
Seaweed Extracts to Control Postharvest Phytopathogenic Fungi in Rocha Pear


 ,
,  , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Harvesting of Seaweeds and Extraction of Compounds
2.2. In Vitro Assays
2.2.1. Mycelial Growth Inhibition Activity
2.2.2. Spore Germination Inhibition Activity
2.3. In Vivo Assays
2.3.1. Aqueous Extract Toxicity in Rocha Pear
2.3.2. Aqueous Extract Antifungal Activity
2.4. FTIR-ATR Spectroscopic Analysis
2.5. Statistical Analysis
3. Results
3.1. Seaweed Extract Yields
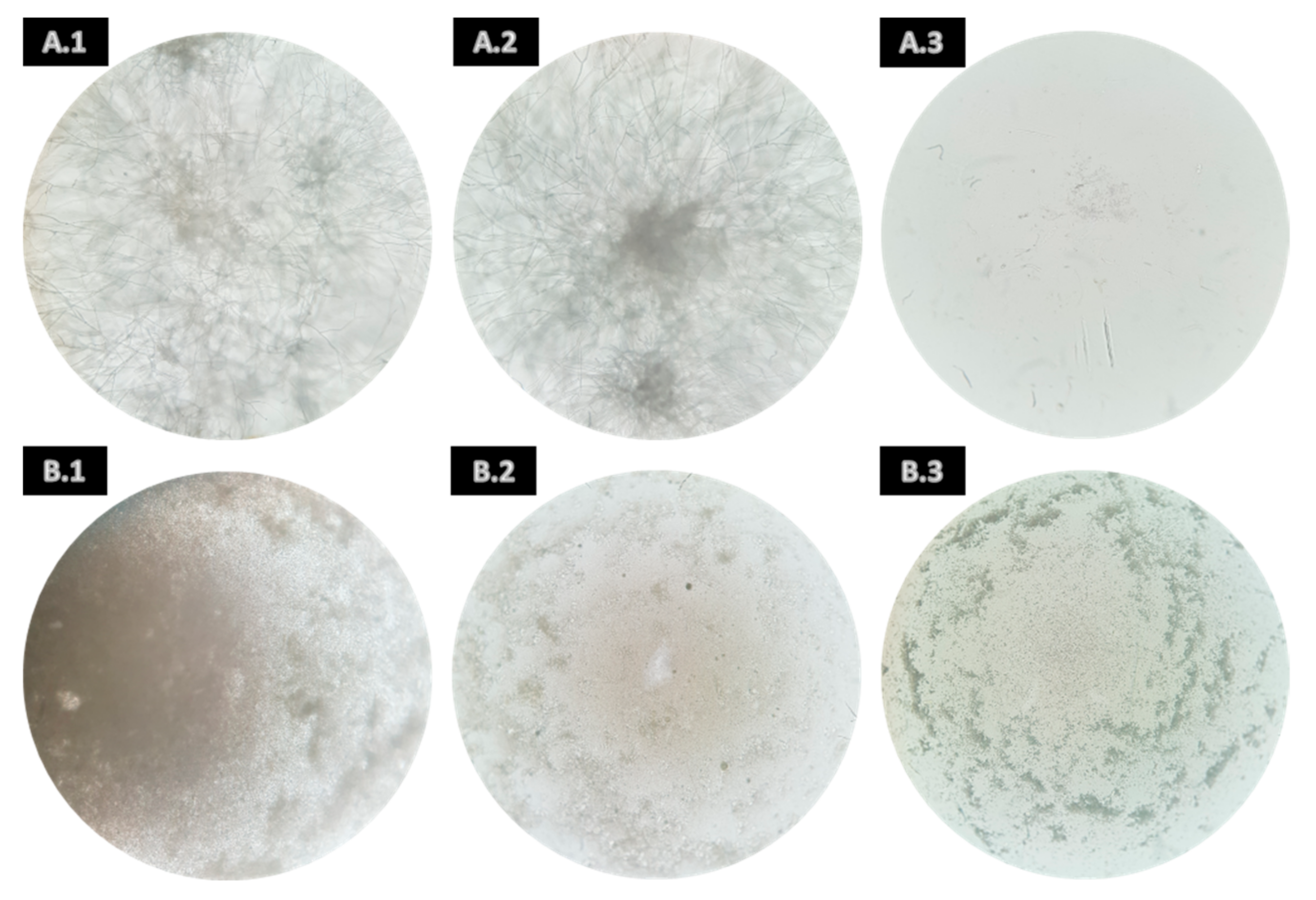
3.2. Mycelial Growth Inhibition Activity
3.3. Spore Germination Inhibition Activity
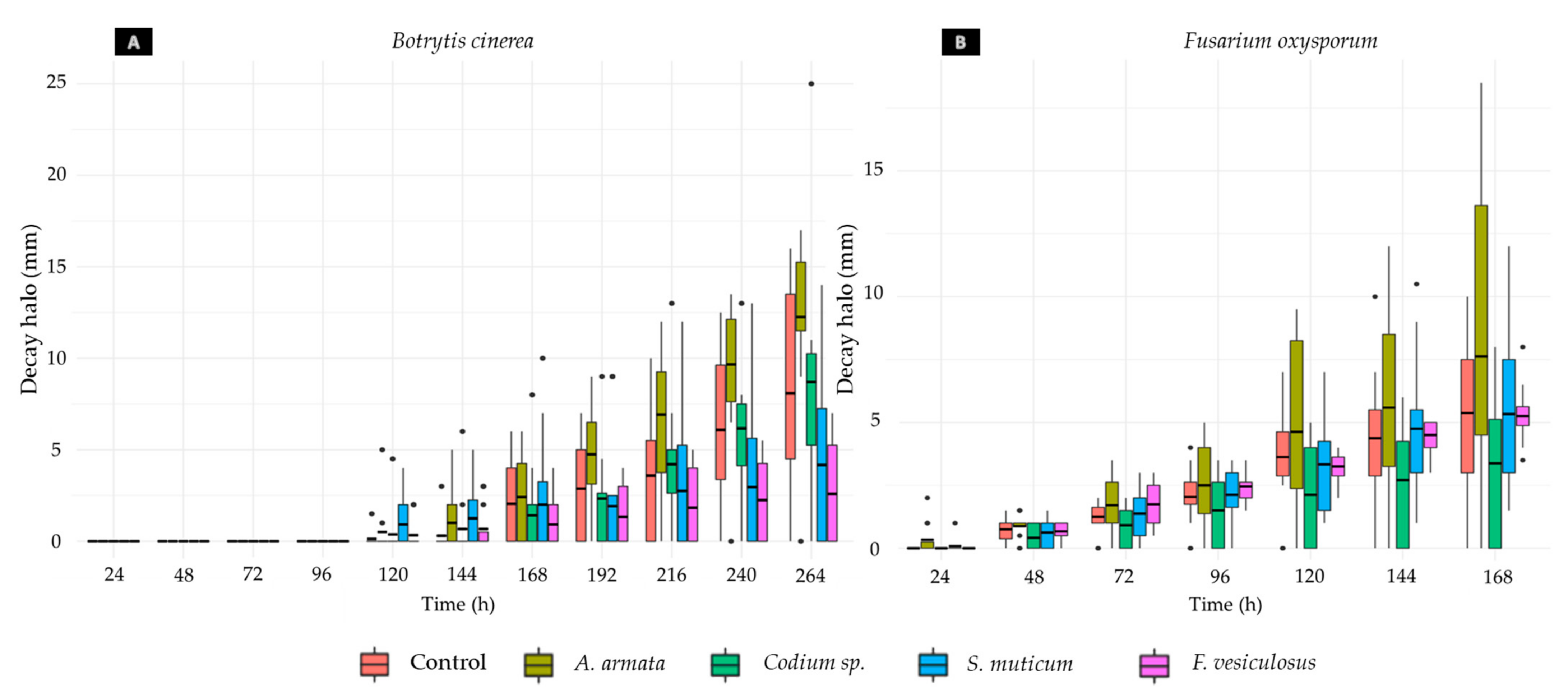
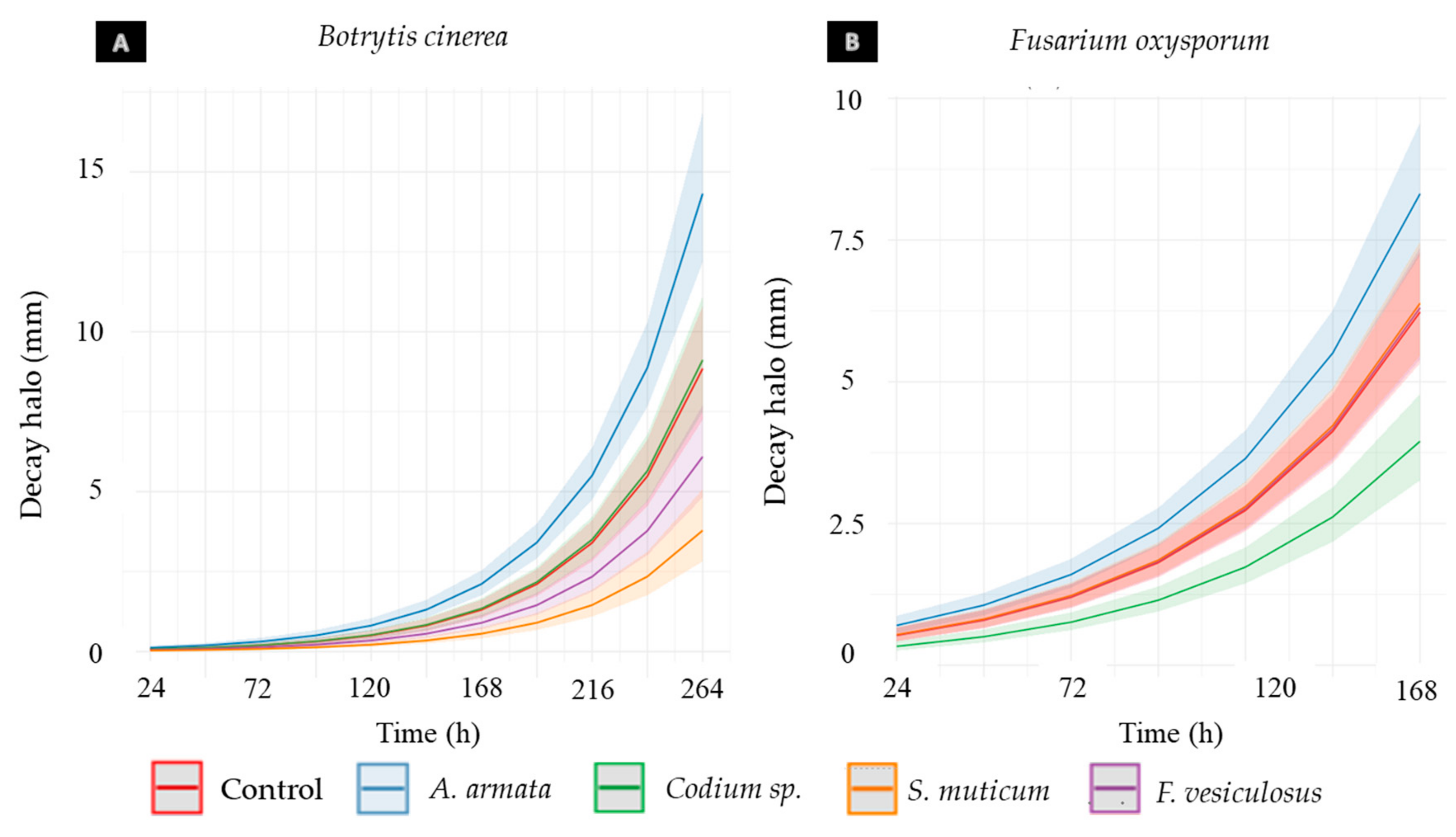
3.4. In Vivo Assays
3.4.1. Aqueous Extract Toxicity in Rocha Pear
3.4.2. Aqueous Extract Antifungal Activity
3.5. Fourier Transform Infrared Spectroscopy (FTIR-ATR)—Analysis
4. Discussion
4.1. In Vitro Assays
4.2. In Vivo Assays
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Transforming Food and Agriculture to Achieve the SDGs. Food and Agriculture Organization of the United Nations (FAO). Available online: http://www.fao.org/sustainability/background/en/ (accessed on 26 December 2022).
- Singh, D.; Sharma, R.R. Postharvest Diseases of Fruits and Vegetables and Their Management. In Postharvest Disinfection of Fruits and Vegetables; Siddiqui, M.W., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 1–52. [Google Scholar]
- Hong, S.Y.; Lansky, E.; Kang, S.S.; Yang, M. A review of pears (Pyrus spp.), ancient functional food for modern times. BMC Complement. Med. Ther. 2021, 21, 219. [Google Scholar] [CrossRef] [PubMed]
- Madbouly, A.K. The Efficacy of Green Synthesized Nanosilver in Reducing the Incidence of Post-Harvest Apple Fruit Brown Rot. J. Fungi 2021, 7, 473. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.Y.; Chen, Y.; Wang, S.Y.; Shi, X.C.; Herrera-Balandrano, D.D.; Polo, V.; Laborda, P. Peel Diffusion and Antifungal Efficacy of Different Fungicides in Pear Fruit: Structure-Diffusion-Activity Relationships. J. Fungi 2022, 8, 547. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Sharma, D.; Gupta, M. Climate Change Impact on Plant Diseases: Opinion, Trends and Mitigation Strategies. In Microbes for Climate Resilient Agriculture; Kashyap, P.L., Srivastava, A.K., Tiwari, S.P.K.S., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2018; pp. 41–56. [Google Scholar]
- Massimiliano; Casaccia, M.; Ciommi, M.; Ferrara, M.; Marchesano, K. Agriculture, climate change and sustainability: The case of EU-28. Ecol. Indic. 2019, 105, 525–543. [Google Scholar] [CrossRef]
- Sardella, D.; Muscat, A.; Brincat, J.-P.; Gatt, R.; Decelis, S.; Valdramidis, V. A Comprehensive Review of the Pear Fungal Diseases. Int. J. Fruit Sci. 2016, 16, 351–377. [Google Scholar] [CrossRef]
- Wenneker, M.; Thomma, B.P.H.J. Latent postharvest pathogens of pome fruit and their management: From single measures to a systems intervention approach. Eur. J. Plant Pathol. 2020, 156, 663–681. [Google Scholar] [CrossRef] [Green Version]
- Brauer, V.S.; Rezende, C.P.; Pessoni, A.M.; de Paula, R.G.; Rangappa, K.S.; Nayaka, S.C.; Gupta, V.K.; Almeida, F. Antifungal Agents in Agriculture: Friends and Foes of Public Health. Biomolecules 2019, 9, 521. [Google Scholar] [CrossRef] [Green Version]
- Oyom, W.; Li, Y.; Prusky, D.; Zhang, Z.; Bi, Y.; Tahergorabi, R. Recent advances in postharvest technology of Asia pears fungi disease control: A review. Physiol. Mol. Plant Pathol. 2022, 117, 101771. [Google Scholar] [CrossRef]
- Palou, L. Postharvest Treatments with Gras Salts to Control Fresh Fruit Decay. Horticulturae 2018, 4, 46. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Shukla, A.; Attri, K.; Kumar, M.; Kumar, P.; Suttee, A.; Singh, G.; Barnwal, R.P.; Singla, N. Global trends in pesticides: A looming threat and viable alternatives. Ecotoxicol. Environ. Saf. 2020, 201, 110812. [Google Scholar] [CrossRef]
- Hernández, A.; Ruiz-Moyano, S.; Galván, A.I.; Merchán, A.V.; Pérez Nevado, F.; Aranda, E.; Serradilla, M.J.; de Guía Córdoba, M.; Martín, A. Anti-Fungal activity of phenolic sweet orange peel extract for controlling fungi responsible for post-harvest fruit decay. Fungal. Biol. 2021, 125, 143–152. [Google Scholar] [CrossRef]
- Brodie, J.; Chan, C.X.; de Clerck, O.; Cock, J.M.; Coelho, S.M.; Gachon, C.; Grossman, A.R.; Mock, T.; Raven, J.A.; Smith, A.G.; et al. The Algal Revolution. Trends Plant Sci. 2017, 22, 726–738. [Google Scholar] [CrossRef] [PubMed]
- Soliman, A.S.; Ahmed, A.Y.; Abdel-ghafour, S.E.; El-sheekh, M.M.; Sobhy, H.M. Antifungal bio-efficacy of the red algae Gracilaria confervoides extracts against three pathogenic fungi of cucumber plant. Middle East J. Appl. Sci. 2018, 8, 727–735. [Google Scholar]
- Vicente, T.F.L.; Lemos, M.F.L.; Félix, R.; Valentão, P.; Félix, C. Marine Macroalgae, a Source of Natural Inhibitors of Fungal Phytopathogens. J. Fungi 2021, 7, 1006. [Google Scholar] [CrossRef] [PubMed]
- Tyskiewicz, K.; Tyskiewicz, R.; Konkol, M.; Rój, E.; Jaroszuk-Sciseł, J.; Skalicka-Wozniak, K. Antifungal Properties of Fucus Vesiculosus L. Supercritical Fluid Extract against Fusarium culmorum and Fusarium oxysporum. Molecules 2019, 24, 3518. [Google Scholar] [CrossRef] [Green Version]
- Pinteus, S.; Alves, C.; Monteiro, H.; Araújo, E.; Horta, A.; Pedrosa, R. Asparagopsis armata and Sphaerococcus coronopifolius as a Natural Source of Antimicrobial Compounds. World J. Microbiol. Biotechnol. 2015, 31, 445–451. [Google Scholar] [CrossRef]
- AlgaeBASE. Available online: https://www.algaebase.org/ (accessed on 1 November 2022).
- Silva, C.O.; Lemos, M.F.L.; Gaspar, R.; Gonçalves, C.; Neto, J.M. The effects of the invasive seaweed Asparagopsis armata on native rock pool communities: Evidences from experimental exclusion. Ecol. Indic. 2021, 125, 107463. [Google Scholar] [CrossRef]
- Pinteus, S.; Lemos, M.F.L.; Alves, C.; Neugebauer, A.; Silva, J.; Thomas, O.P.; Botana, L.M.; Gaspar, H.; Pedrosa, R. Marine invasive macroalgae: Turning a real threat into a major opportunity—The biotechnological potential of Sargassum muticum and Asparagopsis armata. Algal Res. 2018, 34, 217–234. [Google Scholar] [CrossRef]
- Félix, R.; Dias, P.; Félix, C.; Cerqueira, T.; Andrade, P.B.; Valentão, P.; Lemos, M.F.L. The biotechnological potential of Asparagopsis armata: What is nnown of its chemical composition, bioactivities and current market? Algal Res. 2021, 60, 102534. [Google Scholar] [CrossRef]
- Xu, Y.; Wei, J.; Wei, Y.; Han, P.; Dai, K.; Zou, X.; Jiang, S.; Xu, F.; Wang, H.; Sun, J.; et al. Tea tree oil controls brown rot in peaches by damaging the cell membrane of Monilinia fructicola. Postharvest Biol. Technol. 2021, 175, 111474. [Google Scholar] [CrossRef]
- Aredrup, M.C.; Meletiadis, J.; Mouton, J.W.; Lagrou, K.; Hamal, P.; Guinea, J. Subcommittee on Antifungal Susceptibility Testing (AFST) of the ESCMID European Committee for Antimicrobial Susceptibility Testing (EUCAST). Method for the determination of broth dilution minimum inhibitory concentrations of antifungal agents for conidia forming moulds. EUCAST 2020, 12, 23. [Google Scholar]
- Abbaszadeh, S.; Sharifzadeh, A.; Shokri, H.; Khosravi, A.R.; Abbaszadeh, A. Antifungal efficacy of thymol, carvacrol, eugenol and menthol as alternative agents to control the growth of food-relevant fungi. J. Mycol. Med. 2014, 24, e51–e56. [Google Scholar] [CrossRef] [PubMed]
- Kukić, J.; Popović, V.; Petrović, S.; Mucaji, P.; Ćirić, A.; Stojković, D.; Soković, M. Antioxidant and antimicrobial activity of Cynara cardunculus extracts. Food Chem. 2008, 107, 861–868. [Google Scholar] [CrossRef] [Green Version]
- Castro, T.; Roggia, S.; Wekesa, V.W.; de Andrade Moral, R.; Gb Demétrio, C.; Delalibera, I.; Klingen, I. The effect of synthetic pesticides and sulfur used in conventional and organically grown strawberry and soybean on Neozygites floridana, a natural enemy of spider mites. Pest Manag. Sci. 2016, 72, 1752–1757. [Google Scholar] [CrossRef]
- Monakhova, Y.B.; Tsikin, A.M.; Mushtakova, S.P.; Mecozzi, M. Independent component analysis and multivariate curve resolution to improve spectral interpretation of complex spectroscopic data sets: Application to infrared spectra of marine organic matter aggregates. Microchem. J. 2015, 118, 211–222. [Google Scholar] [CrossRef]
- Agatonovic-Kustrin, S.; Ramenskaya, G.; Kustrin, E.; Ortakand, D.B.; Morton, D.W. A new integrated HPTLC-ATR/FTIR approach in marine algae bioprofiling. J. Pharm. Biomed. Anal. 2020, 189, 113488. [Google Scholar] [CrossRef]
- Hssaini, L.; Razouk, R.; Bouslihim, Y. Rapid Prediction of Fig Phenolic Acids and Flavonoids Using Mid-Infrared Spectroscopy Combined with Partial Least Square Regression. Front. Plant. Sci. 2022, 13, 782159. [Google Scholar] [CrossRef]
- Mecozzi, M.; Pietroletti, M.; Gallo, V.; Conti, M.E. Formation of incubated marine mucilages investigated by FTIR and UV-VIS spectroscopy and supported by two-dimensional correlation analysis. Mar. Chem. 2009, 116, 18–35. [Google Scholar] [CrossRef]
- Nunes, N.; Valente, S.; Ferraz, S.; Barreto, M.C.; Pinheiro De Carvalho, M.A.A. Nutraceutical potential of Asparagopsis taxiformis (Delile) Trevisan extracts and assessment of a downstream purification strategy. Heliyon 2018, 4, 957. [Google Scholar] [CrossRef] [Green Version]
- Kalaichelvi, K.; Dhivya, S.M. Screening of phytoconstituents, UV-VIS spectrum and FTIR analysis of Micrococca mercurialis (L.) Benth. Int. J. Herb. Med. 2017, 5, 40–44. [Google Scholar]
- Ragavendran, P.; Sophia, D.; Arul Raj, C.; Gopalakrishnan, V.K. Functional Group Analysis of various extracts of Aerva lanata (L.,) by FTIR Spectrum. Pharmacologyonline 2011, 1, 358–364. [Google Scholar]
- Ferreira, C.A.M.; Félix, R.; Félix, C.; Januário, A.P.; Alves, N.; Novais, S.C.; Dias, J.R.; Lemos, M.F.L. A Biorefinery Approach to the Biomass of the Seaweed Undaria pinnatifida (Harvey Suringar, 1873): Obtaining Phlorotannins-Enriched Extracts for Wound Healing. Biomolecules 2021, 11, 461. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, S.; Alshehri, M.A.; Aziz, A.T.; Panneerselvam, C.; Al-Aoh, H.A.; Maggi, F.; Sut, S.; Dall’Acqua, S. Insecticidal, antibacterial and dye adsorbent properties of Sargassum muticum decorated nano-silver particles. S. Afr. J. Bot. 2021, 139, 432–441. [Google Scholar] [CrossRef]
- Vanavil, B.; Selvaraj, K.; Aanandhalakshmi, R.; Sri, K.U.; Arumugam, M. Bioactive and thermostable sulphated polysaccharide from Sargassum swartzii with drug delivery applications. Int. J. Biol. Macromol. 2020, 153, 190–200. [Google Scholar] [CrossRef]
- Manoharachary, C.; Kunwar, I.K. Host–Pathogen Interaction, Plant Diseases, Disease Management Strategies, and Future Challenges. In Future Challenges in Crop Protection Against Fungal Pathogens, Fungal Biology; Goyal, A., Manoharachary, C., Eds.; Springer: New York, NY, USA, 2014; pp. 185–229. [Google Scholar]
- Belattmania, Z.; Reani, A.; Barakate, M.; Zrid, R.; el Atouani, S.; Hassouani, M.; Eddaoui, A.; Bentiss, F.; Sabour, B. Antimicrobial, antioxidant and alginate potentials of Dictyopteris polypodioides (Dictyotales, Phaeophyceae) from the Moroccan Atlantic coast. Der. Pharma. Chemica. 2016, 8, 216–226. [Google Scholar]
- Zhang, Q.; Xu, C.; Wei, H.; Fan, W.; Li, T. Two pathogenesis-related proteins interact with leucine-rich repeat proteins to promote Alternaria leaf spot resistance in apple. Hortic. Res. 2021, 8, 219. [Google Scholar] [CrossRef]
- AbuQamar, S.; Moustafa, K.; Tran, L.S.P. Mechanisms and strategies of plant defense against Botrytis cinerea. Crit. Rev. Biotechnol. 2017, 37, 262–274. [Google Scholar] [CrossRef]
- Hua, L.; Yong, C.; Zhanquan, Z.; Boqiang, L.; Guozheng, Q.; Shiping, T. Pathogenic mechanisms and control strategies of Botrytis cinerea causing post-harvest decay in fruits and vegetables. Food Qual. Saf. 2018, 2, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Joshi, R. A review of Fusarium oxysporum on its plant interaction and industrial use. J. Med. Plants Stud. 2018, 6, 112–115. [Google Scholar] [CrossRef]
- Jung, B.; Kim, S.; Lee, J. Microcyle Conidiation in Filamentous Fungi. Mycobiology 2014, 42, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Dijksterhuis, J. Fungal spores: Highly variable and stress-resistant vehicles for distribution and spoilage. Food Microbiol. 2019, 81, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Pourakbar, L.; Moghaddama, S.S.; Enshasy, H.A.; Sayyed, R.Z. Antifungal Activity of the Extract of a Macroalgae, Gracilariopsis persica, against Four Plant Pathogenic Fungi in Vitro. Plants 2021, 10, 1781. [Google Scholar] [CrossRef] [PubMed]
- El Baz, F.K.; El-Baroty, G.S.; Ibrahim, A.E.; Abd El Baky, H.H. Cytotoxicity, Antioxidants and Antimicrobial Activities of Lipids Extracted from Some Marine Algae. J. Aquac. Res. Dev. 2014, 5, 284. [Google Scholar] [CrossRef] [Green Version]
- Negara, B.F.S.P.; Sohn, J.H.; Kim, J.S.; Choi, J.S. Antifungal and Larvicidal Activities of Phlorotannins from Brown Seaweeds. Mar. Drugs 2021, 19, 223. [Google Scholar] [CrossRef]
- Righini, H.; Baraldi, E.; Fernández, Y.G.; Quintana, A.M.; Roberti, R. Different Antifungal Activity of Anabaena sp., Ecklonia sp., and Jania sp. Against Botrytis cinerea. Mar. Drugs 2019, 17, 299. [Google Scholar] [CrossRef] [Green Version]
- Biris-Dorhoi, E.S.; Michiu, D.; Pop, C.R.; Rotar, A.M.; Tofana, M.; Pop, O.L.; Socaci, S.A.; Farcas, A.C. Macroalgae—A Sustainable Source of Chemical Compounds with Biological Activities. Nutrients 2020, 12, 3085. [Google Scholar] [CrossRef]
- Benoit, J.M.; Gilmour, C.C.; Mason, R.P. The Influence of Sulfide on Solid-Phase Mercury Bioavailability for Methylation by Pure Cultures of Desulfobulbus propionicus (1pr3). Environ. Sci. Technol. 2001, 35, 127–132. [Google Scholar] [CrossRef]
- Meletiadis, J.; Stergiopoulou, T.; O’Shaughnessy, E.M.; Peter, J.; Walsh, T.J. Concentration-Dependent Synergy and Antagonism within a Triple Antifungal Drug Combination against Aspergillus Species: Analysis by a New Response Surface Model. Antimicrob. Agents Chemother. 2007, 51, 2053–2064. [Google Scholar] [CrossRef] [Green Version]
- Harish, E.; Osherov, N. Fungal Priming: Prepare or Perish. J. Fungi 2022, 8, 448. [Google Scholar] [CrossRef]
- Machado, L.P.; Matsumoto, S.T.; Jamal, C.M.; da Silva, M.B.; da Cruz Centeno, D.; Neto, P.C.; de Carvalho, L.R.; Yokoya, N.S. Chemical analysis and toxicity of seaweed extracts with inhibitory activity against tropical fruit anthracnose fungi. J. Sci. Food. Agric. 2014, 94, 1739–1744. [Google Scholar] [CrossRef]
- Rocha, O.P.; de Felício, R.; Rodrigues, A.H.B.; Ambrósio, D.L.; Cicarelli, R.M.B.; de Albuquerque, S.; Young, M.C.M.; Yokoya, N.S.; Debonsi, H.M. Chemical Profile and Biological Potential of Non-Polar Fractions from Centroceras clavulatum (C. Agardh) Montagne (Ceramiales, Rhodophyta). Molecules 2011, 16, 7105–7114. [Google Scholar] [CrossRef] [PubMed]
- De Felício, R.; de Albuquerque, S.; Young, M.C.M.; Yokoya, N.S.; Debonsi, H.M. Trypanocidal, leishmanicidal and antifungal potential from marine red alga Bostrychia tenella J. Agardh (Rhodomelaceae, Ceramiales). J. Pharm. Biomed. Anal. 2010, 52, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Spriha, S.E.; Rahman, S.A. A Review on Biological Activities of Sugars and Sugar Derivatives. Dhaka Univ. J. Pharm. Sci. 2022, 20, 381–394. [Google Scholar] [CrossRef]
- Jumina, J.; Mutmainah, M.; Purwono, B.; Kurniawan, Y.S.; Syah, Y.M. Antibacterial and Antifungal Activity of Three Monosaccharide Monomyristate Derivatives. Molecules 2019, 24, 3692. [Google Scholar] [CrossRef] [Green Version]
- Augusto, A.; Miranda, A.; Crespo, D.; Campos, M.J.; Raimundo, D.; Pedrosa, R.; Mitchell, G.; Niranjan, K.; Silva, S.F.J. Preservation of fresh-cut Rocha Pear using Codium tomentosum extract. LWT—Food Sci. Technol. 2022, 155, 112938. [Google Scholar] [CrossRef]
- Petrasch, S.; Silva, C.J.; Mesquida-Pesci, S.D.; Gallegos, K.; van den Abeele, C.; Papin, V.; Fernandez-Acero, F.J.; Knapp, S.J.; Blanco-Ulate, B. Infection Strategies Deployed by Botrytis cinerea, Fusarium acuminatum, and Rhizopus stolonifer as a Function of Tomato Fruit Ripening Stage. Front. Plant Sci. 2019, 10, 223. [Google Scholar] [CrossRef] [Green Version]
- Nikkhah, M.; Hashemi, M.; Habibi Najafi, M.B.; Farhoosh, R. Synergistic effects of some essential oils against fungal spoilage on pear fruit. Int. J. Food Microbiol. 2017, 257, 285–294. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Y.; Bi, Y.; Jiang, Q.; Mao, R.; Liu, Z.; Huang, Y.; Zhang, M.; Prusky, D.B. Induction of defense response against Alternaria rot in Zaosu pear fruit by exogenous L-lysine through regulating ROS metabolism and activating defense-related proteins. Postharvest Biol. Technol. 2021, 179, 111567. [Google Scholar] [CrossRef]
- Lutz, M.C.; Colodner, A.; Tudela, M.A.; Carmona, A.C.; Sosa, M.C. Antifungal effects of low environmental risk compounds on development of pear postharvest diseases orchard and postharvest applications. Sci. Hortic. 2022, 295, 110862. [Google Scholar] [CrossRef]
- Pangallo, S.; Li Destri Nicosia, M.G.; Raphael, G.; Levin, E.; Ballistreri, G.; Cacciola, S.O.; Rapisarda, P.; Droby, S.; Schena, L. Elicitation of resistance responses in grapefruit and lemon fruits treated with a pomegranate peel extract. Plant Pathol. 2017, 66, 663–664. [Google Scholar] [CrossRef]
- Sbaihat, L.; Takeyama, K.; Koga, T.; Takemoto, D.; Kawakita, K. Induced resistance in Solanum Lycopersicum by algal elicitor extracted from Sargassum fusiforme. Sci. World J. 2015, 2015, 870520. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extraction Yields (%) | ||||
|---|---|---|---|---|
| Solvents | A. armata | Codium sp. | F. vesiculosus | S. muticum |
| n-hexane | 0.08 | 0.62 | 4.14 | 0.37 |
| Ethyl acetate | 0.29 | 1.19 | 4.62 | 1.25 |
| EtOH:H2O | 2.00 | 20.15 | 20.64 | 6.66 |
| EtOH | 2.01 | 5.96 | 8.60 | 1.22 |
| H2O | 7.91 | 49.78 | 28.69 | 8.99 |
| Relevant Growth Inhibition Rates | ||||
|---|---|---|---|---|
| B. cinerea–A. armata | ||||
| GIR (mm/h) | Extracts | Concentration(mg/mL) | GIR (mm/h) (CI95) | Inhibition (%) |
| n-hexane | 1 | 0.1206 (0.1014, 0.1398) | 55.30 | |
| Ethyl acetate | 1 | 0.1516 (0.1425, 0.1607) | 69.51 | |
| EtOH | 0.5 | 0.1118 (0.0987, 0.1249) | 51.26 | |
| 1 | 0.1109 (0.0898, 0.1319) | 50.85 | ||
| EtOH:H2O | 1 | 0.1475 (0.1400, 0.1549) | 67.63 | |
| Amphotericin B | 30 µg/mL | 0.1417 (0.1345, 0.1488) | 64.97 | |
| GCR (mm/h) | 0.2181 (0.2040, 0.2321) | |||
| B. cinerea–F. vesiculosus | ||||
| Extracts | Concentration (mg/mL) | GIR (mm/h) (CI95) (inhibition %) | Inhibition (%) | |
| GIR (mm/h) | n-hexane | 1 | 0.0977 (0.0859, 0.1095) | 42.59 |
| H2O | 0.1 | 0.1380 (0.1167, 0.1592) | 60.15 | |
| 0.5 | 0.0993 (0.0728, 0.1258) | 43.29 | ||
| Amphotericin B | 30 µg/mL | 0.1711 (0.1587, 0.1834) | 74.56 | |
| GCR (mm/h) | 0.2294 (0.2201, 0.2387) | |||
| P. expansum–A. armata | ||||
| Extracts | Concentration (mg/mL) | GIR (mm/h) (CI95) (inhibition %) | Inhibition (%) | |
| GIR (mm/h) | Ethyl acetate | 1 | 0.0272 (0.0235, 0.0309) | 44.59 |
| Amphotericin B | 30 µg/mL | 0.0447 (0.0437, 0.0455) | 74.07 | |
| GCR (mm/h) | 0.0610 (0.0602, 0.0618) | |||
| A. armata Extracts with Spore Germination Inhibition Activity | ||||
|---|---|---|---|---|
| B. cinerea | F. oxysporum | |||
| Solvents | MIC (mg/mL) | MFC (mg/mL) | MIC (mg/mL) | MFC (mg/mL) |
| n-hexane | 1 | 1 | 0.5 | 1 |
| Ethyl acetate | 0.5 | 1 | 0.5 | 1 |
| EtOH | 0.5 | 0.5 | 1 | - |
| B. cinerea | F. oxysporum | |||||||
|---|---|---|---|---|---|---|---|---|
| Minimum (mm) | Maximum (mm) | Mean ± SD | Number of P0s | Minimum (mm) | Maximum (mm) | Number of P0s | Mean ± SD | |
| Control | 0 | 16 | 8.083 ± 5.780 | 3 | 0 | 10 | 2 | 5.375 ± 3.196 |
| A. armata | 0 | 17 | 12.25 ± 4.361 | 1 | 0 | 18.5 | 1 | 7.625 ± 5.643 |
| Codium sp. | 0 | 25 | 8.708 ± 5.974 | 1 | 0 | 8 | 4 | 3.375 ± 2.754 |
| F. vesiculosus | 0 | 14 | 4.167 ± 5.417 | 7 | 2 | 12 | 0 | 5.333 ± 3.164 |
| S. muticum | 0 | 7 | 2.583 ± 2.753 | 6 | 3.5 | 8 | 0 | 5.250 ± 1.127 |
| B. cinerea | F. oxysporum | |||||
|---|---|---|---|---|---|---|
| Estimate | Std. Error | Real Effect (%) | Estimate | Std. Error | Real Effect (%) | |
| Control | −3.077 | 0.236 | 0.000 | −1.055 | 0.131 | 0.000 |
| A. armata | 0.481 | 0.121 | 61.799 *** | 0.289 | 0.098 | 33.497 * |
| Codium sp. | 0.030 | 0.134 | 3.012 | −0.456 | 0.118 | −36.603 * |
| F. vesiculosus | −0.373 | 0.149 | −31.145 * | 0.012 | 0.104 | 1.196 |
| S. muticum | −0.849 | 0.174 | −57.213 *** | 0.025 | 0.104 | 2.502 |
| Observations | 658 | 418 | ||||
| R2 Nagelkerke | 0.959 | 0.828 | ||||
| Extract | IR (cm−1) | Putative Signal Assignment | References |
|---|---|---|---|
| n-hex (AA) | 2954 | C–H stretching from CH and CH2 aliphatic groups | [29,30] |
| 2923 | O–H stretching from carboxylic acid C–H stretching from CH2 aliphatic groups | [29,31] | |
| 2853 | C–H stretching from CH and CH2 aliphatic groups | [31,32] | |
| 1710 | C=O stretching from aliphatic ketone/carboxylic acid | [32,33] | |
| 1650 | C=O stretching from primary amide | [29] | |
| 1537 | C–N stretching from secondary amide | [29] | |
| 1080 | C–O stretching from primary alcohol | [30] | |
| 720 | C–Cl and C–Br stretching | [34,35] | |
| AE (AA) | 2954 | C–H stretching from CH and CH2 aliphatic groups | [29,30] |
| 2923 | O–H stretching from carboxylic acid C–H stretching from CH2 aliphatic groups | [29,31] | |
| 2853 | C–H stretching from CH and CH2 aliphatic groups | [31,32] | |
| 1709 | C=O stretching from aliphatic ketone/carboxylic acid | [32,33] | |
| 1652 | C=O stretching from primary amide | [29] | |
| 1079 | C–O stretching from ester | [33] | |
| 720 | C–Cl and C–Br stretching | [34,35] | |
| EtOH (AA) | 2924 | O–H stretching from carboxylic acid | [29] |
| 2854 | C–H stretching from alkane group | [32] | |
| 1732 | C=O stretching from aldehyde/ester | [36] | |
| 1649 | C=O stretching from primary amide | [29] | |
| 1174 | C–O stretching from ester | [37] | |
| 1095 | C–O stretching from carbohydrate | [32] | |
| 1035 | C–O stretching from carbohydrate (glycosidic bond) | [38] | |
| AQ (SM) | 2920 | O–H stretching from carboxylic acid | [29] |
| 1603 | C=O stretching from conjugated carbonyl | [30] | |
| 1411 | S=O stretching from sulfate | [38] | |
| 1228 | S=O asymmetric stretching from sulfate esters | [38] | |
| 1027 | C–O stretching from carbohydrate (glycosidic bond) | [38] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toledo, E.; Félix, C.; Vicente, T.F.L.; Augusto, A.; Félix, R.; Toledo, B.; Silva, J.; Trindade, C.; Raimundo, D.; Lemos, M.F.L. Seaweed Extracts to Control Postharvest Phytopathogenic Fungi in Rocha Pear. J. Fungi 2023, 9, 269. https://doi.org/10.3390/jof9020269
Toledo E, Félix C, Vicente TFL, Augusto A, Félix R, Toledo B, Silva J, Trindade C, Raimundo D, Lemos MFL. Seaweed Extracts to Control Postharvest Phytopathogenic Fungi in Rocha Pear. Journal of Fungi. 2023; 9(2):269. https://doi.org/10.3390/jof9020269
Chicago/Turabian StyleToledo, Eloísa, Carina Félix, Tânia F. L. Vicente, Ana Augusto, Rafael Félix, Bernardo Toledo, Joana Silva, Carina Trindade, Délio Raimundo, and Marco F. L. Lemos. 2023. "Seaweed Extracts to Control Postharvest Phytopathogenic Fungi in Rocha Pear" Journal of Fungi 9, no. 2: 269. https://doi.org/10.3390/jof9020269
APA StyleToledo, E., Félix, C., Vicente, T. F. L., Augusto, A., Félix, R., Toledo, B., Silva, J., Trindade, C., Raimundo, D., & Lemos, M. F. L. (2023). Seaweed Extracts to Control Postharvest Phytopathogenic Fungi in Rocha Pear. Journal of Fungi, 9(2), 269. https://doi.org/10.3390/jof9020269