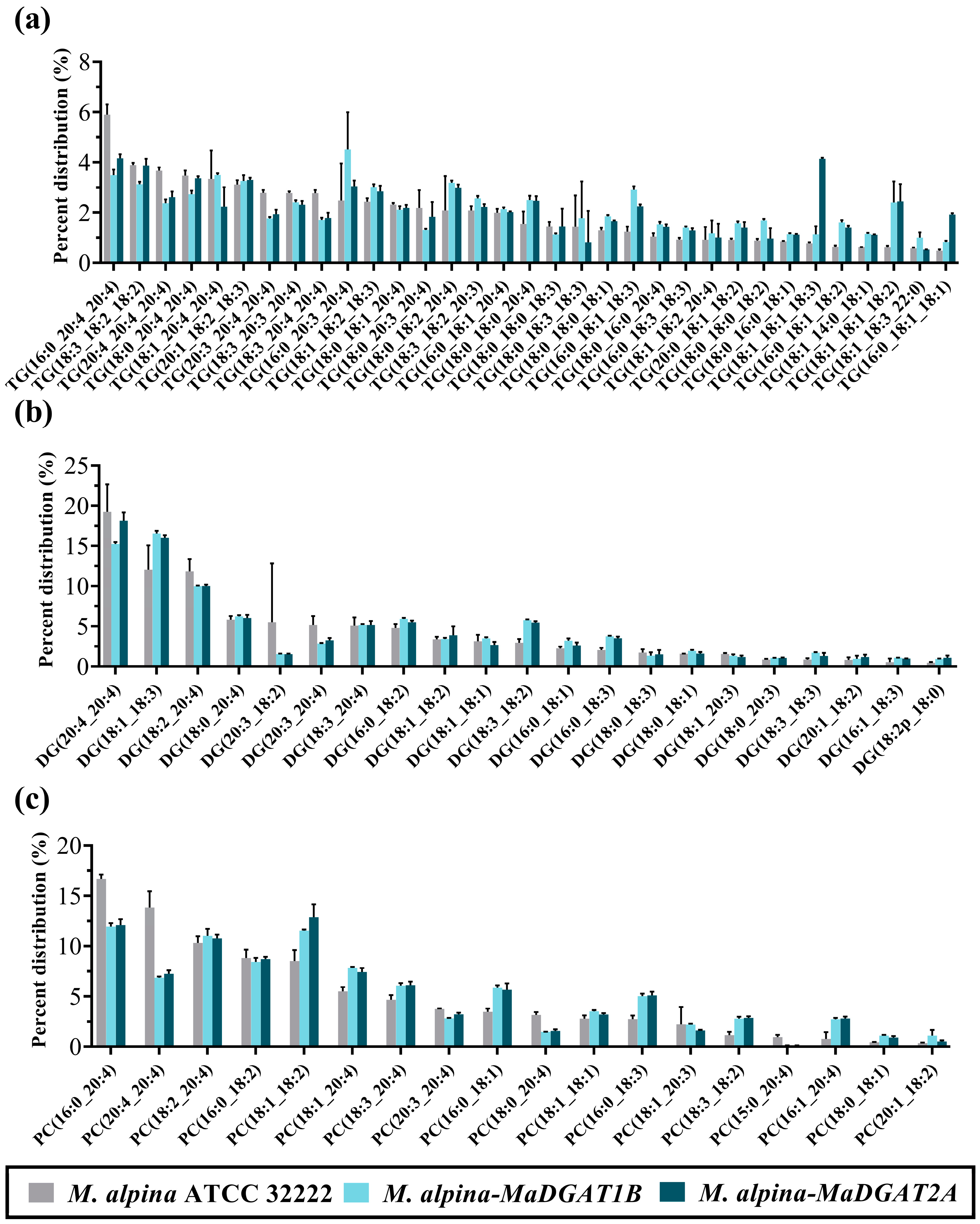
3.2. Lipidomics Profiles of M. alpina-MaDGAT1B and M. alpina-MaDGAT2A
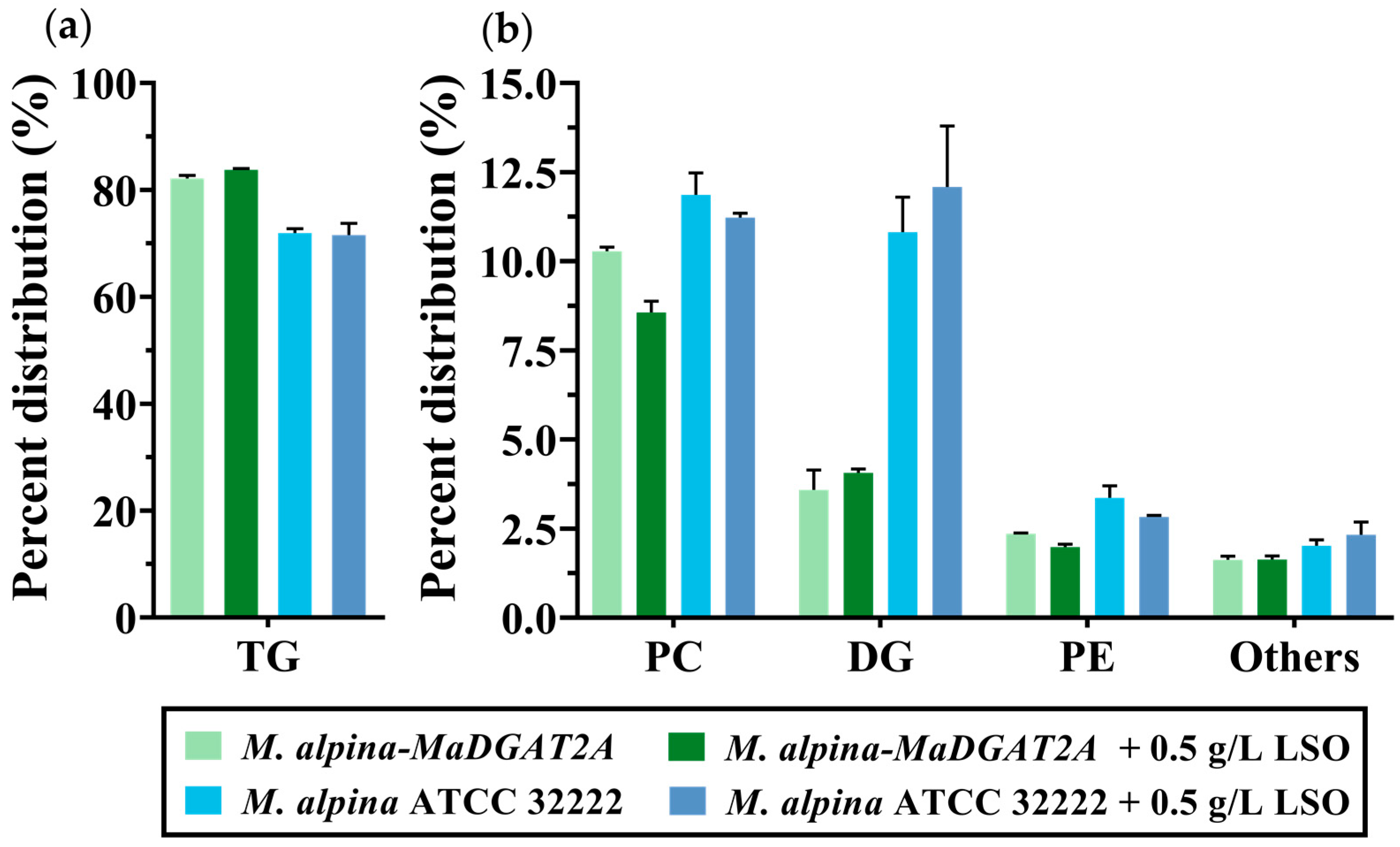
3.3. Effect of LSO Supplementation on TG Production of M. alpina-MaDGAT2A
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xu, C.; Shanklin, J. Triacylglycerol Metabolism, Function, and Accumulation in Plant Vegetative Tissues. Annu. Rev. Plant Biol. 2016, 67, 179–206. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Ambigaipalan, P. Omega-3 Polyunsaturated Fatty Acids and Their Health Benefits. Annu. Rev. Food Sci. Technol. 2018, 9, 345–381. [Google Scholar] [CrossRef] [PubMed]
- Harris, W.S.; Tintle, N.L.; Imamura, F.; Qian, F.; Korat, A.V.A.; Marklund, M.; Djousse, L.; Bassett, J.K.; Carmichael, P.H.; Chen, Y.Y.; et al. Blood n-3 fatty acid levels and total and cause-specific mortality from 17 prospective studies. Nat. Commun. 2021, 12, 2329. [Google Scholar] [CrossRef]
- Bhatt, R.D.; Libby, P.; Verma, S.; Mason, R.P.; Bhatt, D.L. The role of eicosapentaenoic acid in reducing important cardiovascular events, including coronary revascularization. Prog. Cardiovasc. Dis. 2021, 69, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Ratledge, C. Fatty acid biosynthesis in microorganisms being used for Single Cell Oil production. Biochimie 2004, 86, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Ratledge, C. Single Cell Oils for the 21st Century. In Single Cell Oils, 8th ed.; Cohen, Z., Ratledge, C., Eds.; AOCS Press: Urbana, IL, USA, 2010; pp. 3–26. [Google Scholar]
- Chang, L.; Lu, H.; Chen, H.; Tang, X.; Zhao, J.; Zhang, H.; Chen, Y.Q.; Chen, W. Lipid metabolism research in oleaginous fungus Mortierella alpina: Current progress and future prospects. Biotechnol. Adv. 2022, 54, 107794. [Google Scholar] [CrossRef]
- Tang, X.; Chen, H.; Mei, T.; Ge, C.; Gu, Z.; Zhang, H.; Chen, Y.Q.; Chen, W. Characterization of an Omega-3 Desaturase From Phytophthora parasitica and Application for Eicosapentaenoic Acid Production in Mortierella alpina. Front. Microbiol. 2018, 9, 01878. [Google Scholar] [CrossRef]
- Coleman, R. Enzymes of triacylglycerol synthesis and their regulation. Prog. Lipid Res. 2004, 43, 134–176. [Google Scholar] [CrossRef]
- Cao, H. Structure-Function Analysis of Diacylglycerol Acyltransferase Sequences from 70 Organisms. BMC Res. Notes 2011, 4, 249. [Google Scholar] [CrossRef]
- Turchetto-Zolet, A.C.; Maraschin, F.S.; de Morais, G.L.; Cagliari, A.; Andrade, C.M.; Margis-Pinheiro, M.; Margis, R. Evolutionary view of acyl-CoA diacylglycerol acyltransferase (DGAT), a key enzyme in neutral lipid biosynthesis. BMC Evol. Biol. 2011, 11, 263. [Google Scholar] [CrossRef] [Green Version]
- Jeennor, S.; Veerana, M.; Anantayanon, J.; Panchanawaporn, S.; Chutrakul, C.; Laoteng, K. Diacylglycerol acyltransferase 2 of Mortierella alpina with specificity on long-chain polyunsaturated fatty acids: A potential tool for reconstituting lipids with nutritional value. J. Biotechnol. 2017, 263, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Cao, J.; Chen, H.; Tang, X.; Zhang, H.; Chen, W. Functional characterization of two diacylglycerol acyltransferase 1 genes in Mortierella alpina. Lett. Appl. Microbiol. 2022, 74, 194–203. [Google Scholar] [CrossRef]
- Xue, Z.; Yadav Narendra, S.; Macool Daniel, J.; Macool Daniel, J. Mortierella alpina Diacylglycerol Acyltransferase for Alteration of Polyunsaturated Fatty Acids and Oil Content in Oleaginous Organisms. U.S. Patent US-7198937-B2, 3 April 2007. [Google Scholar]
- Lei, W.; Wei, C.; Yun, F.; Yan, R.; Zhennan, G.; Haiqin, C.; Hongchao, W.; Thomas, M.J.; Baixi, Z.; Berquin, I.M.; et al. Genome characterization of the oleaginous fungus Mortierella alpina. PLoS ONE 2011, 6, e28319. [Google Scholar]
- Lu, H.; Chen, H.; Tang, X.; Yang, Q.; Zhang, H.; Chen, Y.Q.; Chen, W. Ultra Performance Liquid Chromatography-Q Exactive Orbitrap/Mass Spectrometry-Based Lipidomics Reveals the Influence of Nitrogen Sources on Lipid Biosynthesis of Mortierella alpina. J. Agric. Food Chem. 2019, 67, 10984–10993. [Google Scholar] [CrossRef] [PubMed]
- Han, X. Lipidomics for studying metabolism. Nat. Rev. Endocrinol. 2016, 12, 668–679. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Zhang, H.; Hu, Z.; Chin, Y.; Li, G.; Huang, J.; Zhang, X.; Jiang, B.; Hu, Y. Differentiation of three commercial tuna species through Q-Exactive Orbitrap mass spectrometry based lipidomics and chemometrics. Food Res. Int. 2022, 158, 111509. [Google Scholar] [CrossRef]
- Barchuk, M.; Dutour, A.; Ancel, P.; Svilar, L.; Miksztowicz, V.; Lopez, G.; Rubio, M.; Schreier, L.; Nogueira, J.P.; Valéro, R.; et al. Untargeted Lipidomics Reveals a Specific Enrichment in Plasmalogens in Epicardial Adipose Tissue and a Specific Signature in Coronary Artery Disease. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 986–1000. [Google Scholar] [CrossRef]
- Wang, H.; Wu, Y.; Xiang, H.; Sun-Waterhouse, D.; Zhao, Y.; Chen, S.; Li, L.; Wang, Y. UHPLC-Q-Exactive Orbitrap MS/MS-based untargeted lipidomics reveals molecular mechanisms and metabolic pathways of lipid changes during golden pomfret (Trachinotus ovatus) fermentation. Food Chem. 2022, 396, 133676. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Sharpe, P.L.; Hong, S.-P.; Yadav, N.S.; Xie, D.; Short, D.R.; Damude, H.G.; Rupert, R.A.; Seip, J.E.; Wang, J.; et al. Production of omega-3 eicosapentaenoic acid by metabolic engineering of Yarrowia lipolytica. Nat. Biotechnol. 2013, 31, 734–740. [Google Scholar] [CrossRef]
- Jiang, W.; Li, C.; Li, Y.; Peng, H. Metabolic Engineering Strategies for Improved Lipid Production and Cellular Physiological Responses in Yeast Saccharomyces cerevisiae. J. Fungi 2022, 8, 427. [Google Scholar] [CrossRef]
- Zulu, N.N.; Popko, J.; Zienkiewicz, K.; Tarazona, P.; Herrfurth, C.; Feussner, I. Heterologous co-expression of a yeast diacylglycerol acyltransferase (ScDGA1) and a plant oleosin (AtOLEO3) as an efficient tool for enhancing triacylglycerol accumulation in the marine diatom Phaeodactylum tricornutum. Biotechnol. Biofuels 2017, 10, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.-W.; Cen, S.-Y.; Liu, Y.-H.; Balamurugan, S.; Zheng, X.-Y.; Alimujiang, A.; Yang, W.-D.; Liu, J.-S.; Li, H.-Y. A type 2 diacylglycerol acyltransferase accelerates the triacylglycerol biosynthesis in heterokont oleaginous microalga Nannochloropsis oceanica. J. Biotechnol. 2016, 229, 65–71. [Google Scholar] [CrossRef]
- Jang, H.-D.; Lin, Y.-Y.; Yang, S.-S. Effect of culture media and conditions on polyunsaturated fatty acids production by Mortierella alpina. Bioresour. Technol. 2005, 96, 1633–1644. [Google Scholar] [CrossRef] [PubMed]
- Guangfei, H.; Haiqin, C.; Lei, W.; Zhennan, G.; Yuanda, S.; Hao, Z.; Wei, C.; Yong, Q.C. Role of malic enzyme during fatty acid synthesis in the oleaginous fungus Mortierella alpina. Appl. Environ. Microbiol. 2014, 80, 2672–2678. [Google Scholar]
- Matyash, V.; Liebisch, G.; Kurzchalia, T.V.; Shevchenko, A.; Schwudke, D. Lipid extraction by methyl-tert-butyl ether for high-throughput lipidomics. J. Lipid Res. 2008, 49, 1137–1146. [Google Scholar] [CrossRef]
- Tu, J.; Yin, Y.; Xu, M.; Wang, R.; Zhu, Z.-J. Absolute quantitative lipidomics reveals lipidome-wide alterations in aging brain. Metabolomics 2018, 14, 5. [Google Scholar] [CrossRef]
- Ando, A.; Sumida, Y.; Negoro, H.; Suroto, D.A.; Ogawa, J.; Sakuradani, E.; Shimizu, S. Establishment of Agrobacterium tumefaciens-Mediated Transformation of an Oleaginous Fungus, Mortierella alpina 1S-4, and Its Application for Eicosapentaenoic Acid Producer Breeding. Appl. Environ. Microbiol. 2009, 75, 5529–5535. [Google Scholar] [CrossRef] [PubMed]
- Xin, F.; Wang, R.; Chang, Y.; Gao, M.; Xie, Z.; Yang, W.; Chen, M.; Zhang, H.; Song, Y. Homologous Overexpression of Diacylglycerol Acyltransferase in Oleaginous Fungus Mucor circinelloides WJ11 Enhances Lipid Accumulation under Static Solid Cultivation. J. Agric. Food Chem. 2022, 70, 9073–9083. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Pan, Y.; Ding, W.; Hu, H.; Liu, J. Lipid production is more than doubled by manipulating a diacylglycerol acyltransferase in algae. GCB Bioenergy 2021, 13, 185–200. [Google Scholar] [CrossRef]
- Haslam, R.P.; Hamilton, M.L.; Economou, C.K.; Smith, R.; Hassall, K.L.; Napier, J.A.; Sayanova, O. Overexpression of an endogenous type 2 diacylglycerol acyltransferase in the marine diatom Phaeodactylum tricornutum enhances lipid production and omega-3 long-chain polyunsaturated fatty acid content. Biotechnol. Biofuels 2020, 13, 87. [Google Scholar] [CrossRef]
- Wei, H.; Shi, Y.; Ma, X.; Pan, Y.; Hu, H.; Li, Y.; Luo, M.; Gerken, H.; Liu, J. A type-I diacylglycerol acyltransferase modulates triacylglycerol biosynthesis and fatty acid composition in the oleaginous microalga, Nannochloropsis oceanica. Biotechnol. Biofuels 2017, 10, 174. [Google Scholar] [CrossRef] [PubMed]
- Xin, Y.; Shen, C.; She, Y.; Chen, H.; Wang, C.; Wei, L.; Yoon, K.; Han, D.; Hu, Q.; Xu, J. Biosynthesis of Triacylglycerol Molecules with a Tailored PUFA Profile in Industrial Microalgae. Mol. Plant 2019, 12, 474–488. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Chen, H.; Gu, Z.; Zhang, H.; Chen, W.; Chen, Y.Q. Application of a delta-6 desaturase with alpha-linolenic acid preference on eicosapentaenoic acid production in Mortierella alpina. Microb. Cell Fact. 2016, 15, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Total Biomass (g/L) | Total Lipid Yield (g/L) |
|---|---|---|
| M. alpina ATCC 32222 | 10.35 ± 0.47 | 3.42 ± 0.34 |
| M. alpina-MaDGAT1B | 9.11 ± 0.12 | 3.66 ± 0.1 |
| M. alpina-MaDGAT2A | 9.39 ± 0.26 | 3.69 ± 0.25 |
| Strains | LSO Concentration (g/L) | Total Biomass (g/L) | Total Lipid Yield (g/L) |
|---|---|---|---|
| M. alpina ATCC 32222 | 0 | 10.35 ± 0.47 | 3.42 ± 0.34 |
| 0.5 | 10.82 ± 0.1 | 3.71 ± 0.11 | |
| M. alpina-MaDGAT2A | 0 | 9.39 ± 0.26 | 3.69 ± 0.25 |
| 0.5 | 9.99 ± 0.19 | 4.26 ± 0.38 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, R.; Chang, L.; Cao, J.; Yang, B.; Chen, H.; Chen, W. Applications of Diacylglycerol Acyltransferase for Triacylglycerol Production in Mortierella alpina. J. Fungi 2023, 9, 219. https://doi.org/10.3390/jof9020219
Yu R, Chang L, Cao J, Yang B, Chen H, Chen W. Applications of Diacylglycerol Acyltransferase for Triacylglycerol Production in Mortierella alpina. Journal of Fungi. 2023; 9(2):219. https://doi.org/10.3390/jof9020219
Chicago/Turabian StyleYu, Ruilin, Lulu Chang, Jun Cao, Bo Yang, Haiqin Chen, and Wei Chen. 2023. "Applications of Diacylglycerol Acyltransferase for Triacylglycerol Production in Mortierella alpina" Journal of Fungi 9, no. 2: 219. https://doi.org/10.3390/jof9020219
APA StyleYu, R., Chang, L., Cao, J., Yang, B., Chen, H., & Chen, W. (2023). Applications of Diacylglycerol Acyltransferase for Triacylglycerol Production in Mortierella alpina. Journal of Fungi, 9(2), 219. https://doi.org/10.3390/jof9020219