


Contribution of the Tyrosinase (MoTyr) to Melanin Synthesis, Conidiogenesis, Appressorium Development, and Pathogenicity in Magnaporthe oryzae

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sequence Alignment Assays
2.2. Fungal Strains and Culture Conditions
2.3. Protoplast Preparation
2.4. Targeted Gene Deletion of MoTyr Mutants
2.5. Assays for Conidial Production, Growth, and Development
2.6. Analysis for Appressorium Integrity
2.7. Rice Sheath Penetration and Plant Infection Assays
2.8. Extraction and Purification of Melanin
2.9. Statistical Analysis
3. Results
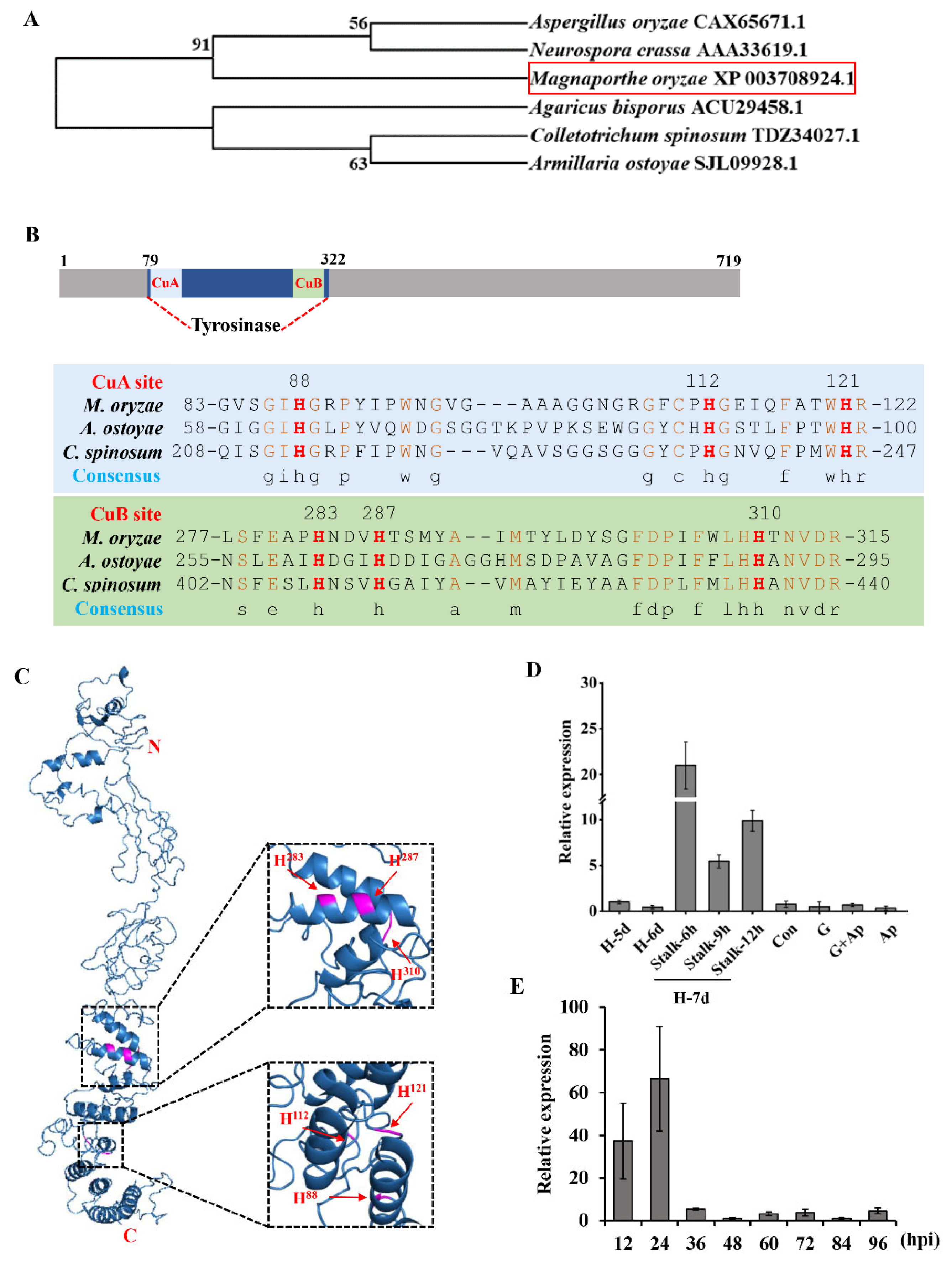
3.1. MGG_14598 Encodes a Tyrosinase in M. oryzae
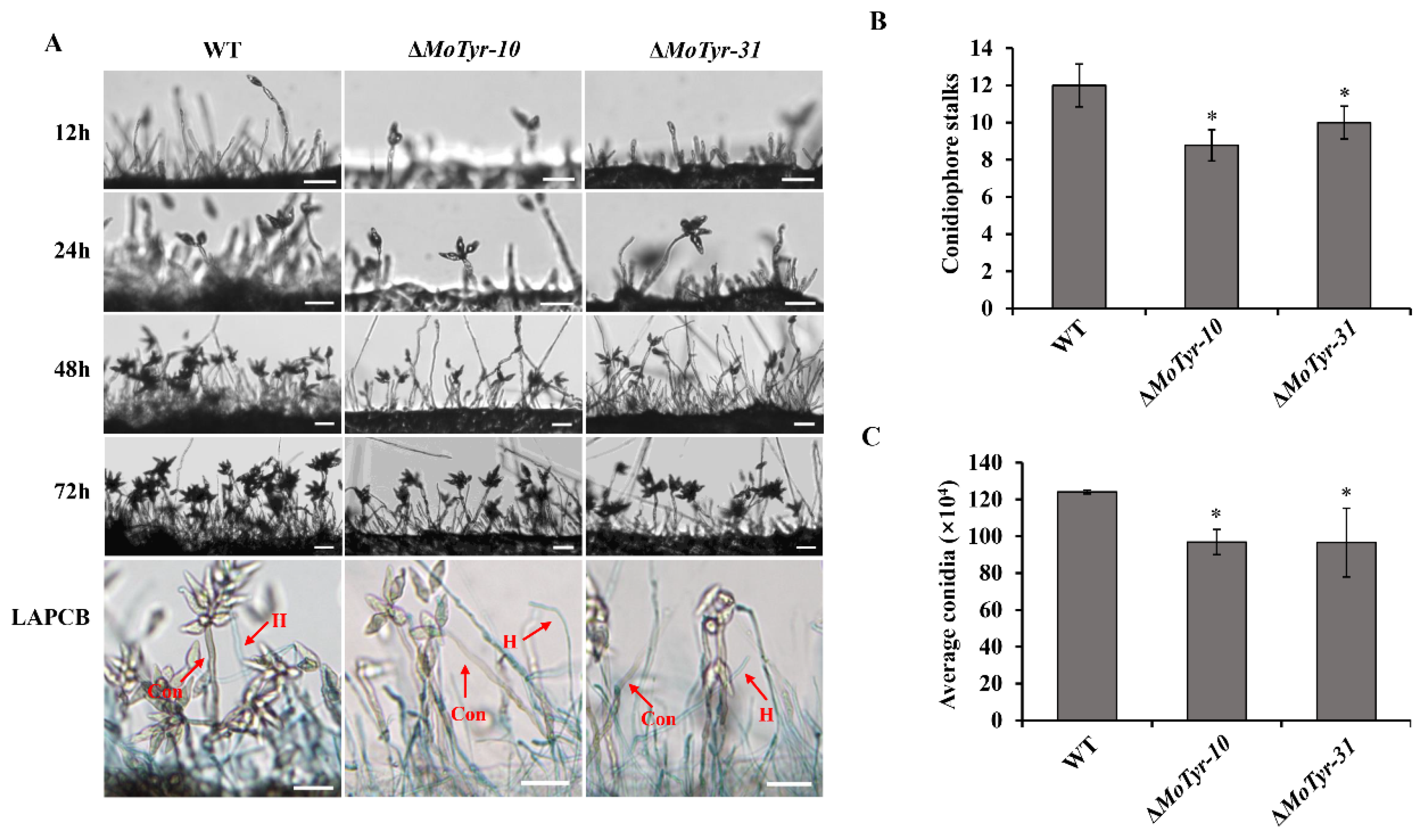
3.2. MoTyr Is Important for Conidiophore Stalks and Conidiation Development
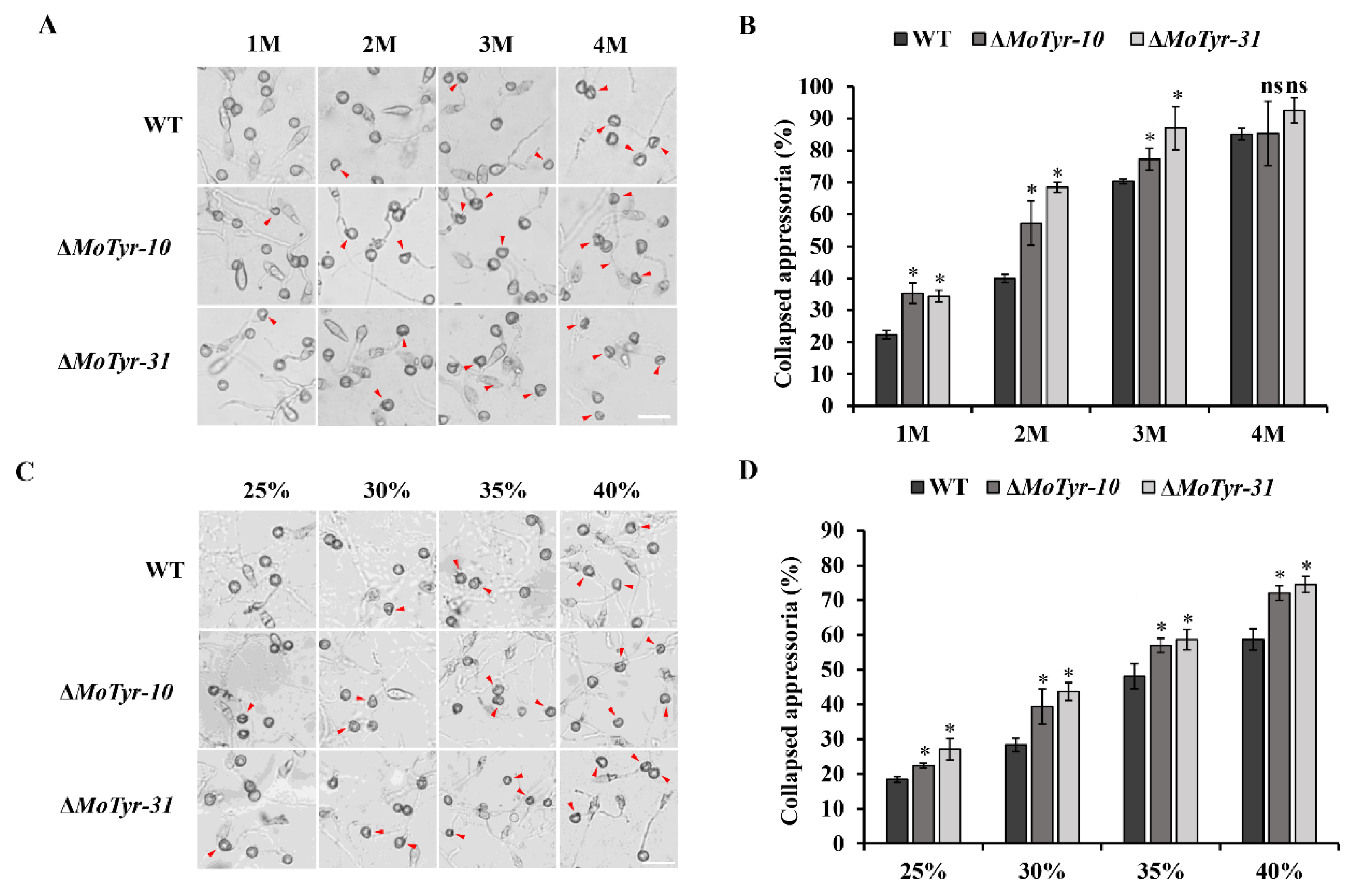
3.3. MoTyr Is Essential for the M. oryzae Appressoria Integrity
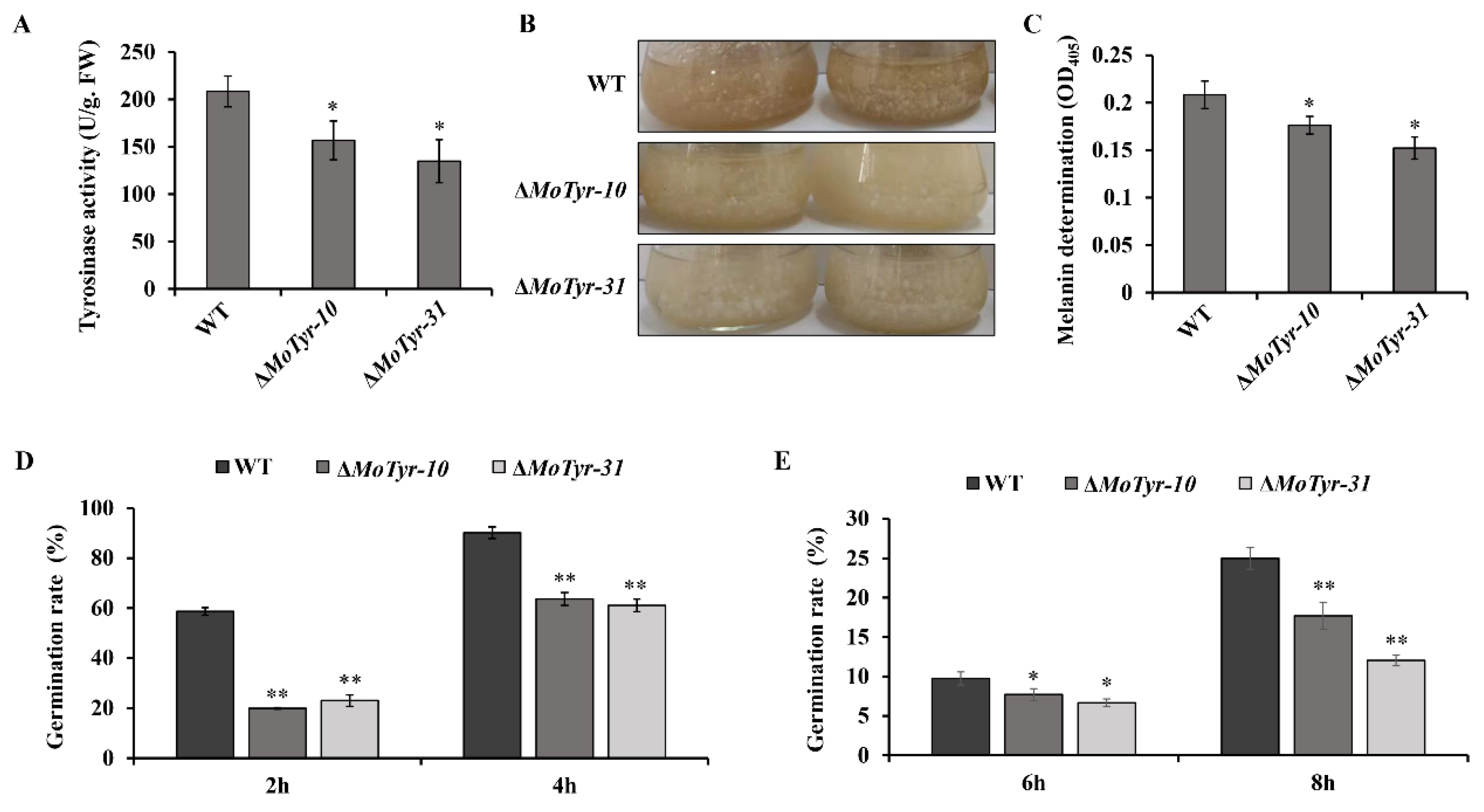
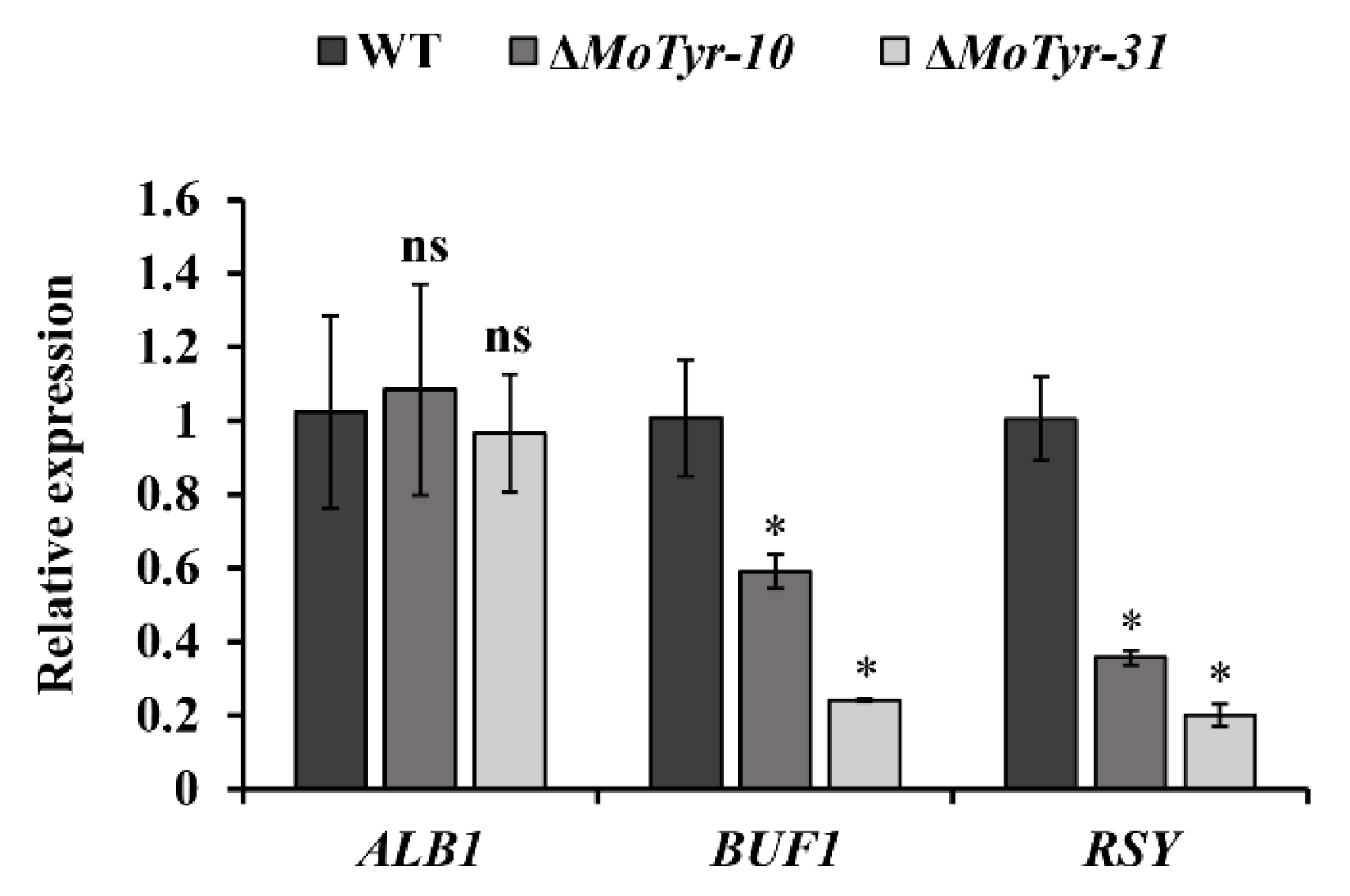
3.4. MoTyr Loss Affected Melanin Synthesis and Stress Resistance in M. oryzae
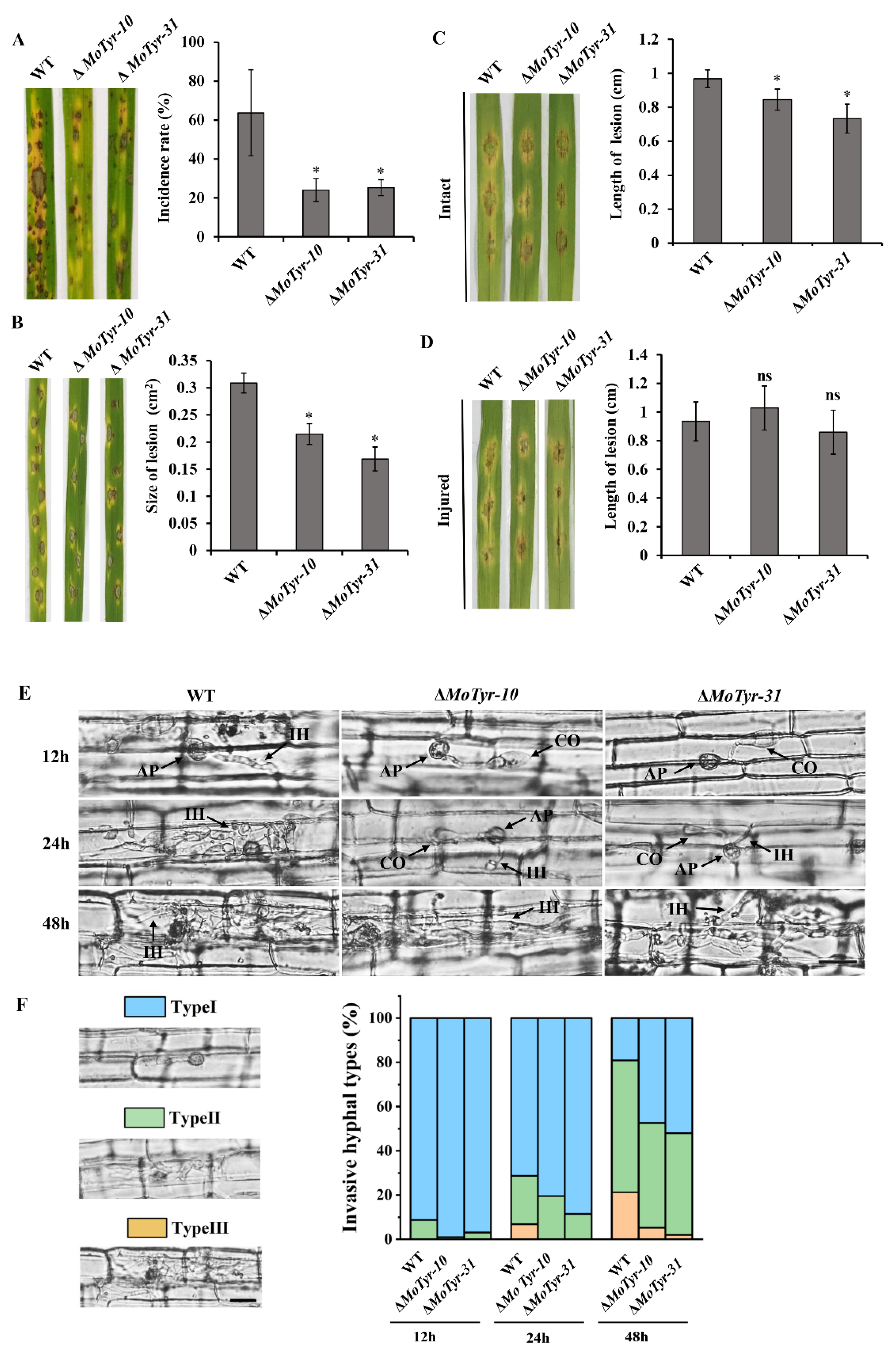
3.5. MoTyr Is Important for Pathogenicity in M. oryzae
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tran-Ly, A.N.; Reyes, C.; Schwarze, F.W.; Ribera, J. Microbial production of melanin and its various applications. World J. Microbiol. Biotechnol. 2020, 36, 170. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Meng, X.; Mo, C.; Wei, X.; Ma, A. Melanin of fungi: From classification to application. World J. Microbiol. Biotechnol. 2022, 38, 228. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.F.Q.; Casadevall, A. The role of melanin in fungal pathogenesis for animal hosts. In Fungal Physiology and Immunopathogenesis; Springer: Cham, Switzerland, 2019; pp. 1–30. [Google Scholar]
- Emidio, C.P.E.E.; Urán, J.M.; Silva, B.R.L.S.; Dias, L.; Doprado, M.; Nosanchuk, J.D.; Taborda, C.P. Melanin as a virulence factor in different species of genus Paracoccidioides. J. Fungi 2020, 6, 291. [Google Scholar] [CrossRef]
- Pal, A.K.; Gajjar, D.U.; Vasavada, A.R. DOPA and DHN pathway orchestrate melanin synthesis in Aspergillus species. Med. Mycol. 2014, 52, 10–18. [Google Scholar]
- Butler, M.J.; Day, A.W. Fungal melanins: A review. Can. J. Microbiol. 1998, 44, 1115–1136. [Google Scholar] [CrossRef]
- Strycker, B.D.; Han, Z.; Bahari, A.; Pham, T.; Lin, X.; Shaw, B.D.; Sokolov, A.V.; Scully, M.O. Raman characterization of fungal DHN and DOPA melanin biosynthesis pathways. J. Fungi 2021, 7, 841. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.A.; Wheeler, M.H. Biosynthesis and functions of fungal melanins. Ann. Rev. Phytopathol. 1986, 24, 411–451. [Google Scholar] [CrossRef]
- Woloshuk, C.P.; Sisler, H.D. Tricyclazole, pyroquilon, tetrachlorophthalide, PCBA, coumarin and related compounds inhibit melanization and epidermal penetration by Pyricularia oryzae. J. Pestic. Sci. Int. Ed. 1982, 7, 161–166. [Google Scholar] [CrossRef] [Green Version]
- Takano, Y.; Kubo, Y.; Kuroda, L.; Furusawa, I. Temporal transcriptional pattern of three melanin biosynthesis genes, PKS1, SCD1 and THR1, in appressorium-differentiating and nondifferentiating conidia of Colletotrichum lagenarium. Appl. Environ. Microbiol. 1997, 63, 351–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubo, Y.; Takano, Y.; Endo, N.; Yasuda, N.; Tajima, S.; Furusawa, I. Cloning and structural analysis of the melanin biosynthesis gene SCDl encoding scytalone dehydratase in Colletotrichum lagenarium. Appl. Environ. Microbiol. 1996, 6l62, 4340–4344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.; Chi, Z.; Liu, G.L.; Hu, Z.; Zhao, S.Z.; Chi, Z.M. Melanin biosynthesis in the desert-derived Aureobasidium melanogenum XJ5-1 is controlled mainly by the CWI signal pathway via a transcriptional activator Cmr1. Curr. Genet. 2020, 66, 173–185. [Google Scholar] [CrossRef]
- Casadevall, A. Melanin, melanin and composition in Cryptococcus neoformans. Infect. Immun. 1996, 64, 2420–2424. [Google Scholar]
- Nosanchuk, J.D.; Gomez, B.L.; Youngchim, S.; Diez, S.; Aisen, P.; Zancope-Oliveira, R.; Restrepo, A.; Casadevall, A.; Hamilton, A.J. Histoplasma capsulatum synthesizes melanin-Like pigments in vitro and during mammalian infection. Infect. Immun. 2002, 70, 5124–5131. [Google Scholar] [CrossRef] [Green Version]
- Morris-Jones, R.; Gomez, B.L.; Diez, S.; Uran, M.; Morris-Jones, S.D.; Casadevall, A.; Nosanchuk, J.D.; Hamilton, A.J. Synthesis of melanin pigment by Candida albicans in vitro and during infection. Infect. Immun. 2005, 73, 6147–6150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Nimse, S.B.; Mathew, D.E.; Dhimmar, A.; Sahastrabudhe, H.; Gajjar, A.; Ghadge, V.A.; Kumar, P.; Shinde, P.B. Microbial melanin: Recent advances in biosynthesis, extraction, characterization, and applications. Biotechnol. Adv. 2021, 53, 107773. [Google Scholar] [CrossRef] [PubMed]
- Chumley, F.; Valent, B. Genetic analysis of melanin-deficient, nonpathogenic mutants of Magnaporthe grisea. Mol. Plant Microbe Interact. 1990, 3, 135–143. [Google Scholar] [CrossRef]
- Kihara, J.; Moriwaki, A.; Ueno, M.; Tokunaga, T.; Arase, S.; Honda, Y. Cloning, functional analysis and expression of a scytalone dehydratase gene (SCD1) involved in melanin biosynthesis of the phytopathogenic fungus Bipolaris oryzae. Curr. Genet. 2004, 45, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Skory, C.D.; Chang, P.K.; Cary, J.; Linz, J.E. Isolation and characterization of a gene from Aspergillus parasiticus associated with the conversion of versicolorin A to sterigmatocystin in aflatoxin biosynthesis. Appl. Environ. Microbiol. 1992, 58, 3527–3537. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Lu, D.; Tian, C. Analysis of melanin biosynthesis in the plant pathogenic fungus Colletotrichum gloeosporioides. Fungal Biol. 2021, 125, 679–692. [Google Scholar] [CrossRef] [PubMed]
- Romero-Martinez, R.; Wheeler, M.; Guerrero-Plata, A.; Rico, G.; Torres-Guerrero, H. Biosynthesis and functions of melanin in Sporothrix schenckii. Infect. Immun. 2000, 68, 3696–3703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, S.; Cao, K.; Liu, N.; Meng, C.; Cao, Z.; Dai, D.; Jia, H.; Zang, J.; Li, Z.; Hao, Z.; et al. The StLAC2 gene is required for cell wall integrity, DHN-melanin synthesis and the pathogenicity of Setosphaeria turcica. Fungal Biol. 2017, 121, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.; Donofrio, N.; Pan, H.Q.; Coughlan, S.; Brown, D.E.; Meng, S.W.; Mitchell, T.; Dean, R.A. Transcriptome analysis reveals new insight into appressorium formation and function in the rice blast fungus Magnaporthe oryzae. Genome Biol. 2008, 9, R85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.; Yan, Y.; Qu, Y.; Wang, J.; Feng, X.; Liu, X.; Lin, F.; Lu, J.-P. Role refinement of melanin synthesis genes by gene knockout reveals their functional diversity in Pyricularia oryzae strains. Microbiol. Res. 2021, 242, 126620. [Google Scholar] [CrossRef]
- Langfelder, K.; Streibel, M.; Jahn, B.; Haase, G.; Brakhage, A.A. Biosynthesis of fungal melanins and their importance for human pathogenic fungi. Fungal Genet. Biol. 2003, 38, 143–158. [Google Scholar] [CrossRef] [PubMed]
- Pavan, M.E.; López, N.I.; Pettinari, M.J. Melanin biosynthesis in bacteria, regulation and production perspectives. Appl. Microbiol. Biotechnol. 2020, 104, 1357–1370. [Google Scholar] [CrossRef]
- Ismaya, W.T.; Rozeboom, H.J.; Weijn, A.; Mes, J.J.; Fusetti, F.; Wichers, H.J.; Dijkstra, B.W. Crystal structure of Agaricus bisporus mushroom tyrosinase: Identity of the tetramer subunits and interaction with tropolone. Biochemistry 2011, 50, 5477–5486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldfeder, M.; Egozy, M.; Shuster Ben-Yosef, V.; Adir, N.; Fishman, A. Changes in tyrosinase specificity by ionic liquids and sodium dodecyl sulfate. Appl. Microbiol. Biotechnol. 2013, 97, 1953–1961. [Google Scholar] [CrossRef] [PubMed]
- Pretzler, M.; Bijelic, A.; Rompel, A. Heterologous expression and characterization of functional mushroom tyrosinase (AbPPO4). Sci. Rep. 2017, 7, 1810. [Google Scholar] [CrossRef] [Green Version]
- Zolghadri, S.; Bahrami, A.; Hassan Khan, M.T.; Munoz-Munoz, J.; Garcia-Molina, F.; Garcia-Canovas, F.; Saboury, A.A. A comprehensive review on tyrosinase inhibitors. J. Enzym. Inhib. Med. Chem. 2019, 34, 279–309. [Google Scholar] [CrossRef] [Green Version]
- Cabezudo, I.; Ramallo, I.A.; Alonso, V.L.; Furlan, R.L. Effect directed synthesis of a new tyrosinase inhibitor with anti-browning activity. Food Chem. 2021, 341, 128232. [Google Scholar] [CrossRef]
- Rao, A.; Pimprikar, P.; Bendigiri, C.; Kumar, A.R.; Zinjarde, S. Cloning and expression of a tyrosinase from Aspergillus oryzae in Yarrowia lipolytica: Application in L-DOPA biotransformation. Appl. Microbiol. Biotechnol. 2011, 92, 951–959. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Zhang, N.; Yan, S.; Jiang, S.; Yin, H. A novel tyrosinase from Armillaria ostoyae with comparable monophenolase and diphenolase activities suffers substrate inhibition. Appl. Microbiol. Biotechnol. 2021, 87, e00275-21. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Liu, F.; Kong, Q.; Wu, Y.; He, Y.; Sang, H. Fungal melanin-induced metabolic reprogramming in macrophages is crucial for inflammation. J. Med. Mycol. 2023, 101359. [Google Scholar] [CrossRef] [PubMed]
- Boddy, L. Pathogens of Autotrophs. Fungi 2016, 3, 245–292. [Google Scholar]
- Gupta, L.; Vermani, M.; Kaur Ahluwalia, S.; Vijayaraghavan, P. Molecular virulence determinants of Magnaporthe oryzae: Disease pathogenesis and recent interventions for disease management in rice plant. Mycology 2021, 12, 174–187. [Google Scholar] [CrossRef]
- Batool, W.; Shabbir, A.; Lin, L.; Chen, X.; An, Q.; He, X.; Pan, S.; Chen, S.; Chen, Q.; Wang, Z.; et al. Translation initiation factor eIF4E positively modulates conidiogenesis, appressorium formation, host invasion and stress homeostasis in the filamentous fungi Magnaporthe oryzae. Front. Plant Sci. 2021, 12, 646343. [Google Scholar] [CrossRef]
- Batool, W.; Liu, C.; Fan, X.; Zhang, P.; Hu, Y.; Wei, Y.; Zhang, S.-H. AGC/AKT protein kinase SCH9 is critical to pathogenic development and overwintering survival in Magnaporthe oryzae. J. Fungi 2022, 8, 810. [Google Scholar] [CrossRef]
- Abdul, W.; Aliyu, S.R.; Lin, L.; Sekete, M.; Chen, X.; Otieno, F.J.; Yang, T.; Lin, Y.H.; Norvienyeku, J.; Wang, Z.-H. Family-four aldehyde dehydrogenases play an indispensable role in the pathogenesis of Magnaporthe oryzae. Front. Plant Sci. 2018, 9, 980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, Y.; Wei, Y.; Zhang, P.; Liu, X.; Li, X.; Wang, S.; Liang, H.; Zhang, S.-H. The bicarbonate transporter (MoAE4) localized on both cytomembrane and tonoplast promotes pathogenesis in Magnaporthe oryzae. J. Fungi 2021, 7, 955. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Jiang, H.; Zheng, Q.; Chen, X.; Wang, R.; Yang, S.; Zhao, G.; Liu, J.; Norvienyeku, J.; Wang, Z. Isopropylmalate isomerase MoLeu1 orchestrates leucine biosynthesis, fungal development, and pathogenicity in Magnaporthe oryzae. Appl. Microbiol. Biotechnol. 2019, 103, 327–337. [Google Scholar] [CrossRef]
- Nakamura, M.; Nakajima, T.; Ohba, Y.; Yamauchi, S.; Lee, B.-R.; Ichishima, E. Identification of copper ligands in Aspergillus oryzae tyrosinase by site-directed mutagenesis. Biochem. J. 2000, 350, 537–545. [Google Scholar] [CrossRef]
- Kanteev, M.; Goldfeder, M.; Chojnacki, M.; Adir, N.; Fishman, A. The mechanism of copper uptake by tyrosinase from Bacillus megaterium. J. Biol. Inorg. Chem. 2013, 18, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Noh, H.; Lee, S.; Jo, H.J.; Choi, H.W.; Hong, S.; Kong, K. Histidine residues at the copper-binding site in human tyrosinase are essential for its catalytic activities. J. Enzym. Inhib. Med. Chem. 2020, 35, 726–732. [Google Scholar] [CrossRef] [PubMed]
- Setaluri, V. Sorting and targeting of melanosomal membrane proteins: Signals, pathways, and mechanisms. Pigment. Cell Res. 2000, 13, 128–134. [Google Scholar] [CrossRef]
- Decker, H.; Schweikardt, T.; Nillius, D.; Salzbrunn, U.; Jaenicke, E.; Tuczek, F. Similar enzyme activation and catalysis in hemocyanins and tyrosinases. Gene 2007, 398, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Cordero, R.J.; Casadevall, A. Melanin. Curr. Biol. 2020, 30, 142–143. [Google Scholar] [CrossRef] [PubMed]
- Cordero, R.J.; Casadevall, A. Functions of fungal melanin beyond virulence. Fungal Biol. Rev. 2017, 31, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, C.W.; Koide, R.T. The function of melanin in the ectomycorrhizal fungus Cenococcum geophilum under water stress. Fungal Ecol. 2013, 6, 479–486. [Google Scholar] [CrossRef]
- Rangel Montoya, E.A.; Paolinelli, M.; Rolshausen, P.; Hernandez Martinez, R. The role of melanin in the grapevine trunk disease pathogen Lasiodiplodia gilanensis. Phytopathol. Mediterr. 2020, 59, 549–563. [Google Scholar]
- Kampatsikas, I.; Bijelic, A.; Pretzler, M.; Rompel, A. Three recombinantly expressed apple tyrosinases suggest the amino acids responsible for mono-versus diphenolase activity in plant polyphenol oxidases. Sci. Rep. 2017, 7, 8860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neupane, N.; Bhusal, K. A review of blast disease of rice in Nepal. J. Plant Pathol. Microbiol. 2021, 11, 528. [Google Scholar]
- Talbot, N.J. On the trail of a cereal killer: Exploring the biology of Magnaporthe grisea. Annu. Rev. Microbiol. 2003, 57, 177–202. [Google Scholar] [CrossRef] [Green Version]
- Pennisi, E. Armed and dangerous. Science 2010, 327, 804–805. [Google Scholar]
- Butler, M.J.; Day, A.W.; Henson, J.M.; Money, N.P. Pathogenic properties of fungal melanins. Mycologia 2001, 93, 1–8. [Google Scholar] [CrossRef]
- Nosanchuk, J.D.; Casadevall, A. The contribution of melanin to microbial pathogenesis. Cell. Microbiol. 2003, 5, 203–223. [Google Scholar] [CrossRef]
- Shi, H.; Meng, S.; Qiu, J.; Wang, C.; Shu, Y.; Luo, C.; Kou, Y. MoWhi2 regulates appressorium formation and pathogenicity via the MoTor signalling pathway in Magnaporthe oryzae. Mol. Plant Pathol. 2021, 22, 969–983. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Leung, H. Genetic analysis of sporulation in Magnaporthe grisea by chemical and insertional mutagenesis. Mol. Plant Microbe Interact. 1995, 8, 949–959. [Google Scholar] [CrossRef]
- Yang, J.; Zhao, X.; Sun, J.; Kang, Z.; Ding, S.; Xu, J.R.; Peng, Y.L. A novel protein Com1 is required for normal conidium morphology and full virulence in Magnaporthe oryzae. Mol. Plant Microbe Interact. 2010, 23, 112–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Li, G.; Lin, C.; He, C. Conidiophore stalk-less1 encodes a putative zinc-finger protein involved in the early stage of conidiation and mycelial infection in Magnaporthe oryzae. Mol. Plant Microbe Interact. 2009, 22, 402–410. [Google Scholar] [CrossRef] [Green Version]
- Dang, Y.; Wei, Y.; Batool, W.; Sun, X.; Li, X.; Zhang, S.-H. Contribution of the mitochondrial carbonic anhydrase (MoCA1) to conidiogenesis and pathogenesis in Magnaporthe oryzae. Front. Microbiol. 2022, 13, 845570. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Dagdas, Y.F.; Zhu, X.; Zheng, S.; Chen, L.; Cartwright, Z.; Talbot, N.J.; Wang, Z. The glycogen synthase kinase MoGsk1, regulated by Mps1 MAP kinase, is required for fungal development and pathogenicity in Magnaporthe oryzae. Sci. Rep. 2017, 7, 945. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Xie, S.; Zhao, X.; Chen, X.; Zheng, W.; Lu, G.; Xu, J.; Wang, Z. A homeobox gene is essential for conidiogenesis of the rice blast fungus Magnaporthe oryzae. Mol. Plant Microbe Interact. 2010, 23, 366–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, J.; Frases, S.; Wang, H.; Casadevall, A.; Stark, R.E. Following fungal melanin biosynthesis with solid-state NMR: Biopolymer molecular structures and possible connections to cell-wall polysaccharides. Biochemistry 2008, 47, 4701–4710. [Google Scholar] [CrossRef]
- Zhang, P.; Wang, X.N.; Fan, A.L.; Zheng, Y.J.; Liu, X.Z.; Wang, S.H.; Zou, H.X.; Oakley, B.R.; Keller, N.P.; Yin, W.B. A cryptic pigment biosynthetic pathway uncovered by heterologous expression is essential for conidial development in Pestalotiopsis fici. Mol. Microbiol. 2017, 105, 469–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filippova, S.N.; Kuznetsov, V.D.; Zaslavskaya, P.L. Production of melanins by Streptomyces galbus as a response to the elevated temperature of its cultivation and melanin localization. Mikrobiologiya 1987, 56, 710–712. [Google Scholar]
- Rosas, A.L.; Casadevall, A. Melanization affects susceptibility of Cryptococcus neoformans to heat and cold. FEMS Microbiol. Lett. 1997, 153, 265–272. [Google Scholar] [CrossRef]
- Foster, A.J.; Ryder, L.S.; Kershaw, M.J.; Talbot, N.J. The role of glycerol in the pathogenic lifestyle of the rice blast fungus Magnaporthe oryzae. Environ. Microbiol. 2017, 19, 1008–1016. [Google Scholar] [CrossRef] [Green Version]
- Qi, Z.; Wang, Q.I.; Dou, X.; Wang, W.; Zhao, Q.; Lv, R.; Zhang, H.; Zheng, X.; Wang, P.; Zhang, Z. MoSwi6, an APSES family transcription factor, interacts with MoMps1 and is required for hyphal and conidial morphogenesis, appressorial function and pathogenicity of Magnaporthe oryzae. Mol. Plant Pathol. 2012, 13, 677–689. [Google Scholar] [CrossRef]
- Ludwig, N.; Lohrer, M.; Hempel, M.; Mathea, S.; Schliebner, I.; Menzel, M.; Kiesow, A.; Schaffrath, U.; Deising, H.B.; Horbach, R. Melanin is not required for turgor generation but enhances cell-wall rigidity in appressoria of the corn pathogen Colletotrichum graminicola. Mol. Plant Microbe Interact. 2014, 27, 315–327. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Wei, Y.; Zhang, S.-H. The C3HC type zinc-finger protein (ZFC3) interacting with Lon/MAP1 is important for mitochondrial gene regulation, infection hypha development and longevity of Magnaporthe oryzae. BMC Microbiol. 2020, 20, 23. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, X.; Zhang, P.; Batool, W.; Liu, C.; Hu, Y.; Wei, Y.; He, Z.; Zhang, S.-H. Contribution of the Tyrosinase (MoTyr) to Melanin Synthesis, Conidiogenesis, Appressorium Development, and Pathogenicity in Magnaporthe oryzae. J. Fungi 2023, 9, 311. https://doi.org/10.3390/jof9030311
Fan X, Zhang P, Batool W, Liu C, Hu Y, Wei Y, He Z, Zhang S-H. Contribution of the Tyrosinase (MoTyr) to Melanin Synthesis, Conidiogenesis, Appressorium Development, and Pathogenicity in Magnaporthe oryzae. Journal of Fungi. 2023; 9(3):311. https://doi.org/10.3390/jof9030311
Chicago/Turabian StyleFan, Xiaoning, Penghui Zhang, Wajjiha Batool, Chang Liu, Yan Hu, Yi Wei, Zhengquan He, and Shi-Hong Zhang. 2023. "Contribution of the Tyrosinase (MoTyr) to Melanin Synthesis, Conidiogenesis, Appressorium Development, and Pathogenicity in Magnaporthe oryzae" Journal of Fungi 9, no. 3: 311. https://doi.org/10.3390/jof9030311
APA StyleFan, X., Zhang, P., Batool, W., Liu, C., Hu, Y., Wei, Y., He, Z., & Zhang, S. -H. (2023). Contribution of the Tyrosinase (MoTyr) to Melanin Synthesis, Conidiogenesis, Appressorium Development, and Pathogenicity in Magnaporthe oryzae. Journal of Fungi, 9(3), 311. https://doi.org/10.3390/jof9030311