
The New GPI-Anchored Protein, SwgA, Is Involved in Nitrogen Metabolism in the Pathogenic Filamentous Fungus Aspergillus fumigatus

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Growth Conditions
2.2. Protein Sequence Analysis
2.3. Construction of the ΔswgA Mutant, and the Revertant Strain ΔswgA::swgA
2.4. Manipulation of DNA, RNA, Southern Blotting
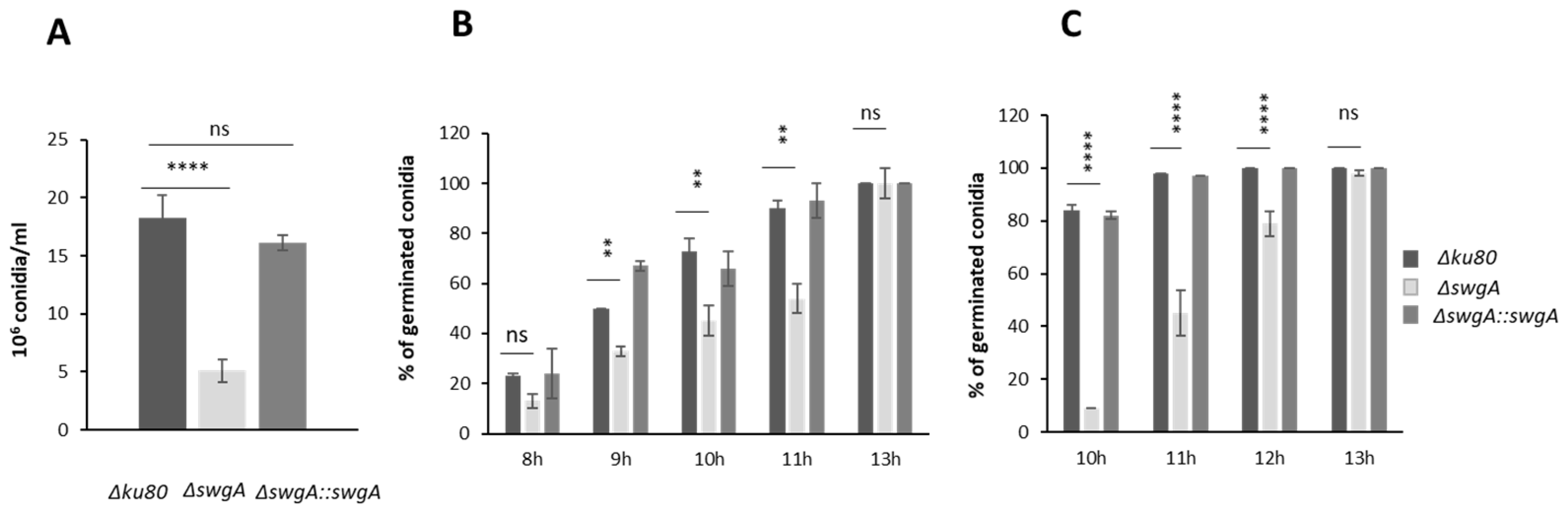
2.5. Conidiation, Conidial Morphology, and Germination
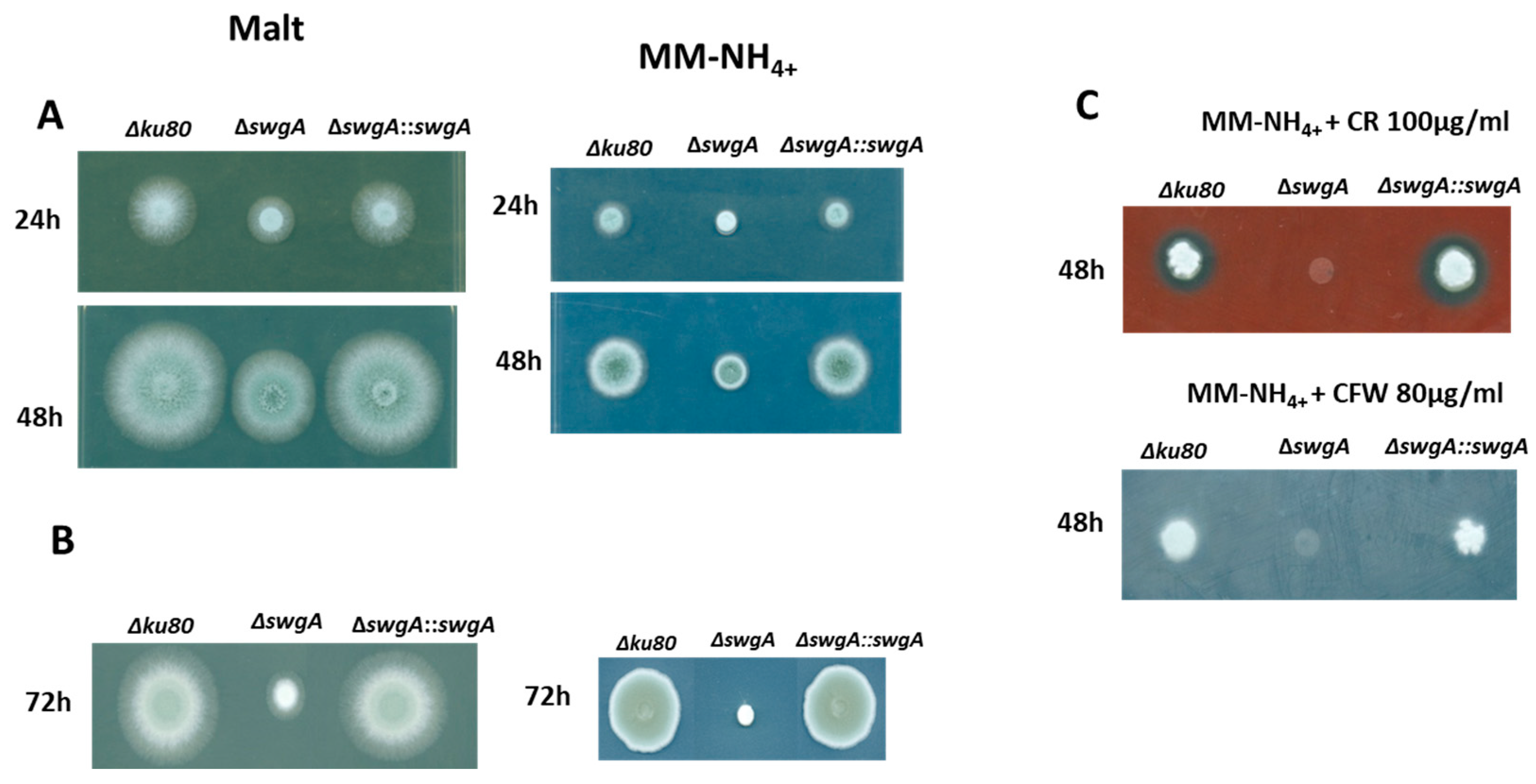
2.6. Growth and Susceptibility of the Mutant Strains to Congo Red and Calcofluor White
2.7. Hydrophobicity Measurements
2.8. Scanning Electron Microscopy (SEM) Analysis
2.9. Adhesion Assays and Biofilm Quantification
2.10. Anti-SwgA Antibodies
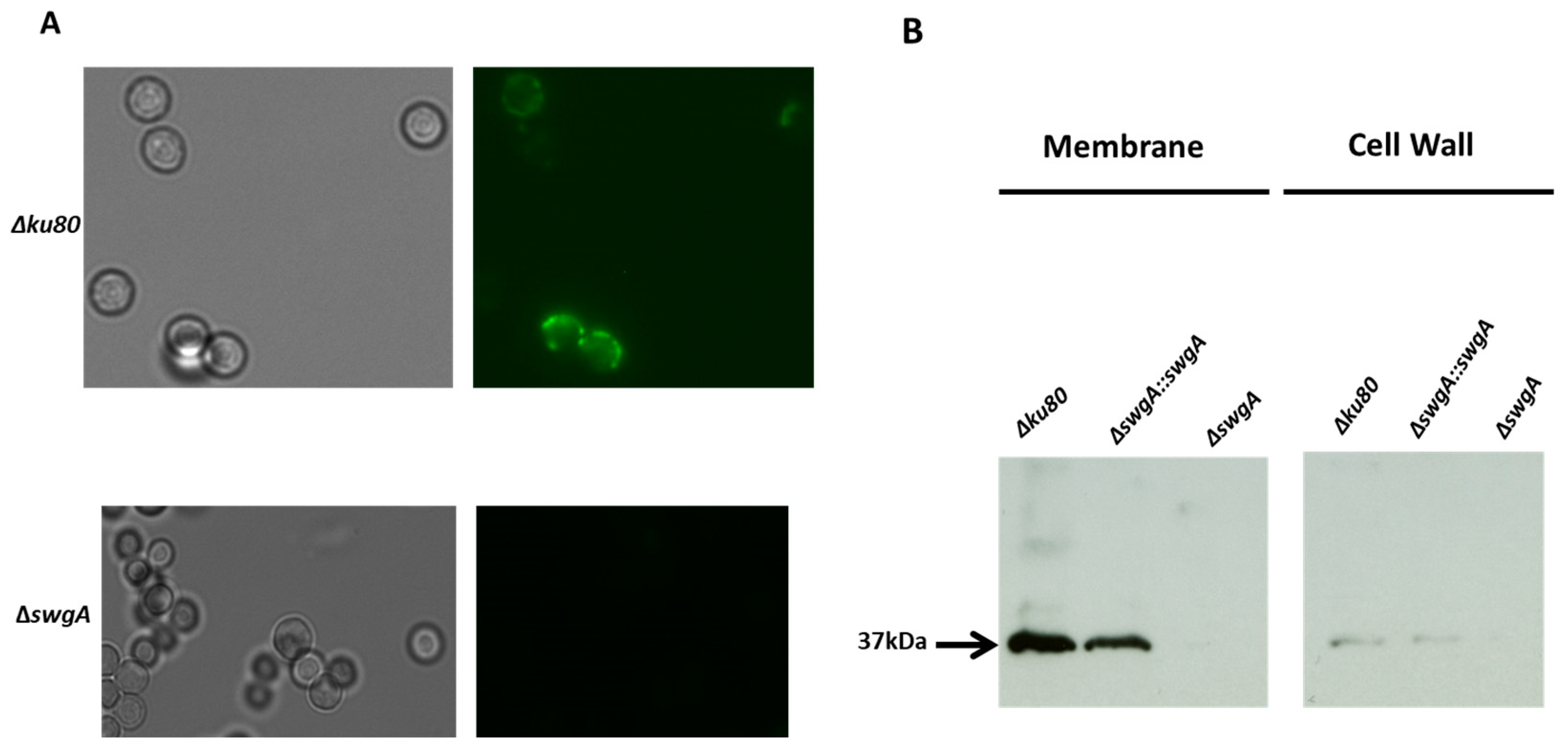
2.11. Immunodetection of SwgA in Cell Wall or Membrane
2.12. Immunofluorescence of Cell Wall Polysaccharides
2.13. Carbohydrate Analysis of the Cell Wall Fractions and Supernatant
2.14. In Vivo Experiments in a Murine Model of Invasive Aspergillosis
2.15. Statistical Analysis
3. Results
3.1. A Unique GPI-Anchored Protein Common to Aspergillus Section Clavati
3.2. SwgA Is Not Involved in Cell Wall Synthesis
3.3. The Growth Phenotype of the ∆swgA Mutant Is Temperature Sensitive
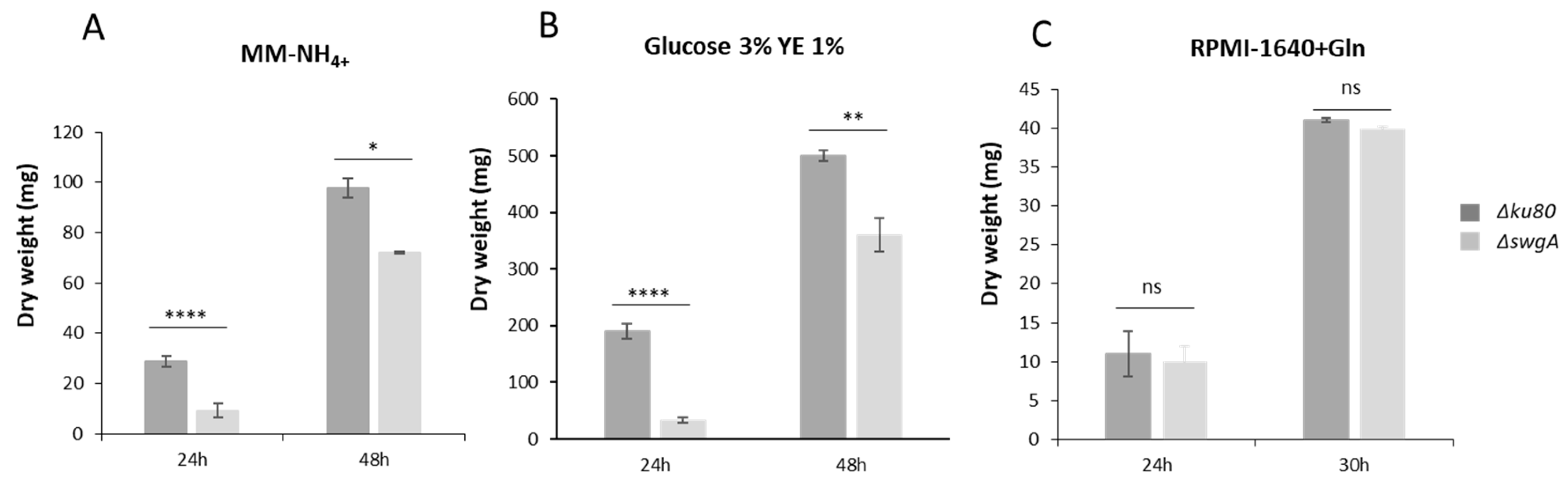
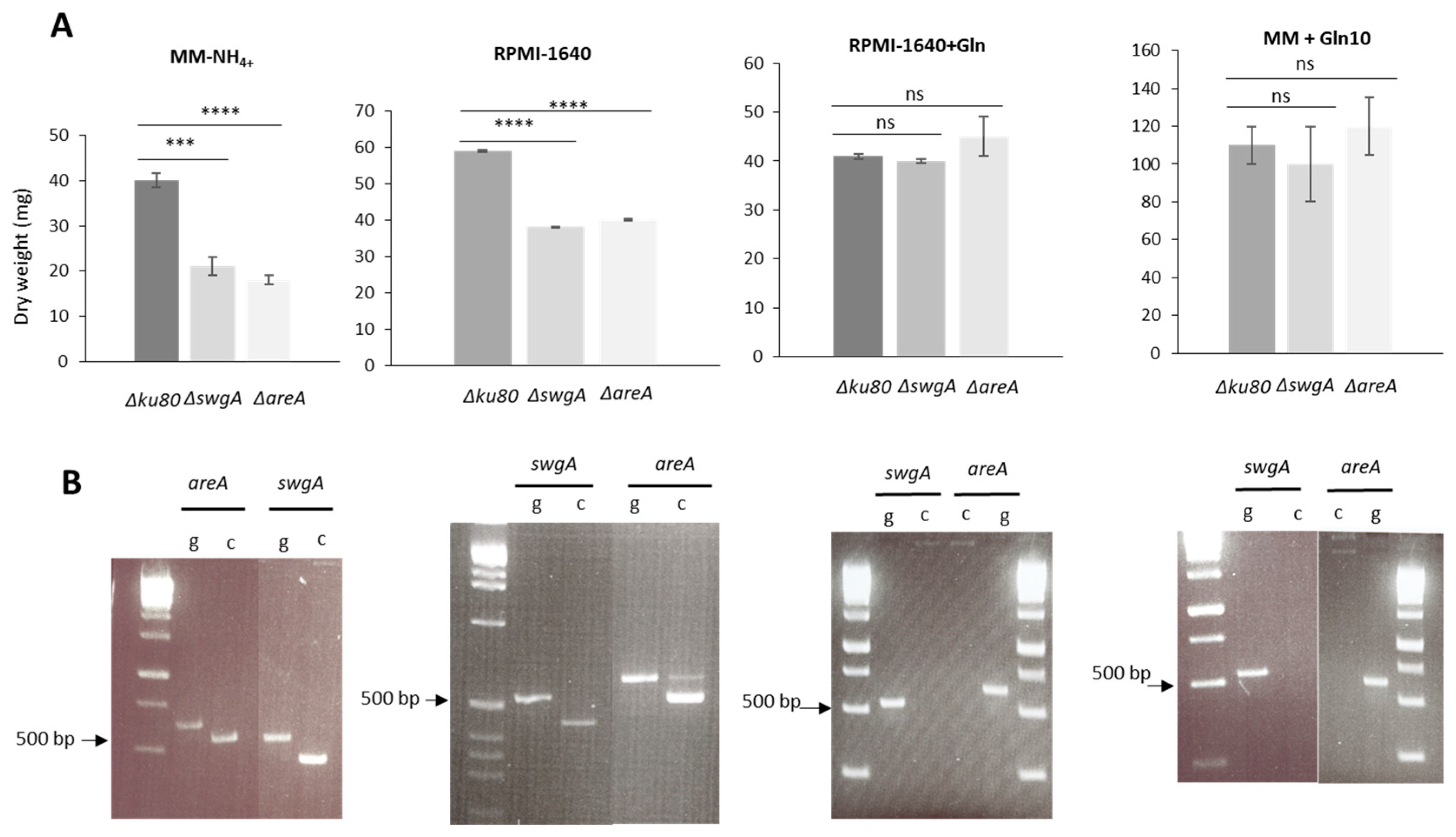
3.4. swgA Is Involved in Nitrogen Metabolism and Is Regulated by the Transcription Factor AreA (AFUA_6G01970)
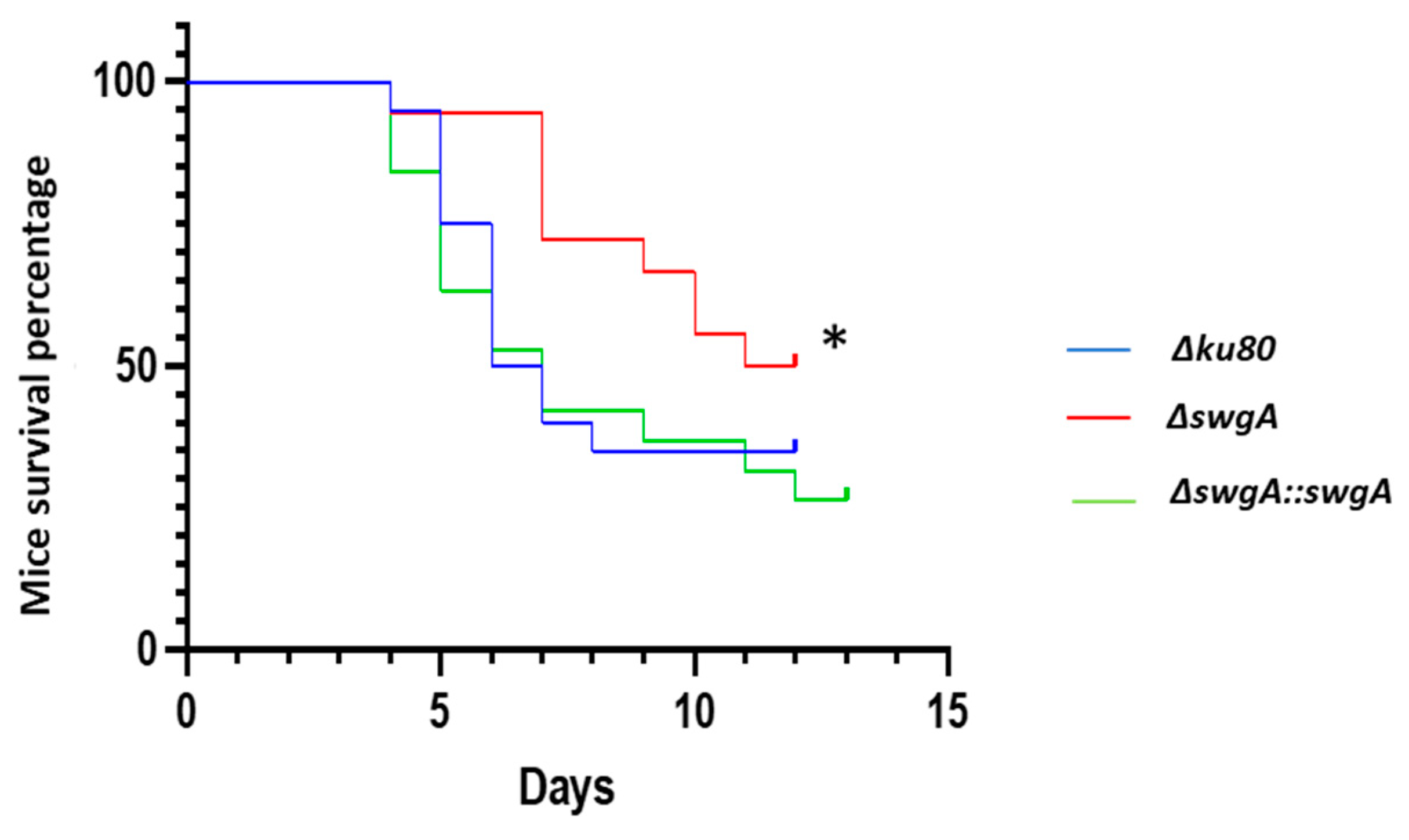
3.5. Virulence of the ΔswgA Mutant Is Affected
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kinoshita, T.; Fujita, M. Biosynthesis of GPI-Anchored Proteins: Special Emphasis on GPI Lipid Remodeling. J. Lipid Res. 2016, 57, 6–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samalova, M.; Carr, P.; Bromley, M.; Blatzer, M.; Moya-Nilges, M.; Latgé, J.-P.; Mouyna, I. GPI Anchored Proteins in Aspergillus Fumigatus and Cell Wall Morphogenesis. Curr. Top. Microbiol. Immunol. 2020, 425, 167–186. [Google Scholar] [PubMed]
- Gastebois, A.; Clavaud, C.; Aimanianda, V.; Latgé, J.-P. Aspergillus Fumigatus: Cell Wall Polysaccharides, Their Biosynthesis and Organization. Future Microbiol. 2009, 4, 583–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouyna, I.; Fontaine, T.; Vai, M.; Monod, M.; Fonzi, W.A.; Diaquin, M.; Popolo, L.; Hartland, R.P.; Latgé, J.P. Glycosylphosphatidylinositol-Anchored Glucanosyltransferases Play an Active Role in the Biosynthesis of the Fungal Cell Wall. J. Biol. Chem. 2000, 275, 14882–14889. [Google Scholar] [CrossRef] [Green Version]
- Mouyna, I.; Morelle, W.; Vai, M.; Monod, M.; Léchenne, B.; Fontaine, T.; Beauvais, A.; Sarfati, J.; Prévost, M.-C.; Henry, C.; et al. Deletion of GEL2 Encoding for a Beta(1-3)Glucanosyltransferase Affects Morphogenesis and Virulence in Aspergillus Fumigatus. Mol. Microbiol. 2005, 56, 1675–1688. [Google Scholar] [CrossRef]
- Gastebois, A.; Fontaine, T.; Latgé, J.-P.; Mouyna, I. Beta(1-3)Glucanosyltransferase Gel4p Is Essential for Aspergillus Fumigatus. Eukaryot. Cell 2010, 9, 1294–1298. [Google Scholar] [CrossRef] [Green Version]
- Aimanianda, V.; Simenel, C.; Garnaud, C.; Clavaud, C.; Tada, R.; Barbin, L.; Mouyna, I.; Heddergott, C.; Popolo, L.; Ohya, Y.; et al. The Dual Activity Responsible for the Elongation and Branching of β-(1,3)-Glucan in the Fungal Cell Wall. MBio 2017, 8, e00619-17. [Google Scholar] [CrossRef] [Green Version]
- Muszkieta, L.; Fontaine, T.; Beau, R.; Mouyna, I.; Vogt, M.S.; Trow, J.; Cormack, B.P.; Essen, L.-O.; Jouvion, G.; Latgé, J.-P. The Glycosylphosphatidylinositol-Anchored DFG Family Is Essential for the Insertion of Galactomannan into the β-(1,3)-Glucan-Chitin Core of the Cell Wall of Aspergillus Fumigatus. mSphere 2019, 4, e00397-19. [Google Scholar] [CrossRef] [Green Version]
- Mouyna, I.; Aimanianda, V.; Hartl, L.; Prevost, M.-C.; Sismeiro, O.; Dillies, M.-A.; Jagla, B.; Legendre, R.; Coppee, J.-Y.; Latgé, J.-P. GH16 and GH81 Family β-(1,3)-Glucanases in Aspergillus Fumigatus Are Essential for Conidial Cell Wall Morphogenesis. Cell. Microbiol. 2016, 18, 1285–1293. [Google Scholar] [CrossRef] [Green Version]
- Gastebois, A.; Mouyna, I.; Simenel, C.; Clavaud, C.; Coddeville, B.; Delepierre, M.; Latgé, J.-P.; Fontaine, T. Characterization of a New Beta(1-3)-Glucan Branching Activity of Aspergillus Fumigatus. J. Biol. Chem. 2010, 285, 2386–2396. [Google Scholar] [CrossRef]
- Caro, L.H.; Tettelin, H.; Vossen, J.H.; Ram, A.F.; van den Ende, H.; Klis, F.M. In Silicio Identification of Glycosyl-Phosphatidylinositol-Anchored Plasma-Membrane and Cell Wall Proteins of Saccharomyces Cerevisiae. Yeast 1997, 13, 1477–1489. [Google Scholar] [CrossRef]
- Plaine, A.; Walker, L.; Da Costa, G.; Mora-Montes, H.M.; McKinnon, A.; Gow, N.A.R.; Gaillardin, C.; Munro, C.A.; Richard, M.L. Functional Analysis of Candida Albicans GPI-Anchored Proteins: Roles in Cell Wall Integrity and Caspofungin Sensitivity. Fungal Genet. Biol. 2008, 45, 1404–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timmermans, B.; De Las Peñas, A.; Castaño, I.; Van Dijck, P. Adhesins in Candida Glabrata. J. Fungi 2018, 4, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaknin, Y.; Shadkchan, Y.; Levdansky, E.; Morozov, M.; Romano, J.; Osherov, N. The Three Aspergillus Fumigatus CFEM-Domain GPI-Anchored Proteins (CfmA-C) Affect Cell-Wall Stability but Do Not Play a Role in Fungal Virulence. Fungal Genet. Biol. 2014, 63, 55–64. [Google Scholar] [CrossRef]
- Woo, P.C.Y.; Lau, S.K.P.; Lau, C.C.Y.; Tung, E.T.K.; Au-Yeung, R.K.H.; Cai, J.-P.; Chong, K.T.K.; Sze, K.H.; Kao, R.Y.; Hao, Q.; et al. Mp1p Homologues as Virulence Factors in Aspergillus Fumigatus. Med. Mycol. 2018, 56, 350–360. [Google Scholar] [CrossRef] [Green Version]
- Levdansky, E.; Kashi, O.; Sharon, H.; Shadkchan, Y.; Osherov, N. The Aspergillus Fumigatus CspA Gene Encoding a Repeat-Rich Cell Wall Protein Is Important for Normal Conidial Cell Wall Architecture and Interaction with Host Cells. Eukaryot. Cell 2010, 9, 1403–1415. [Google Scholar] [CrossRef] [Green Version]
- Valsecchi, I.; Sarikaya-Bayram, Ö.; Wong Sak Hoi, J.; Muszkieta, L.; Gibbons, J.; Prevost, M.-C.; Mallet, A.; Krijnse-Locker, J.; Ibrahim-Granet, O.; Mouyna, I.; et al. MybA, a Transcription Factor Involved in Conidiation and Conidial Viability of the Human Pathogen Aspergillus Fumigatus. Mol. Microbiol. 2017, 105, 880–900. [Google Scholar] [CrossRef] [Green Version]
- Fraczek, M.G.; Bromley, M.; Buied, A.; Moore, C.B.; Rajendran, R.; Rautemaa, R.; Ramage, G.; Denning, D.W.; Bowyer, P. The Cdr1B Efflux Transporter Is Associated with Non-Cyp51a-Mediated Itraconazole Resistance in Aspergillus Fumigatus. J. Antimicrob. Chemother. 2013, 68, 1486–1496. [Google Scholar] [CrossRef]
- Zhao, C.; Fraczek, M.G.; Dineen, L.; Lebedinec, R.; Macheleidt, J.; Heinekamp, T.; Delneri, D.; Bowyer, P.; Brakhage, A.A.; Bromley, M. High-Throughput Gene Replacement in Aspergillus Fumigatus. Curr. Protoc. Microbiol. 2019, 54, e88. [Google Scholar] [CrossRef] [Green Version]
- Valsecchi, I.; Dupres, V.; Stephen-Victor, E.; Guijarro, J.I.; Gibbons, J.; Beau, R.; Bayry, J.; Coppee, J.-Y.; Lafont, F.; Latgé, J.-P.; et al. Role of Hydrophobins in Aspergillus Fumigatus. J. Fungi 2017, 4, 2. [Google Scholar] [CrossRef]
- Sánchez, O.; Aguirre, J. Efficient Transformation of Aspergillus nidulans by Electroporation of Germinated Conidia. Fungal Genet. Rep. 1996, 43, 48–51. [Google Scholar] [CrossRef] [Green Version]
- Girardin, H.; Latgé, J.P.; Srikantha, T.; Morrow, B.; Soll, D.R. Development of DNA Probes for Fingerprinting Aspergillus Fumigatus. J. Clin. Microbiol. 1993, 31, 1547–1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beauvais, A.; Schmidt, C.; Guadagnini, S.; Roux, P.; Perret, E.; Henry, C.; Paris, S.; Mallet, A.; Prévost, M.-C.; Latgé, J.P. An Extracellular Matrix Glues Together the Aerial-Grown Hyphae of Aspergillus Fumigatus. Cell. Microbiol. 2007, 9, 1588–1600. [Google Scholar] [CrossRef]
- Beauvais, A.; Bozza, S.; Kniemeyer, O.; Formosa, C.; Formosa, C.; Balloy, V.; Henry, C.; Roberson, R.W.; Dague, E.; Chignard, M.; et al. Deletion of the α-(1,3)-Glucan Synthase Genes Induces a Restructuring of the Conidial Cell Wall Responsible for the Avirulence of Aspergillus Fumigatus. PLoS Pathog. 2013, 9, e1003716. [Google Scholar] [CrossRef]
- Millet, N.; Latgé, J.-P.; Mouyna, I. Members of Glycosyl-Hydrolase Family 17 of A. fumigatus Differentially Affect Morphogenesis. J. Fungi 2018, 4, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubois, M.; Gilles, K.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Stalhberger, T.; Simenel, C.; Clavaud, C.; Eijsink, V.G.H.; Jourdain, R.; Delepierre, M.; Latgé, J.-P.; Breton, L.; Fontaine, T. Chemical Organization of the Cell Wall Polysaccharide Core of Malassezia Restricta. J. Biol. Chem. 2014, 289, 12647–12656. [Google Scholar] [CrossRef] [Green Version]
- Littlejohn, K.A.; Hooley, P.; Cox, P. Bioinformatics Predicts Diverse Aspergillus Hydrophobins with Novel Properties. Food Hydrocoll. 2012, 27, 503–516. [Google Scholar] [CrossRef]
- Chaudhuri, R.; Ansari, F.A.; Raghunandanan, M.V.; Ramachandran, S. FungalRV: Adhesin Prediction and Immunoinformatics Portal for Human Fungal Pathogens. BMC Genom. 2011, 12, 192. [Google Scholar] [CrossRef] [Green Version]
- Henry, C.; Fontaine, T.; Heddergott, C.; Robinet, P.; Aimanianda, V.; Beau, R.; Beauvais, A.; Mouyna, I.; Prevost, M.-C.; Fekkar, A.; et al. Biosynthesis of Cell Wall Mannan in the Conidium and the Mycelium of Aspergillus Fumigatus. Cell. Microbiol. 2016, 18, 1881–1891. [Google Scholar] [CrossRef]
- Liu, Z.; Raj, S.; van Rhijn, N.; Fraczek, M.; Michel, J.-P.; Sismeiro, O.; Legendre, R.; Varet, H.; Fontaine, T.; Bromley, M.; et al. Functional Genomic and Biochemical Analysis Reveals Pleiotropic Effect of Congo Red on Aspergillus Fumigatus. mBio 2021, 12, e00863-21. [Google Scholar] [CrossRef] [PubMed]
- Aimanianda, V.; Bayry, J.; Bozza, S.; Kniemeyer, O.; Perruccio, K.; Elluru, S.R.; Clavaud, C.; Paris, S.; Brakhage, A.A.; Kaveri, S.V.; et al. Surface Hydrophobin Prevents Immune Recognition of Airborne Fungal Spores. Nature 2009, 460, 1117–1121. [Google Scholar] [CrossRef] [PubMed]
- Pérez, A.; Pedrós, B.; Murgui, A.; Casanova, M.; López-Ribot, J.L.; Martínez, J.P. Biofilm Formation by Candida Albicans Mutants for Genes Coding Fungal Proteins Exhibiting the Eight-Cysteine-Containing CFEM Domain. FEMS Yeast Res. 2006, 6, 1074–1084. [Google Scholar] [CrossRef] [Green Version]
- Pérez, A.; Ramage, G.; Blanes, R.; Murgui, A.; Casanova, M.; Martínez, J.P. Some Biological Features of Candida Albicans Mutants for Genes Coding Fungal Proteins Containing the CFEM Domain. FEMS Yeast Res. 2011, 11, 273–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulkarni, R.D.; Kelkar, H.S.; Dean, R.A. An Eight-Cysteine-Containing CFEM Domain Unique to a Group of Fungal Membrane Proteins. Trends Biochem. Sci. 2003, 28, 118–121. [Google Scholar] [CrossRef] [PubMed]
- Al Abdallah, Q.; Choe, S.-I.; Campoli, P.; Baptista, S.; Gravelat, F.N.; Lee, M.J.; Sheppard, D.C. A Conserved C-Terminal Domain of the Aspergillus Fumigatus Developmental Regulator MedA Is Required for Nuclear Localization, Adhesion and Virulence. PLoS ONE 2012, 7, e49959. [Google Scholar] [CrossRef] [Green Version]
- Sheppard, D.C. Molecular Mechanism of Aspergillus Fumigatus Adherence to Host Constituents. Curr. Opin. Microbiol. 2011, 14, 375–379. [Google Scholar] [CrossRef] [Green Version]
- Takahashi-Nakaguchi, A.; Sakai, K.; Takahashi, H.; Hagiwara, D.; Toyotome, T.; Chibana, H.; Watanabe, A.; Yaguchi, T.; Yamaguchi, M.; Kamei, K.; et al. Aspergillus Fumigatus Adhesion Factors in Dormant Conidia Revealed through Comparative Phenotypic and Transcriptomic Analyses. Cell. Microbiol. 2018, 20, e12802. [Google Scholar] [CrossRef] [Green Version]
- Perez-Cuesta, U.; Guruceaga, X.; Cendon-Sanchez, S.; Pelegri-Martinez, E.; Hernando, F.L.; Ramirez-Garcia, A.; Abad-Diaz-de-Cerio, A.; Rementeria, A. Nitrogen, Iron and Zinc Acquisition: Key Nutrients to Aspergillus Fumigatus Virulence. J. Fungi 2021, 7, 518. [Google Scholar] [CrossRef]
- Tudzynski, B. Nitrogen Regulation of Fungal Secondary Metabolism in Fungi. Front. Microbiol. 2014, 5, 656. [Google Scholar] [CrossRef]
- Scazzocchio, C. The Fungal GATA Factors. Curr. Opin. Microbiol. 2000, 3, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Christensen, T.; Hynes, M.J.; Davis, M.A. Role of the Regulatory Gene AreA of Aspergillus Oryzae in Nitrogen Metabolism. Appl. Environ. Microbiol. 1998, 64, 3232–3237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langdon, T.; Sheerins, A.; Ravagnani, A.; Gielkens, M.; Caddick, M.X.; Arst, H.N. Mutational Analysis Reveals Dispensability of the N-Terminal Region of the Aspergillus Transcription Factor Mediating Nitrogen Metabolite Repression. Mol. Microbiol. 1995, 17, 877–888. [Google Scholar] [CrossRef]
- Hensel, M.; Arst, H.N.; Aufauvre-Brown, A.; Holden, D.W. The Role of the Aspergillus Fumigatus AreA Gene in Invasive Pulmonary Aspergillosis. Mol. Gen. Genet. 1998, 258, 553–557. [Google Scholar] [CrossRef] [PubMed]
- Zou, C.; Wang, P.; Liang, S.; Han, S.; Zheng, S.; Lin, Y. Deletion of the GCW13 Gene Derepresses Gap1-Dependent Uptake of Amino Acids in Pichia Pastoris Grown on Methanol as the Sole Carbon Source. Biochem. Biophys. Res. Commun. 2018, 501, 226–231. [Google Scholar] [CrossRef]
- Brega, E.; Zufferey, R.; Mamoun, C.B. Candida Albicans Csy1p Is a Nutrient Sensor Important for Activation of Amino Acid Uptake and Hyphal Morphogenesis. Eukaryot. Cell 2004, 3, 135–143. [Google Scholar] [CrossRef] [Green Version]
- Kraidlova, L.; Van Zeebroeck, G.; Van Dijck, P.; Sychrová, H. The Candida Albicans GAP Gene Family Encodes Permeases Involved in General and Specific Amino Acid Uptake and Sensing. Eukaryot. Cell 2011, 10, 1219–1229. [Google Scholar] [CrossRef] [Green Version]
- Ries, L.N.A.; Beattie, S.; Cramer, R.A.; Goldman, G.H. Overview of Carbon and Nitrogen Catabolite Metabolism in the Virulence of Human Pathogenic Fungi. Mol. Microbiol. 2018, 107, 277–297. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hexose | GlcNAc | GalNAc | |
|---|---|---|---|
| Δku80-AI | 66.85 ± 2.5 | 28.17 ± 1.9 | 4.98 ± 0.7 |
| ΔswgA-AI | 64.95 ± 5 | 30.31 ± 4.6 | 4.74 ± 0.54 |
| Δku80-AS | 42.63 ± 12.6 | 0 | 57.37 ± 12.7 |
| ΔswgA-AS | 27.54 ± 8.2 | 0 | 72.46 ± 8.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samalova, M.; Flamant, P.; Beau, R.; Bromley, M.; Moya-Nilges, M.; Fontaine, T.; Latgé, J.-P.; Mouyna, I. The New GPI-Anchored Protein, SwgA, Is Involved in Nitrogen Metabolism in the Pathogenic Filamentous Fungus Aspergillus fumigatus. J. Fungi 2023, 9, 256. https://doi.org/10.3390/jof9020256
Samalova M, Flamant P, Beau R, Bromley M, Moya-Nilges M, Fontaine T, Latgé J-P, Mouyna I. The New GPI-Anchored Protein, SwgA, Is Involved in Nitrogen Metabolism in the Pathogenic Filamentous Fungus Aspergillus fumigatus. Journal of Fungi. 2023; 9(2):256. https://doi.org/10.3390/jof9020256
Chicago/Turabian StyleSamalova, Marketa, Patricia Flamant, Rémi Beau, Mike Bromley, Maryse Moya-Nilges, Thierry Fontaine, Jean-Paul Latgé, and Isabelle Mouyna. 2023. "The New GPI-Anchored Protein, SwgA, Is Involved in Nitrogen Metabolism in the Pathogenic Filamentous Fungus Aspergillus fumigatus" Journal of Fungi 9, no. 2: 256. https://doi.org/10.3390/jof9020256
APA StyleSamalova, M., Flamant, P., Beau, R., Bromley, M., Moya-Nilges, M., Fontaine, T., Latgé, J. -P., & Mouyna, I. (2023). The New GPI-Anchored Protein, SwgA, Is Involved in Nitrogen Metabolism in the Pathogenic Filamentous Fungus Aspergillus fumigatus. Journal of Fungi, 9(2), 256. https://doi.org/10.3390/jof9020256