
Structural Analysis of the A Mating Type Locus and Development of the Mating Type Marker of Agaricus bisporus var. bisporus
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Sequence Analysis of the A Mating Type Locus
2.3. Genomic DNA Extraction
2.4. Identification of Mating Types
2.5. Isolation of Homokaryotic Strains from Basidiospores
3. Results
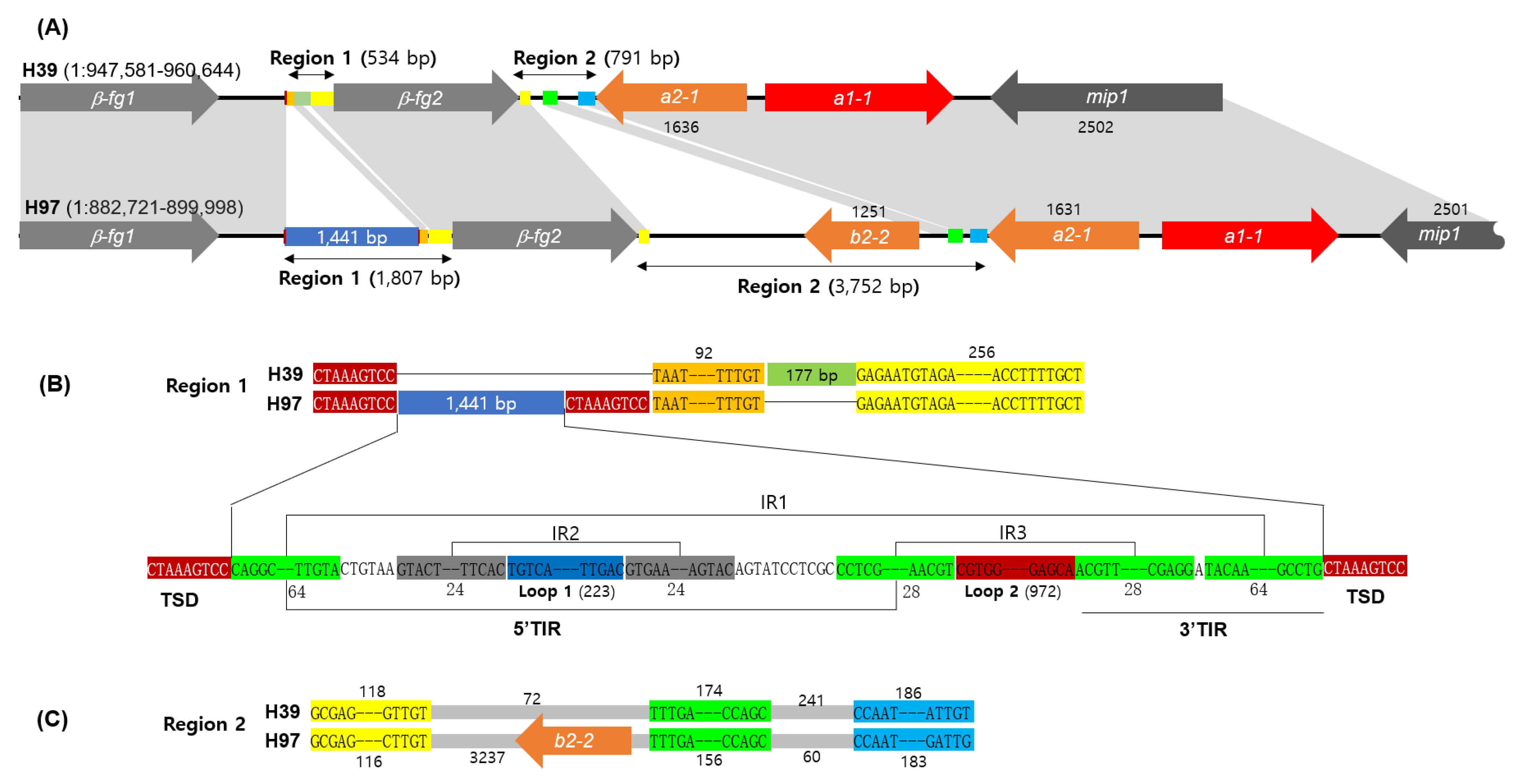
3.1. The A Mating Type Locus Structure of A. bisporus H39
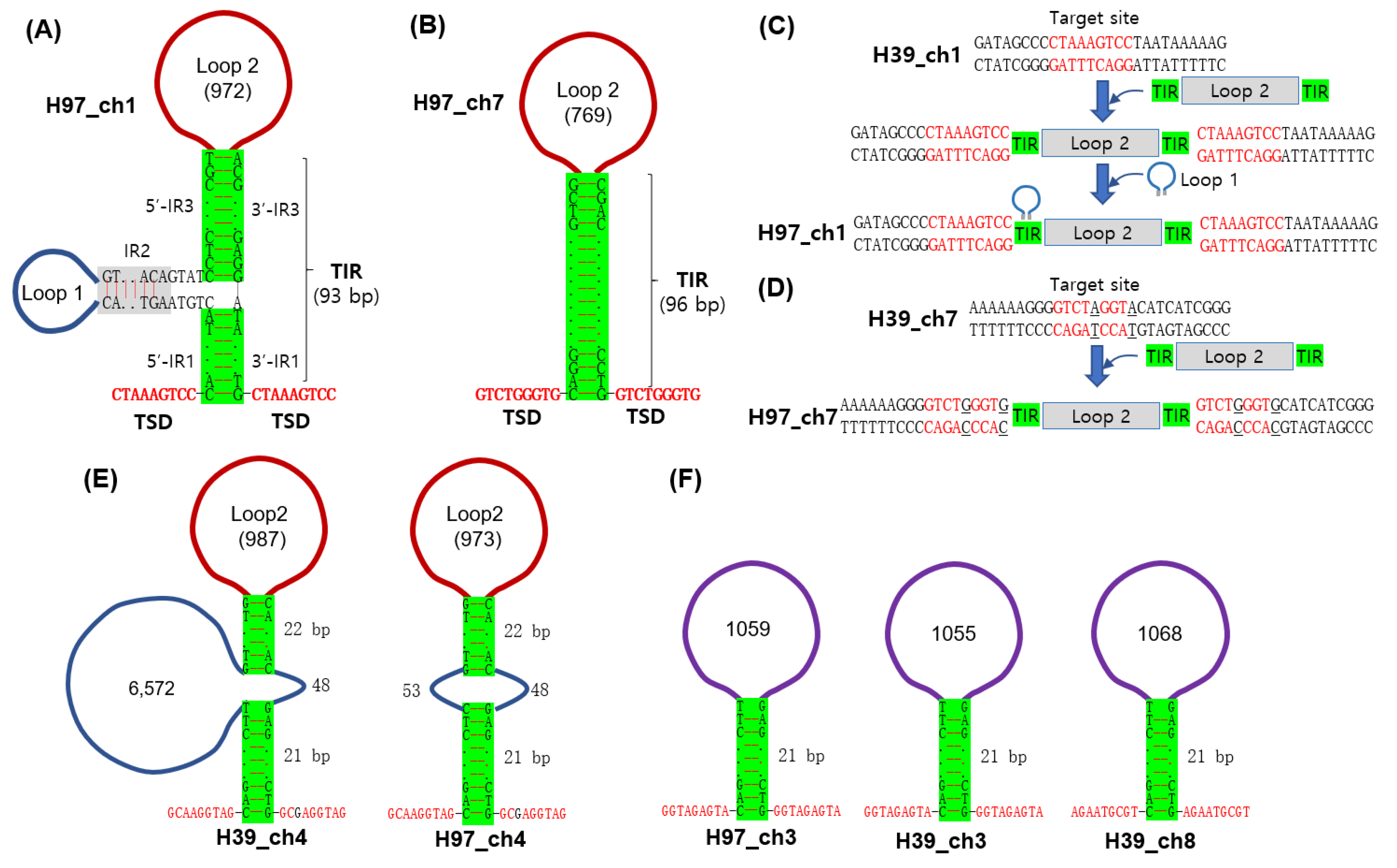
3.2. Analysis of Region 1 in the A Mating Type Locus
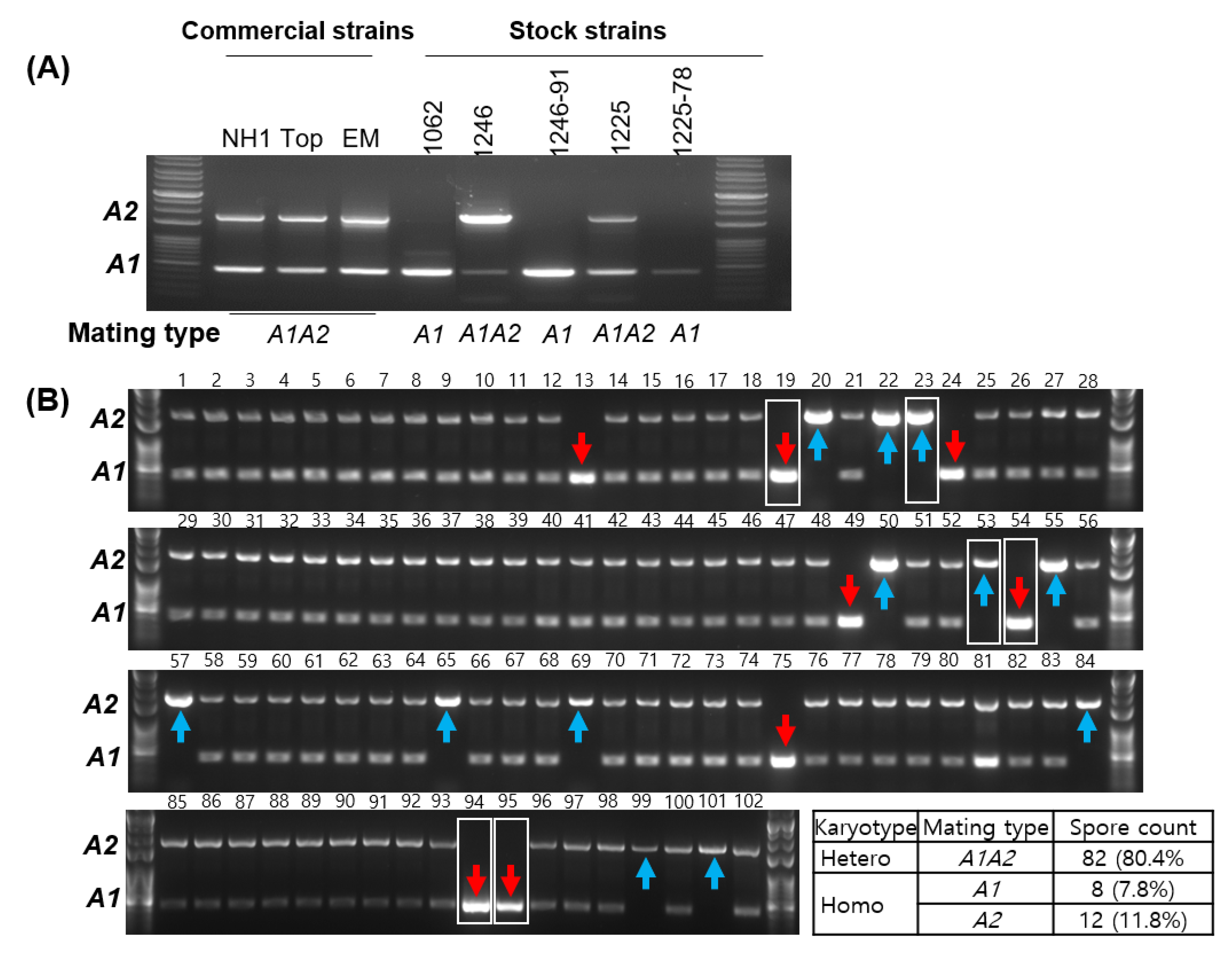
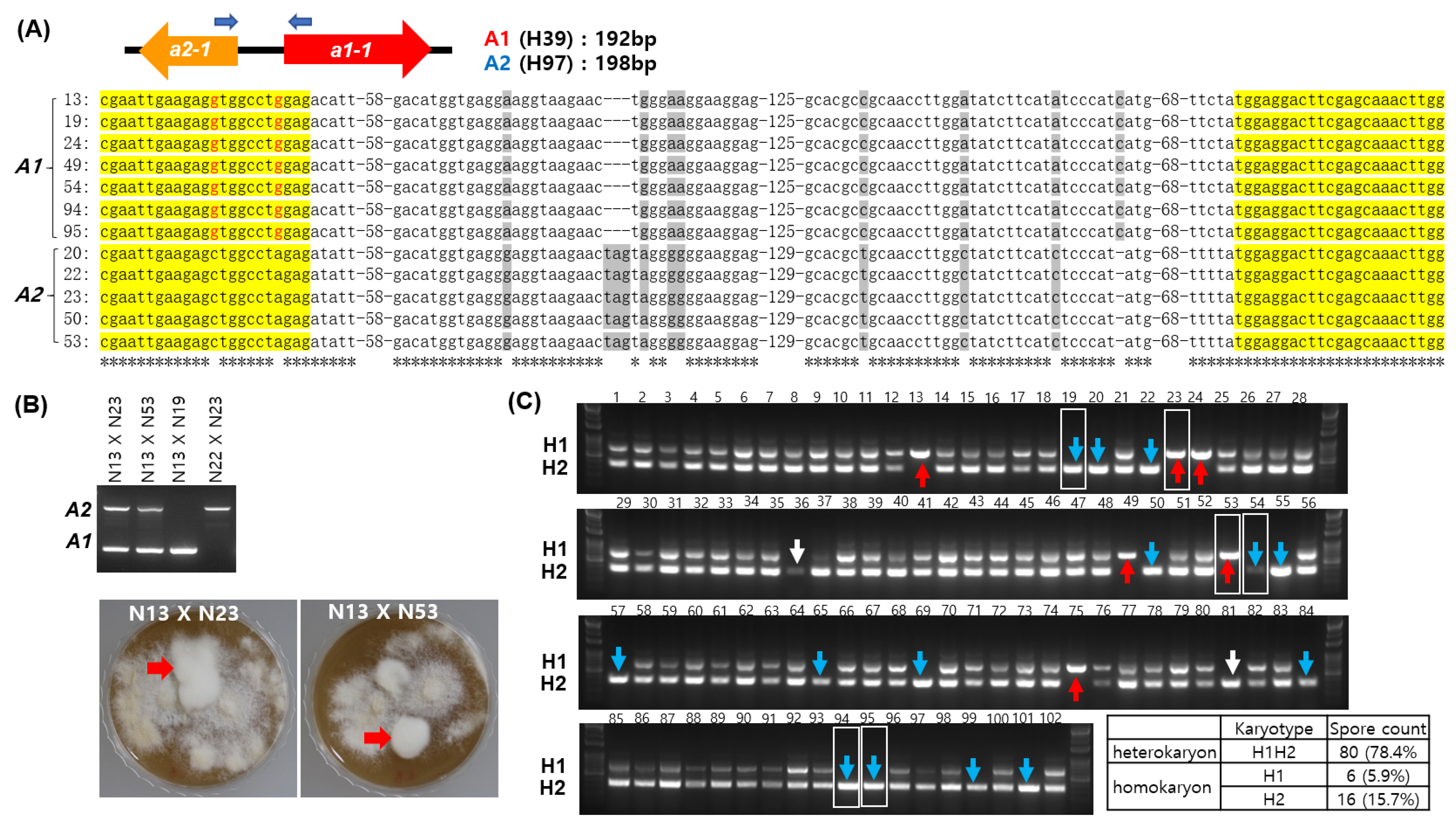
3.3. Simple Verification of the A Mating Type through PCR
3.4. Validation of the A Mating Type Marker and Comparison with the 39Tr 2/5-2/4 Marker
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Appels, F.V.; Dijksterhuis, J.; Lukasiewicz, C.E.; Jansen, K.; Wösten, H.A.; Krijgsheld, P. Hydrophobin gene deletion and environmental growth conditions impact mechanical properties of mycelium by affecting the density of the material. Sci. Rep. 2018, 8, 4703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Stone, M.; Schlagnhaufer, C.; Romaine, C.P. A fruiting body tissue method for efficient Agrobacterium-mediated transformation of Agaricus bisporus. Appl. Environ. Microbiol. 2000, 66, 4510–4513. [Google Scholar] [CrossRef] [Green Version]
- Mikosch, T.S.; Lavrijssen, B.; Sonnenberg, A.S.; van Griensven, L.J. Transformation of the cultivated mushroom Agaricus bisporus (Lange) using T-DNA from Agrobacterium tumefaciens. Curr. Genet. 2001, 39, 35–39. [Google Scholar] [CrossRef]
- Kim, M.S.; Ro, H.S. Generation of iron-independent siderophore-producing Agaricus bisporus through the constitutive expression of hapX. Genes 2021, 12, 724. [Google Scholar] [CrossRef]
- Sugano, S.S.; Suzuki, H.; Shimokita, E.; Chiba, H.; Noji, S.; Osakabe, Y.; Osakabe, K. Genome editing in the mushroom-forming basidiomycete Coprinopsis cinerea, optimized by a high-throughput transformation system. Sci. Rep. 2017, 7, 1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, H.; Xiao, H.; Zou, G.; Zhou, Z.; Zhong, J.J. CRISPR-Cas9 assisted gene disruption in the higher fungus Ganoderma species. Proc. Biochem. 2017, 56, 57–61. [Google Scholar] [CrossRef]
- Moon, S.; An, J.Y.; Choi, Y.J.; Oh, Y.L.; Ro, H.S.; Ryu, H. Construction of a CRISPR/Cas9-mediated genome editing system in Lentinula edodes. Mycobiology 2021, 49, 599–603. [Google Scholar] [CrossRef]
- Yamasaki, F.; Nakazawa, T.; Oh, M.; Bao, D.; Kawauchi, M.; Sakamoto, M.; Honda, Y. Gene targeting of dikaryotic Pleurotus ostreatus nuclei using the CRISPR/Cas9 system. FEMS Microbiol. Lett. 2022, 369, fnac083. [Google Scholar] [CrossRef]
- Waltz, E. Gene-edited CRISPR mushroom escapes US regulation. Nature 2016, 532, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerrigan, R.W.; Royer, J.C.; Baller, L.M.; Kohli, Y.; Horgen, P.A.; Anderson, J. Meiotic behavior and linkage relationships in the secondarily homothallic fungus Agaricus bisporus. Genetics 1993, 133, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Suman, B.C.; Gupta, D. Characterization of single spore isolates of Agaricus bisporus (Lange) imbach using conventional and molecular methods. Curr. Microbiol. 2014, 69, 474–483. [Google Scholar] [CrossRef]
- Summerbell, R.C.; Castle, A.J.; Horgen, P.A.; Anderson, J.B. Inheritance of restriction fragment length polymorphisms in Agaricus brunnescens. Genetics 1989, 123, 293–300. [Google Scholar] [CrossRef]
- Gehrmann, T.; Pelkmans, J.F.; Ohm, R.A.; Vos, A.M.; Sonnenberg, A.S.; Baars, J.J.; Wösten, H.; Reinders, M.; Abeel, T. Nucleus-specific expression in the multinuclear mushroom-forming fungus Agaricus bisporus reveals different nuclear regulatory programs. Proc. Natl. Acad. Sci. USA 2018, 115, 4429–4434. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, K.; Maeda, H.; Sunagawa, M.; Tamai, Y.; Shiraishi, S. Screening of heterozygous DNA markers in shiitake (Lentinula edodes) using de-dikaryotization via preparation of protoplasts and isolation of four meiotic monokaryons from one basidium. J. Wood Sci. 2000, 46, 395–400. [Google Scholar] [CrossRef]
- Nazrul, M.I.; YinBing, B. Differentiation of homokaryons and heterokaryons of Agaricus bisporus with inter-simple sequence repeat markers. Microbiol. Res. 2011, 166, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.N.; Wei, G.A.O.; Wang, Q.Y.; Qu, J.B.; Zhang, J.X.; Huang, C.Y. Identification of commercial cultivars of Agaricus bisporus in China using genome-wide microsatellite markers. J. Integrat. Agricult. 2019, 18, 580–589. [Google Scholar] [CrossRef]
- Pardo, E.H.; O’Shea, S.F.; Casselton, L.A. Multiple versions of the A mating type locus of Coprinus cinereus are generated by three paralogous pairs of multiallelic homeobox genes. Genetics 1996, 144, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Baars, J.J.; Dolstra, O.; Visser, R.G.; Sonnenberg, A.S. Genetic variation and combining ability analysis of bruising sensitivity in Agaricus bisporus. PloS ONE 2013, 8, e76826. [Google Scholar] [CrossRef] [Green Version]
- Foulongne-Oriol, M.; Taskent, O.; Kües, U.; Sonnenberg, A.S.M.; van Peer, A.F.; Giraud, T. Mating-type locus organization and mating-type chromosome differentiation in the bipolar edible button mushroom Agaricus bisporus. Genes 2021, 12, 1079. [Google Scholar] [CrossRef]
- Ha, B.; Kim, S.; Kim, M.; Moon, Y.J.; Song, Y.; Ryu, J.S.; Ryu, H.; Ro, H.S. Diversity of A mating type in Lentinula edodes and mating type preference in the cultivated strains. J. Microbiol. 2018, 56, 416–425. [Google Scholar] [CrossRef]
- Ju, Y.; Kim, S.; Kim, M.; Ryu, J.S.; Ro, H.S. Structure analysis of A and B mating type loci in a representative commercial strain of Pleurotus eryngii. Sci. Hortic. 2020, 274, 109686. [Google Scholar] [CrossRef]
- Kües, U. From two to many: Multiple mating types in Basidiomycetes. Fungal Biol. Rev. 2015, 29, 126–166. [Google Scholar] [CrossRef]
- Sonnenberg, A.S.; de Groot, P.W.; Schaap, P.J.; Baars, J.J.; Visser, J.; Van Griensven, L.J. Isolation of expressed sequence tags of Agaricus bisporus and their assignment to chromosomes. Appl. Environ. Microbiol. 1996, 62, 4542–4547. [Google Scholar] [CrossRef] [Green Version]
- Sonnenberg, A.S.; Gao, W.; Lavrijssen, B.; Hendrickx, P.; Sedaghat-Tellgerd, N.; Foulongne-Oriol, M.; Kong, W.S.; Schijlen, E.; Baars, J.J.; Visser, R.G. A detailed analysis of the recombination landscape of the button mushroom Agaricus bisporus var. bisporus. Fungal Genet. Biol. 2016, 93, 35–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raper, C.A.; Kaye, G. Sexual and other relationships in the genus Agaricus. Microbiology 1978, 105, 135–151. [Google Scholar] [CrossRef] [Green Version]
- Kerrigan, R.W.; Baller, L.M.; Horgen, P.A.; Anderson, J.B. Strategies for the efficient recovery of Agaricus bisporus homokaryons. Mycologia 1992, 84, 575–579. [Google Scholar] [CrossRef]
- Yi, R.; Tachikawa, T.; Ishikawa, M.; Mukaiyama, H.; Bao, D.; Aimi, T. Genomic structure of the A mating-type locus in a bipolar basidiomycete, Pholiota nameko. Mycol. Res. 2009, 113, 240–248. [Google Scholar] [CrossRef]
- James, T.Y.; Srivilai, P.; Kües, U.; Vilgalys, R. Evolution of the bipolar mating system of the mushroom Coprinellus disseminatus from its tetrapolar ancestors involves loss of mating-type-specific pheromone receptor function. Genetics 2006, 172, 1877–1891. [Google Scholar] [CrossRef] [Green Version]
- James, T.Y.; Lee, M.; van Diepen, L.T.A. A single mating-type locus composed of homeodomain genes promotes nuclear migration and heterokaryosis in the white-rot fungus Phanerochaete chrysosporium. Eukaryot. Cell 2011, 10, 249–261. [Google Scholar] [CrossRef] [Green Version]
- Casselton, L.A.; Olesnicky, N.S. Molecular genetics of mating recognition in basidiomycete fungi. Microbiol. Mol. Biol. Rev. 1998, 62, 55–70. [Google Scholar] [CrossRef] [Green Version]
- Gioti, A.; Mushegian, A.A.; Strandberg, R.; Stajich, J.E.; Johannesson, H. Unidirectional evolutionary transitions in fungal mating systems and the role of transposable elements. Mol. Biol. Evol. 2012, 29, 3215–3226. [Google Scholar] [CrossRef] [Green Version]
- Bakkeren, G.; Jiang, G.; Warren, R.L.; Butterfield, Y.; Shin, H.; Chiu, R.; Linning, R.; Schein, J.; Lee, N.; Hu, G.; et al. Mating factor linkage and genome evolution in basidiomycetous pathogens of cereals. Fungal Genet. Biol. 2006, 43, 655–666. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Song, Y.; Ha, B.; Moon, Y.J.; Kim, M.; Ryu, H.; Ro, H.S. Variable number tandem repeats in the mitochondrial DNA of Lentinula edodes. Genes 2019, 10, 542. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, Y.-J.; Jung, S.; Eom, H.; Hoang, T.; Han, H.-G.; Kim, S.; Ro, H.-S. Structural Analysis of the A Mating Type Locus and Development of the Mating Type Marker of Agaricus bisporus var. bisporus. J. Fungi 2023, 9, 284. https://doi.org/10.3390/jof9030284
Choi Y-J, Jung S, Eom H, Hoang T, Han H-G, Kim S, Ro H-S. Structural Analysis of the A Mating Type Locus and Development of the Mating Type Marker of Agaricus bisporus var. bisporus. Journal of Fungi. 2023; 9(3):284. https://doi.org/10.3390/jof9030284
Chicago/Turabian StyleChoi, Yeon-Jae, Sujin Jung, Hyerang Eom, Thimen Hoang, Hui-Gang Han, Sinil Kim, and Hyeon-Su Ro. 2023. "Structural Analysis of the A Mating Type Locus and Development of the Mating Type Marker of Agaricus bisporus var. bisporus" Journal of Fungi 9, no. 3: 284. https://doi.org/10.3390/jof9030284
APA StyleChoi, Y. -J., Jung, S., Eom, H., Hoang, T., Han, H. -G., Kim, S., & Ro, H. -S. (2023). Structural Analysis of the A Mating Type Locus and Development of the Mating Type Marker of Agaricus bisporus var. bisporus. Journal of Fungi, 9(3), 284. https://doi.org/10.3390/jof9030284