
Arbuscular Mycorrhizal Fungi Improve the Growth, Water Status, and Nutrient Uptake of Cinnamomum migao and the Soil Nutrient Stoichiometry under Drought Stress and Recovery

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Growth Substrate, AMF, and Seeds
2.3. Experimental Design
2.4. Specimen Collection
2.5. Plant Property Analysis
2.5.1. AMF Colonization Rate
2.5.2. Plant Growth and Water Status Analysis
2.5.3. Nutrient Analysis
2.6. Soil Property Analysis
2.7. Statistical Analyses
3. Results
3.1. AMF Colonization Rate
3.2. Plant Water Status
3.3. Plant Growth
3.4. Plant C, N, and P Contents
3.5. Plant C:N:P Ratios
3.6. Soil C, N, and P Contents
3.7. Soil C:N:P Ratios
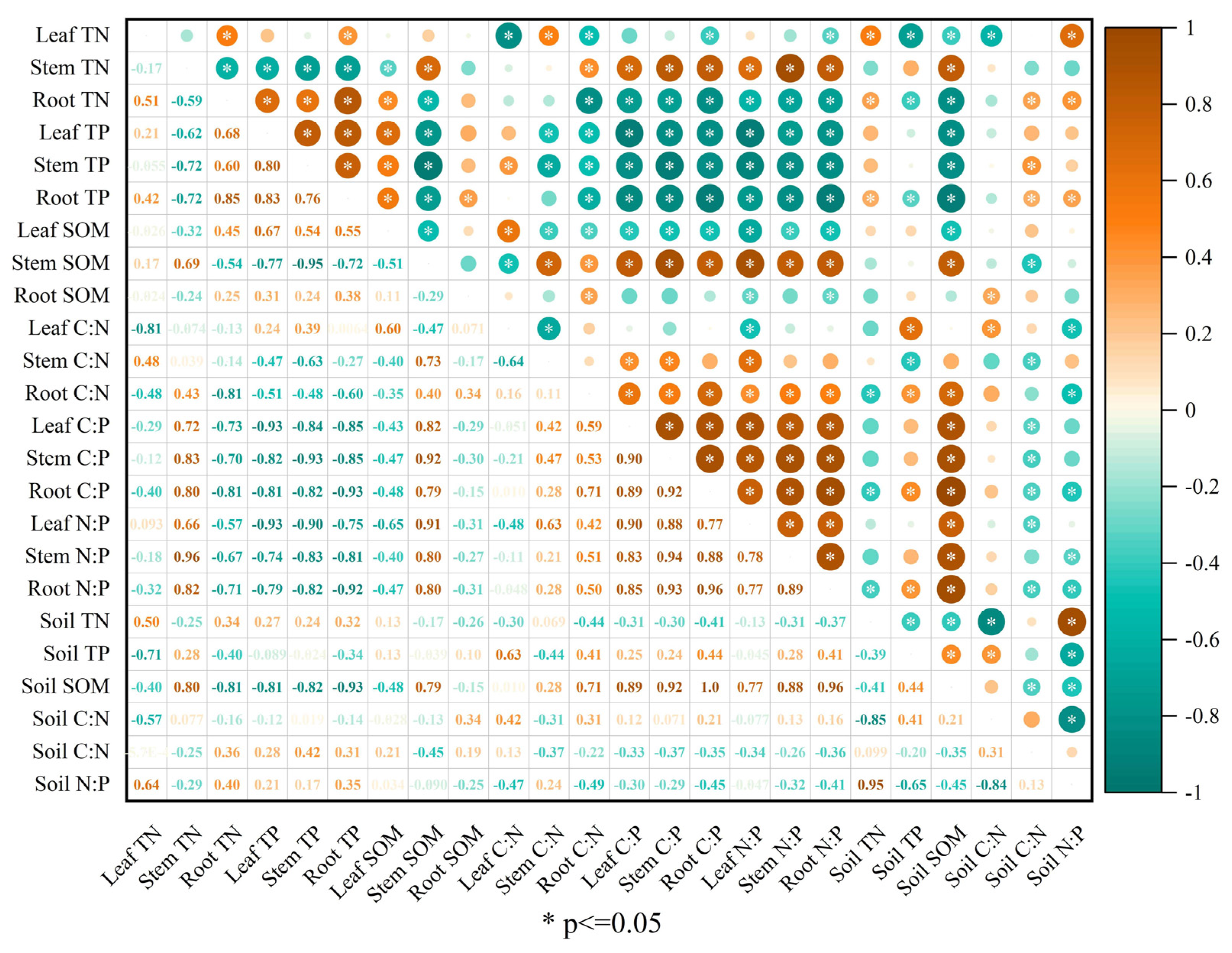
3.8. Relationships among Plant and Soil Nutrient Stoichiometries
3.9. Multivariate Statistical Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaisermann, A.; Vries, F.T.; Griffiths, R.I.; Bardgett, R.D. Legacy effects of drought on plant–soil feedbacks and plant–plant interactions. New Phytol. 2017, 215, 1413–1424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catalin, S.I.; Ionut, M. Drought phenomena and groundwater scarcity in eastern Romania (Siret-Prut region). EGU Gen. Assem. 2013, 15, 6997. [Google Scholar]
- Yuan, Z.; Jiao, F.; Shi, X.; Sardans, J.; Maestre, F.T.; Delgado-Baquerizo, M.; Reich, P.B.; Peñuelas, J. Experimental and observational studies find contrasting responses of soil nutrients to climate change. Elife 2017, 6, e23255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, W.; Xu, C.; Ma, W.; Yue, X.; Liang, X.; Zuo, X.; Knapp, A.K.; Smith, M.D.; Sardans, J.; Dijkstra, F.A.; et al. Effects of extreme drought on plant nutrient uptake and resorption in rhizomatous vs bunchgrass-dominated grasslands. Oecologia 2018, 188, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Zuo, X.; Ma, W.; Xu, C.; Li, A.; Yu, Q.; Knapp, A.K.; Tognetti, R.; Dijkstra, F.A.; Li, M.-H.; et al. Differential responses of canopy nutrients to experimental drought along a natural aridity gradient. Ecology 2018, 99, 2230–2239. [Google Scholar] [CrossRef] [Green Version]
- Begum, N.; Akhtar, K.; Ahanger, M.A.; Iqbal, M.; Wang, P.; Mustafa, N.S.; Zhang, L. Arbuscular mycorrhizal fungi improve growth, essential oil, secondary metabolism, and yield of tobacco (Nicotiana tabacum L.) under drought stress conditions. Environ. Sci. Pollut. Res. 2021, 28, 45276–45295. [Google Scholar] [CrossRef]
- Symanczik, S.; Lehmann, M.F.; Wiemken, A.; Boller, T.; Courty, P.-E. Effects of two contrasted arbuscular mycorrhizal fungal isolates on nutrient uptake by Sorghum bicolor under drought. Mycorrhiza 2018, 28, 779–785. [Google Scholar] [CrossRef]
- Zhang, H.; Churchill, A.C.; Anderson, I.C.; Igwenagu, C.; Power, S.A.; Plett, J.M.; Macdonald, C.A.; Pendall, E.; Carrillo, Y.; Powell, J.R. Ecological stoichiometry and fungal community turnover reveal variation among mycorrhizal partners in their responses to warming and drought. Mol. Ecology. 2023, 32, 229–243. [Google Scholar] [CrossRef]
- Van de Waal, D.B.; Elser, J.J.; Martiny, A.C.; Sterner, R.W.; Cotner, J.B. Editorial: Progress in Ecological Stoichiometry. Front. Microbiol. 2018, 9, 1957. [Google Scholar] [CrossRef] [Green Version]
- Du, E.; Terrer, C.; Pellegrini, A.; Ahlström, A.; Van Lissa, C.J.; Zhao, X.; Xia, N.; Wu, X.; Jackson, R.B. Global patterns of terrestrial nitrogen and phosphorus limitation. Nat. Geosci. 2020, 13, 221–226. [Google Scholar] [CrossRef]
- Hao, Q.; Song, Z.; Zhang, X.; Li, Q.; Yang, W.; Yang, S.; Tan, Q. Effects of Si on N and P stoichiometry in degraded grassland of northern China. Land Degrad. Dev. 2022, 33, 960–973. [Google Scholar] [CrossRef]
- Sun, Y.; Liao, J.; Zou, X.; Xu, X.; Yang, J.; Chen, H.Y.; Ruan, H. Coherent responses of terrestrial C:N stoichiometry to drought across plants, soil, and microorganisms in forests and grasslands. Agric. For. Meteorol. 2020, 292, 108104. [Google Scholar] [CrossRef]
- Zhao, W.; Reich, P.B.; Yu, Q.; Zhao, N.; Yin, C.; Zhao, C.; Li, D.; Hu, J.; Li, T.; Yin, H.; et al. Shrub type dominates the vertical distribution of leaf C:N:P stoichiometry across an extensive altitudinal gradient. Biogeosciences 2018, 15, 2033–2053. [Google Scholar] [CrossRef] [Green Version]
- Guo, F.; Jiang, G.; Yuan, D.; Polk, J.S. Evolution of major environmental geological problems in karst areas of Southwestern China. Environ. Earth Sci. 2012, 69, 2427–2435. [Google Scholar] [CrossRef]
- Brinkmann, R.; Parise, M. Karst Environments: Problems, Management, Human Impacts, and Sustainability: An Introduction to the Special Issue. J. Cave Karst Stud. 2012, 74, 135–136. [Google Scholar] [CrossRef]
- Shi, P.; Duan, J.; Zhang, Y.; Li, P.; Wang, X.; Li, Z.; Xiao, L.; Xu, G.; Lu, K.; Cheng, S.; et al. The effects of ecological construction and topography on soil organic carbon and total nitrogen in the Loess Plateau of China. Environ. Earth Sci. 2018, 78, 5. [Google Scholar] [CrossRef]
- Huang, X.; Tian, T.; Chen, J.; Wang, D.; Tong, B.; Liu, J. Transcriptome analysis of Cinnamomum migao seed germination in medicinal plants of Southwest China. BMC Plant Biol. 2021, 21, 1–21. [Google Scholar] [CrossRef]
- Li, L.X. Study on Cinnamomum migao Population Characteristics in Guizhou. Master’s Thesis, Guizhou University, Guizhou, China, 2017. [Google Scholar]
- Chen, J.-Z.; Huang, X.-L.; Xiao, X.-F.; Liu, J.-M.; Liao, X.-F.; Sun, Q.-W.; Peng, L.; Zhang, L. Seed Dormancy Release and Germination Requirements of Cinnamomum migao, an Endangered and Rare Woody Plant in Southwest China. Front. Plant Sci. 2022, 13, 11. [Google Scholar] [CrossRef]
- Huang, X.; Chen, J.; Liu, J.; Li, J.; Wu, M.; Tong, B. Autotoxicity Hinders the Natural Regeneration of Cinnamomum migao H. W. Li in Southwest China. Forests 2019, 10, 919. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Liu, J.M.; Wen, A.H.; Deng, M.M.; Xiong, X.; Liu, J.J. Simulated photosynthetic responses of Cinnamomum migao during drought stress evaluated using Light-response Models. Acta Ecol. Sin. 2019, 39, 913–922, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Xiao, X.; Chen, J.; Liao, X.; Yan, Q.; Liang, G.; Liu, J.; Wang, D.; Guan, R. Different Arbuscular Mycorrhizal Fungi Established by Two Inoculation Methods Improve Growth and Drought Resistance of Cinnamomum migao Seedlings Differently. Biology 2022, 11, 220. [Google Scholar] [CrossRef] [PubMed]
- Dai, R.; Zhong, J. Spatial and temporal variation characteristics of drought in Guizhou Province based on VCI and division of arid regions. Pearl River 2021, 42, 34–40. (In Chinese) [Google Scholar] [CrossRef]
- Staddon, P.L.; Gregersen, R.; Jakobsen, I. The response of two Glomus mycorrhizal fungi and a fine endophyte to elevated atmospheric CO2, soil warming and drought. Glob. Chang. Biol. 2004, 10, 1909–1921. [Google Scholar] [CrossRef]
- De Vries, F.T.; Griffiths, R.I.; Knight, C.G.; Nicolitch, O.; Williams, A. Harnessing rhizosphere microbiomes for drought-resilient crop production. Science 2020, 368, 270–274. [Google Scholar] [CrossRef]
- Guo, Y.; Gao, P.; Li, F.; Duan, T. Effects of AM fungi and grass endophytes on perennial ryegrass Bipolaris sorokiniana leaf spot disease under limited soil nutrients. Eur. J. Plant Pathol. 2019, 154, 659–671. [Google Scholar] [CrossRef]
- Li, F.; Guo, Y.; Christensen, M.J.; Gao, P.; Li, Y.; Duan, T. An arbuscular mycorrhizal fungus and Epichloë festucae var. lolii reduce Bipolaris sorokiniana disease incidence and improve perennial ryegrass growth. Mycorrhiza 2017, 28, 159–169. [Google Scholar] [CrossRef]
- Van Der Heijden, M.G.A.; Martin, F.M.; Selosse, M.-A.; Sanders, I.R. Mycorrhizal ecology and evolution: The past, the present, and the future. New Phytol. 2015, 205, 1406–1423. [Google Scholar] [CrossRef]
- Smith, S.E.; Jakobsen, I.; Grønlund, M.; Smith, F.A. Roles of arbuscular mycorrhizas in plant phosphorus nutrition: Interactions between pathways of phosphorus uptake in arbuscular mycorrhizal roots have important implications for understanding and manipulating plant phosphorus acquisition. Plant Physiol. 2011, 156, 1050–1057. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.E.; Facelli, E.; Pope, S.; Smith, F.A. Plant performance in stressful environments: Interpreting new and established knowledge of the roles of arbuscular mycorrhizas. Plant Soil 2009, 326, 3–20. [Google Scholar] [CrossRef]
- del Mar Alguacil, M.; Lozano, Z.; Campoy, M.J.; Roldán, A. Phosphorus fertilization management modifies the biodiversity of am fungi in a tropical savanna forage system. Soil Biol. Biochem. 2010, 42, 1114–1122. [Google Scholar] [CrossRef]
- Kaur, S.; Suseela, V. Unraveling Arbuscular Mycorrhiza-Induced Changes in Plant Primary and Secondary Metabolome. Metabolites 2020, 10, 335. [Google Scholar] [CrossRef] [PubMed]
- Heflish, A.A.; Hanfy, A.E.; Ansari, M.J.; Dessoky, E.S.; Attia, A.O.; Elshaer, M.M.; Gaber, M.K.; Kordy, A.; Doma, A.S.; Abdelkhalek, A.; et al. Green biosynthesized silver nanoparticles using Acalypha wilkesiana extract control root-knot nematode. J. King Saud Univ.-Sci. 2021, 33, 101516. [Google Scholar] [CrossRef]
- Püschel, D.; Bitterlich, M.; Rydlová, J.; Jansa, J. Drought accentuates the role of mycorrhiza in phosphorus uptake. Soil Biol. Biochem. 2021, 157, 108243. [Google Scholar] [CrossRef]
- Bowles, T.M.; Jackson, L.E.; Cavagnaro, T.R. Mycorrhizal fungi enhance plant nutrient acquisition and modulate nitrogen loss with variable water regimes. Glob. Chang. Biol. 2017, 24, e171–e182. [Google Scholar] [CrossRef] [PubMed]
- Hashem, A.; Kumar, A.; Al-Dbass, A.M.; Alqarawi, A.A.; Al-Arjani, A.-B.F.; Singh, G.; Farooq, M.; Abd_Allah, E.F. Arbuscular mycorrhizal fungi and biochar improves drought tolerance in chickpea. Saudi J. Biol. Sci. 2018, 26, 614–624. [Google Scholar] [CrossRef] [PubMed]
- Bücking, H.; Shachar-Hill, Y. Phosphate uptake, transport and transfer by the arbuscular mycorrhizal fungus Glomus intraradices is stimulated by increased carbohydrate availability. New Phytol. 2005, 165, 899–912. [Google Scholar] [CrossRef]
- Konvalinková, T.; Püschel, D.; Řezáčová, V.; Gryndlerová, H.; Jansa, J. Carbon flow from plant to arbuscular mycorrhizal fungi is reduced under phosphorus fertilization. Plant Soil 2017, 419, 319–333. [Google Scholar] [CrossRef]
- Liu, H.; Chen, W.; Wu, M.; Wu, R.; Zhou, Y.; Gao, Y.; Ren, A. Arbuscular mycorrhizal fungus inoculation reduces the drought-resistance advantage of endophyte-infected versus endophyte-free Leymus chinensis. Mycorrhiza 2017, 27, 791–799. [Google Scholar] [CrossRef]
- Wei, Y.; Wang, S.; Liu, X.; Huang, T. Genetic diversity of arbuscular mycorrhizal fungi in karst microhabitats of Guizhou Province, China. Chin. J. Plant Ecol. 2012, 35, 1083–1090, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Liang, Y.; Pan, F.; He, X.; Chen, X.; Su, Y. Effect of vegetation types on soil arbuscular mycorrhizal fungi and nitrogen-fixing bacterial communities in a karst region. Environ. Sci. Pollut. Res. 2016, 23, 18482–18491. [Google Scholar] [CrossRef]
- Hui, N.; Sun, N.; Du, H.; Umair, M.; Kang, H.; Liu, X.; Romantschuk, M.; Liu, C. Karst rocky desertification does not erode ectomycorrhizal fungal species richness but alters microbial community structure. Plant Soil 2019, 445, 383–396. [Google Scholar] [CrossRef]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161, IN16–IN18. [Google Scholar] [CrossRef]
- He, J.; Dong, T.; Wu, H.; Zou, Y.; Wu, Q.; Kamil, K. Mycorrhizas induce diverse responses of root TIP aquaporin gene expression to drought stress in trifoliate orange. Sci. Hortic. 2019, 243, 64–69. [Google Scholar] [CrossRef]
- Castillo, F.J. Antioxidative protection in the inducible CAM plant Sedum album L. following the imposition of severe water stress and recovery. Oecologia 1996, 107, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Yeomans, J.C.; Bremner, J.M. A rapid and precise method for routine determination of organic carbon in soil. Commun. Soil Sci. Plant Anal. 1988, 19, 1467–1476. [Google Scholar] [CrossRef]
- Thomas, R.L.; Sheard, R.W.; Moyer, J.R. Comparison of conventional and automated procedures for nitrogen, phosphorus, and potassium analysis of plant material using a individual digestion. Agron. J. 1967, 59, 240–243. [Google Scholar] [CrossRef]
- State Forestry Administration, P.R.C. Determination of Total Nitrogen, Phosphorus, Potassium, Sodium, Calcium, Magnesium in Forest Plant and Forest Floor; LY/T 1271-1999; State Forestry Administration: Beijing, China, 1999.
- Su, H.; Cui, J.; Adamowski, J.F.; Zhang, X.; Biswas, A.; Cao, J. Using Leaf Ecological Stoichiometry to Direct the Management of Ligularia virgaurea on the Northeast Qinghai-Tibetan Plateau. Front. Environ. Sci. 2022, 9, 805405. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon and organic matter. Methods Soil Anal. Part 3 Chem. Methods 1966, 5, 961–1010. [Google Scholar] [CrossRef]
- Bray, R.H.; Kurtz, L.T. Determination of total, organic, and available forms of phosphorus in soils. Soil Sci. 1945, 59, 39–46. [Google Scholar] [CrossRef]
- Liu, T.; Li, Z.; Hui, C.; Tang, M.; Zhang, H. Effect of Rhizophagus irregularis on osmotic adjustment, antioxidation and aquaporin PIP genes expression of Populus × canadensis ‘Neva’ under drought stress. Acta Physiol. Plant. 2016, 38, 1–9. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, J.; Huang, Y. Effects of arbuscular mycorrhizal fungi on the drought tolerance of Cyclobalanopsis glauca seedlings under greenhouse conditions. New For. 2014, 45, 545–556. [Google Scholar] [CrossRef]
- Sánchez-Castro, I.; Ferrol, N.; Barea, J. Analyzing the community composition of arbuscular mycorrhizal fungi colonizing the roots of representative shrubland species in a Mediterranean ecosystem. J. Arid. Environ. 2012, 80, 1–9. [Google Scholar] [CrossRef]
- Sánchez-Castro, I.; Ferrol, N.; Cornejo, P.; Barea, J.M. Temporal dynamics of arbuscular mycorrhizal fungi colonizing roots of representative shrub species in a semi-arid Mediterranean ecosystem. Mycorrhiza 2011, 22, 449–460. [Google Scholar] [CrossRef]
- Köhl, L.; van der Heijden, M.G. Arbuscular mycorrhizal fungal species differ in their effect on nutrient leaching. Soil Biol. Biochem. 2016, 94, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Cavagnaro, T.R.; Gao, L.L.; Smith, F.A.; Smith, S.E. Morphology of Arbuscular Mycorrhizas is Influenced by Fungal Identity. New Phytol. 2001, 151, 469–475. [Google Scholar] [CrossRef]
- Tatsumi, C.; Taniguchi, T.; Du, S.; Yamanaka, N.; Tateno, R. Soil nitrogen cycling is determined by the competition between mycorrhiza and ammonia-oxidizing prokaryotes. Ecology 2019, 101, e02963. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekaran, M.; Chanratana, M.; Kim, K.; Seshadri, S.; Sa, T. Impact of Arbuscular Mycorrhizal Fungi on Photosynthesis, Water Status, and Gas Exchange of Plants Under Salt Stress–A Meta-Analysis. Front. Plant Sci. 2019, 10, 457. [Google Scholar] [CrossRef]
- Marschner, P.; Baumann, K. Changes in bacterial community structure induced by mycorrhizal colonisation in split-root maize. Plant Soil 2003, 251, 279–289. [Google Scholar] [CrossRef]
- Aslam, M.M.; Okal, E.J.; Idris, A.L.; Qian, Z.; Xu, W.; Karanja, J.K.; Wani, S.H.; Yuan, W. Rhizosphere microbiomes can regulate plant drought tolerance. Pedosphere 2021, 32, 61–74. [Google Scholar] [CrossRef]
- Hodge, A. Root decisions. Plant Cell Environ. 2009, 32, 628–640. [Google Scholar] [CrossRef]
- Yang, G.; Guo, L.P.; Guo, X.H.; Chen, M.; Chen, M.L.; Zhou, J. Selectivity Infection of Arbuscular Mycorrhizal Fungi in Medicinal Plants. Chin. J. Inf. Tradit Chin. Med. 2012, 19, 53–55, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Wei, Z.H.; Zhao, S.X.; Li, Z.W.; Zhao, J.J.; Zhou, N.; Guo, D.Q. Effects of Mixed Inoculation of Different Arbuscular Mycorrhizal Fungi on Saussurea Costus Rhizosphere Microorganisms and Soil Enzyme Activities. Chin. Wild Plant Resour. 2021, 40, 6–11, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Segal, L.M.; Wilson, R.A. Reactive oxygen species metabolism and plant-fungal interactions. Fungal Genet. Biol. 2018, 110, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, Y.-N.; Wu, Q.-S.; Kuča, K. Unravelling the role of arbuscular mycorrhizal fungi in mitigating the oxidative burst of plants under drought stress. Plant Biol. 2021, 23, 50–57. [Google Scholar] [CrossRef]
- Morte, A.; Lovisolo, C.; Schubert, A. Effect of drought stress on growth and water relations of the mycorrhizal association Helianthemum almeriense-Terfezia claveryi. Mycorrhiza 2000, 10, 115–119. [Google Scholar] [CrossRef]
- Porcel, R.; Gómez, M.; Kaldenhoff, R.; Ruiz-Lozano, J.M. Impairment of NtAQP1 gene expression in tobacco plants does not affect root colonisation pattern by arbuscular mycorrhizal fungi but decreases their symbiotic efficiency under drought. Mycorrhiza 2005, 15, 417–423. [Google Scholar] [CrossRef]
- Jördens, C.; Scheller, M.; Breitenstein, B.; Selmar, D.; Koch, M. Evaluation of leaf water status by means of permittivity at terahertz frequencies. J. Biol. Phys. 2009, 35, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Vurukonda, S.S.K.P.; Vardharajula, S.; Shrivastava, M.; SkZ, A. Multifunctional Pseudomonas putida strain FBKV2 from arid rhizosphere soil and its growth promotional effects on maize under drought stress. Rhizosphere 2016, 1, 4–13. [Google Scholar] [CrossRef]
- Huang, Z.; Zou, Z.; He, C.; He, Z.; Zhang, Z.; Li, J. Physiological and photosynthetic responses of melon (Cucumis melo L.) seedlings to three Glomus species under water deficit. Plant Soil 2010, 339, 391–399. [Google Scholar] [CrossRef]
- Gong, M.; Tang, M.; Chen, H.; Zhang, Q.; Feng, X. Effects of two Glomus species on the growth and physiological performance of Sophora davidii seedlings under water stress. New For. 2012, 44, 399–408. [Google Scholar] [CrossRef]
- Chareesri, A.; De Deyn, G.B.; Sergeeva, L.; Polthanee, A.; Kuyper, T.W. Increased arbuscular mycorrhizal fungal colonization reduces yield loss of rice (Oryza sativa L.) under drought. Mycorrhiza 2020, 30, 315–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bárzana, G.; Aroca, R.; Paz, J.A.; Chaumont, F.; Martinez-Ballesta, M.C.; Carvajal, M.; Ruiz-Lozano, J.M. Arbuscular mycorrhizal symbiosis increases relative apoplastic water flow in roots of the host plant under both well-watered and drought stress conditions. Ann. Bot. 2012, 109, 1009–1017. [Google Scholar] [CrossRef] [PubMed]
- Augé, R.M.; Toler, H.D.; Saxton, A. Arbuscular mycorrhizal symbiosis alters stomatal conductance of host plants more under drought than under amply watered conditions: A meta-analysis. Mycorrhiza 2014, 25, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Muthukumar, T.; Udaiyan, K. Growth response and nutrient utilization of Casuarina equisetifolia seedlings inoculated with bioinoculants under tropical nursery conditions. New For. 2010, 40, 101–118. [Google Scholar] [CrossRef]
- Anamala, P.; Sultana, U.; Sindhura, P.; Gul, M.Z. Plant GrowthPromoting Rhizobacteria (PGPR): A unique strategy for sustainable agriculture. In Handbook of Research on Microbial Remediation and Microbial Biotechnology for Sustainable Soil; IGI Global: Hershey, PA, USA, 2021; pp. 332–357. [Google Scholar] [CrossRef]
- Shaffique, S.; Khan, M.A.; Imran, M.; Kang, S.-M.; Park, Y.-S.; Wani, S.H.; Lee, I.-J. Research Progress in the Field of Microbial Mitigation of Drought Stress in Plants. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef]
- Saboor, A.; Ali, M.A.; Hussain, S.; El Enshasy, H.A.; Hussain, S.; Ahmed, N.; Gafur, A.; Sayyed, R.; Fahad, S.; Danish, S.; et al. Zinc nutrition and arbuscular mycorrhizal symbiosis effects on maize (Zea mays L.) growth and productivity. Saudi J. Biol. Sci. 2021, 28, 6339–6351. [Google Scholar] [CrossRef]
- Grümberg, B.C.; Urcelay, C.; Shroeder, M.A.; Vargas-Gil, S.; Luna, C.M. The role of inoculum identity in drought stress mitigation by arbuscular mycorrhizal fungi in soybean. Biol. Fertil. Soils 2014, 51, 1–10. [Google Scholar] [CrossRef]
- Al-Arjani, A.-B.F.; Hashem, A.; Abd_Allah, E.F. Arbuscular mycorrhizal fungi modulates dynamics tolerance expression to mitigate drought stress in Ephedra foliata Boiss. Saudi J. Biol. Sci. 2019, 27, 380–394. [Google Scholar] [CrossRef]
- Ortas, I. Mycorrhizas in fruit nutrition: Important breakthroughs. In Fruit Crops; Diagnosis and Management of Nutrient Constraints; Srivastava, A.K., Hu, C.X., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 339–351. [Google Scholar] [CrossRef]
- Gholamhoseini, M.; Ghalavand, A.; Dolatabadian, A.; Jamshidi, E.; Khodaei-Joghan, A. Effects of arbuscular mycorrhizal inoculation on growth, yield, nutrient uptake and irrigation water productivity of sunflowers grown under drought stress. Agric. Water Manag. 2013, 117, 106–114. [Google Scholar] [CrossRef]
- Wang, L.; Chen, X.; Du, Y.; Zhang, D.; Tang, Z. Nutrients Regulate the Effects of Arbuscular Mycorrhizal Fungi on the Growth and Reproduction of Cherry Tomato. Front. Microbiol. 2022, 13, 843010. [Google Scholar] [CrossRef]
- Doubková, P.; Vlasáková, E.; Sudová, R. Arbuscular mycorrhizal symbiosis alleviates drought stress imposed on Knautia arvensis plants in serpentine soil. Plant Soil 2013, 370, 149–161. [Google Scholar] [CrossRef]
- Ran, Z.; Yang, X.; Zhang, Y.; Zhou, J.; Guo, L. Effects of arbuscular mycorrhizal fungi on photosynthesis and biosynthesis of ginsenoside in Panax quinquefolius L. Theor. Exp. Plant Physiol. 2021, 33, 235–248. [Google Scholar] [CrossRef]
- Ghani, M.I.; Ali, A.; Atif, M.J.; Ali, M.; Amin, B.; Cheng, Z. Arbuscular Mycorrhizal Fungi and Dry Raw Garlic Stalk Amendment Alleviate Continuous Monocropping Growth and Photosynthetic Declines in Eggplant by Bolstering Its Antioxidant System and Accumulation of Osmolytes and Secondary Metabolites. Front. Plant Sci. 2022, 13, 849521. [Google Scholar] [CrossRef] [PubMed]
- Etesami, H.; Jeong, B.R.; Glick, B.R. Contribution of Arbuscular Mycorrhizal Fungi, Phosphate–Solubilizing Bacteria, and Silicon to P Uptake by Plant. Front. Plant Sci. 2021, 12, 699618. [Google Scholar] [CrossRef]
- Ilyas, F.; Ali, M.A.; Modhish, A.; Ahmed, N.; Hussain, S.; Bilal, M.; Arshad, M.; Danish, S.; Ghoneim, A.M.; Ilyas, A.; et al. Synchronisation of zinc application rates with arbuscular mycorrhizal fungi and phosphorus to maximise wheat growth and yield in zinc-deficient soil. Crop. Pasture Sci. 2022, 74, 157–172. [Google Scholar] [CrossRef]
- Oliveira, T.C.; Cabral, J.S.R.; Santana, L.R.; Tavares, G.G.; Santos, L.D.S.; Paim, T.P.; Müller, C.; Silva, F.G.; Costa, A.C.; Souchie, E.L.; et al. The arbuscular mycorrhizal fungus Rhizophagus clarus improves physiological tolerance to drought stress in soybean plants. Sci. Rep. 2022, 12, 1–15. [Google Scholar] [CrossRef]
- Pereira, C.M.R.; López-García, Á.; Maia, L.C.; Frøslev, T.G.; Kjøller, R.; Rosendahl, S. Arbuscular mycorrhizal fungal communities of pristine rainforests and adjacent sugarcane fields recruit from different species pools. Soil Biol. Biochem. 2022, 167, 108585. [Google Scholar] [CrossRef]
- Li, Y.L.; Jin, Z.X.; Luo, G.Y.; Chen, C.; Sun, Z.S.; Wang, X.Y. Effects of arbuscular mycorrhizal fungi inoculation on non-structural carbohydrate contents and C:N:P stoichiometry of Heptacodium miconioides under drought stress. Ying Yong Sheng Tai Xue Bao=J. Appl. Ecology. 2022, 33, 963–971, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Elser, J.J.; Acharya, K.; Kyle, M.; Cotner, J.B.; Makino, W.; Markow, T.; Watts, T.; Hobbie, S.E.; Fagan, W.; Schade, J.; et al. Growth rate-stoichiometry couplings in diverse biota. Ecol. Lett. 2003, 6, 936–943. [Google Scholar] [CrossRef] [Green Version]
- Walder, F.; Niemann, H.; Natarajan, M.; Lehmann, M.; Boller, T.; Wiemken, A. Mycorrhizal Networks: Common Goods of Plants Shared under Unequal Terms of Trade. Plant Physiol. 2012, 159, 789–797. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Tienda, J.; Valderas, A.; Camañes, G.; Agustín, P.G.; Ferrol, N. Kinetics of NH+4 uptake by the arbuscular mycorrhizal fungus Rhizophagus irregularis. Mycorrhiza 2012, 22, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Leigh, J.; Hodge, A.; Fitter, A.H. Arbuscular mycorrhizal fungi can transfer substantial amounts of nitrogen to their host plant from organic material. New Phytol. 2009, 181, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Ingraffia, R.; Saia, S.; Giovino, A.; Amato, G.; Badagliacca, G.; Giambalvo, D.; Martinelli, F.; Ruisi, P.; Frenda, A.S. Addition of high C:N crop residues to a P-limited substrate constrains the benefits of arbuscular mycorrhizal symbiosis for wheat P and N nutrition. Mycorrhiza 2021, 31, 441–454. [Google Scholar] [CrossRef] [PubMed]
- Bücking, H.; Kafle, A. Role of arbuscular mycorrhizal fungi in the nitrogen uptake of plants: Current knowledge and research gaps. Agronomy 2015, 5, 587–612. [Google Scholar] [CrossRef] [Green Version]
- Del-Saz, N.F.; Romero-Munar, A.; Cawthray, G.R.; Aroca, R.; Baraza, E.; Flexas, J.; Lambers, H.; Ribas-Carbó, M. Arbuscular mycorrhizal fungus colonization in Nicotiana tabacum decreases the rate of both carboxylate exudation and root respiration and increases plant growth under phosphorus limitation. Plant Soil 2017, 416, 97–106. [Google Scholar] [CrossRef]
- Wang, Y.; Lin, J.; Yang, F.; Tao, S.; Yan, X.; Zhou, Z.; Zhang, Y. Arbuscular mycorrhizal fungi improve the growth and performance in the seedlings of Leymus chinensis under alkali and drought stresses. Peerj 2022, 10, e12890. [Google Scholar] [CrossRef]
- Huang, Z.; Liu, B.; Davis, M.; Sardans, J.; Peñuelas, J.; Billings, S. Long-term nitrogen deposition linked to reduced water use efficiency in forests with low phosphorus availability. New Phytol. 2015, 210, 431–442. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Pan, G.; Li, L.; Hu, Z.; Wang, X. Leaf N/P ratio and nutrient reuse between dominant species and stands: Predicting phosphorus deficiencies in Karst ecosystems, southwestern China. Environ. Earth Sci. 2011, 64, 299–309. [Google Scholar] [CrossRef]
- Lambers, H.; Shane, M.W.; Cramer, M.D.; Pearse, S.J.; Veneklaas, E.J. Root Structure and Functioning for Efficient Acquisition of Phosphorus: Matching Morphological and Physiological Traits. Ann. Bot. 2006, 98, 693–713. [Google Scholar] [CrossRef] [Green Version]
- Pold, G.; Kwiatkowski, B.L.; Rastetter, E.B.; Sistla, S.A. Sporadic P limitation constrains microbial growth and facilitates SOM accumulation in the stoichiometrically coupled, acclimating microbe–plant–soil model. Soil Biol. Biochem. 2021, 165, 108489. [Google Scholar] [CrossRef]
- Leake, J.; Johnson, D.; Donnelly, D.; Muckle, G.; Boddy, L.; Read, D. Networks of power and influence: The role of mycorrhizal mycelium in controlling plant communities and agroecosystem functioning. Can. J. Bot. 2004, 82, 1016–1045. [Google Scholar] [CrossRef] [Green Version]
- Cavagnaro, T.R.; Bender, S.F.; Asghari, H.R.; van der Heijden, M.G. The role of arbuscular mycorrhizas in reducing soil nutrient loss. Trends Plant Sci. 2015, 20, 283–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosier, A.; Medeiros, F.H.V.; Bais, H.P. Defining plant growth promoting rhizobacteria molecular and biochemical networks in beneficial plant-microbe interactions. Plant Soil 2018, 428, 35–55. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Shen, C.; Sun, B.; Wang, X.N.; Wei, D.; Lyu, L.Y. Effects of drought stress on C, N and P stoichiometry of Ulmus pumila seedlings in Horqin sandy land, China. Ying Yong Sheng Tai Xue Bao J. Appl. Ecol. 2018, 29, 2286–2294, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Yuan, Z.; Chen, H.Y.; Reich, P.B. Global-scale latitudinal patterns of plant fine-root nitrogen and phosphorus. Nat. Commun. 2011, 2, 344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Guo, W.H.; Yang, X.Q.; Han, Y.Z.; Yu, M.K.; Wu, T.G. Variations in leaf C, N, P stoichiometry of Quercus acutissima provenance forests. Ying Yong Sheng Tai Xue Bao J. Appl. Ecol. 2016, 27, 2225–2230, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Hessen, D.O.; Jensen, T.C.; Kyle, M.; Elser, J.J. RNA responses to N- and P-limitation; reciprocal regulation of stoichiometry and growth rate in Brachionus. Funct. Ecol. 2007, 21, 956–962. [Google Scholar] [CrossRef]
- Yang, Y.; Tang, M.; Sulpice, R.; Chen, H.; Tian, S.; Ban, Y. Arbuscular Mycorrhizal Fungi Alter Fractal Dimension Characteristics of Robinia pseudoacacia L. Seedlings Through Regulating Plant Growth, Leaf Water Status, Photosynthesis, and Nutrient Concentration Under Drought Stress. J. Plant Growth Regul. 2014, 33, 612–620. [Google Scholar] [CrossRef]
- Hidri, R.; Mahmoud, O.M.-B.; Debez, A.; Abdelly, C.; Barea, J.-M.; Azcon, R. Modulation of C:N:P stoichiometry is involved in the effectiveness of a PGPR and AM fungus in increasing salt stress tolerance of Sulla carnosa Tunisian provenances. Appl. Soil Ecol. 2019, 143, 161–172. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, P.; Liu, X.; Xiao, L.; Shi, P.; Zhao, B. Effects of farmland conversion on the stoichiometry of carbon, nitrogen, and phosphorus in soil aggregates on the Loess Plateau of China. Geoderma 2019, 351, 188–196. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Wang, T.; Zou, B.Z.; Wang, S.R.; Huang, Z.Q.; Wan, X.H. Soil C: N: P stoichiometry and nutrient dynamics in Cunninghamia lanceolata plantations during different growth stages. Ying Yong Sheng Tai Xue Bao J. Appl. Ecol. 2020, 31, 3597–3604, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Bender, S.F.; van der Heijden, M.G. Soil biota enhance agricultural sustainability by improving crop yield, nutrient uptake and reducing nitrogen leaching losses. J. Appl. Ecol. 2014, 52, 228–239. [Google Scholar] [CrossRef]
- Storer, K.; Coggan, A.; Ineson, P.; Hodge, A. Arbuscular mycorrhizal fungi reduce nitrous oxide emissions from N2O hotspots. New Phytol. 2017, 220, 1285–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreau, D.; Bardgett, R.D.; Finlay, R.D.; Jones, D.L.; Philippot, L. A plant perspective on nitrogen cycling in the rhizosphere. Funct. Ecol. 2019, 33, 540–552. [Google Scholar] [CrossRef] [Green Version]
- Parihar, M.; Meena, V.S.; Mishra, P.K.; Rakshit, A.; Choudhary, M.; Yadav, R.P.; Rana, K.; Bisht, J.K. Arbuscular mycorrhiza: A viable strategy for soil nutrient loss reduction. Arch. Microbiol. 2019, 201, 723–735. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; He, F.; Kuzyakov, Y.; Xiao, H.; Hoang, D.T.T.; Pu, S.; Razavi, B.S. Nutrients in the rhizosphere: A meta-analysis of content, availability, and influencing factors. Sci. Total. Environ. 2022, 826, 153908. [Google Scholar] [CrossRef]
- Zhang, S.; Lehmann, A.; Zheng, W.; You, Z.; Rillig, M.C. Arbuscular mycorrhizal fungi increase grain yields: A meta-analysis. New Phytol. 2018, 222, 543–555. [Google Scholar] [CrossRef]
- Manzoni, S.; Trofymow, J.A.; Jackson, R.B.; Porporato, A. Stoichiometric controls on carbon, nitrogen, and phosphorus dynamics in decomposing litter. Ecol. Monogr. 2010, 80, 89–106. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Muledeer, T.; Tian, D.; Feng, Z.-Z. Seasonal dynamics of soil microbial biomass carbon, nitrogen and phosphorus stoichiometry across global forest ecosystems. J. Plant Ecol. 2019, 43, 532–542, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Zhang, D.; Wang, C.; Li, X.; Yang, X.; Zhao, L.; Liu, L.; Zhu, C.; Li, R. Linking plant ecological stoichiometry with soil nutrient and bacterial communities in apple orchards. Appl. Soil Ecol. 2017, 126, 1–10. [Google Scholar] [CrossRef]
- Xiao, L.; Bi, Y.; Du, S.; Wang, Y.; Guo, C.; Christie, P. Response of ecological stoichiometry and stoichiometric homeostasis in the plant-litter-soil system to re-vegetation type in arid mining subsidence areas. J. Arid. Environ. 2021, 184, 104298. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, B.-R.; An, S.-S. Ecological stoichiometry in leaves, roots, litters and soil among different plant communities in a desertified region of Northern China. Catena 2018, 166, 328–338. [Google Scholar] [CrossRef]
- Wang, J.; Wang, J.; Wang, L.; Zhang, H.; Guo, Z.; Wang, G.G.; Smith, W.K.; Wu, T. Does stoichiometric homeostasis differ among tree organs and with tree age? For. Ecol. Manag. 2019, 453, 117637. [Google Scholar] [CrossRef]
- Bi, Y.; Guo, Y.; Christie, P. Mining subsidence area reconstruction with N2-fixing plants promotes arbuscular mycorrhizal fungal biodiversity and microbial biomass C:N:P stoichiometry of cyanobacterial biocrusts. For. Ecol. Manag. 2022, 503, 119763. [Google Scholar] [CrossRef]
- Yang, G.; Yang, X.; Zhang, W.; Wei, Y.; Ge, G.; Lu, W.; Sun, J.; Liu, N.; Kan, H.; Shen, Y.; et al. Arbuscular mycorrhizal fungi affect plant community structure under various nutrient conditions and stabilize the community productivity. Oikos 2015, 125, 576–585. [Google Scholar] [CrossRef]
- Mariotte, P.; Canarini, A.; Dijkstra, F.A. Stoichiometric N:P flexibility and mycorrhizal symbiosis favour plant resistance against drought. J. Ecol. 2017, 105, 958–967. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Water Regimes | AMF Status | Plant Height (cm) | Stem Diameter (mm) | Leaf Area (cm2) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| PS | SS | REC | PS | SS | REC | PS | SS | REC | ||
| DS | CK | 47.94 ± 2.26 c | 51.32 ± 2.62 d | 59.58 ± 1.82 f | 6.76 ± 0.19 d | 7.51 ± 0.30 e | 7.86 ± 0.22 d | 17.50 ± 1.33 a | 18.05 ± 1.25 b | 19.93 ± 1.53 c |
| C. etunicatum | 93.04 ± 3.21 a | 98.86 ± 3.10 ab | 103.54 ± 2.33 b | 10.88 ± 0.33 a | 11.89 ± 0.10 ab | 12.17 ± 0.36 a | 22.37 ± 2.57 a | 23.99 ± 2.68 ab | 26.47 ± 2.43 ab | |
| F. mosseae | 82.18 ± 4.83 ab | 88.50 ± 4.73 c | 95.26 ± 3.17 c | 10.01 ± 0.34 ab | 10.92 ± 0.33 ab | 11.59 ± 0.37 ab | 19.10 ± 1.30 a | 20.28 ± 1.63 ab | 22.53 ± 1.53 bc | |
| Co-inoculation | 74.66 ± 2.86 b | 82.42 ± 3.65 c | 87.46 ± 2.27 d | 8.69 ± 0.62 bc | 9.25 ± 0.60 cd | 10.45 ± 0.47 c | 21.74 ± 2.67 a | 22.94 ± 2.68 ab | 24.95 ± 2.10 abc | |
| WW | CK | 45.22 ± 2.09 c | 55.42 ± 1.33 d | 69.92 ± 2.60 e | 7.16 ± 0.14 cd | 7.84 ± 0.12 de | 8.53 ± 0.21 d | 17.99 ± 1.15 a | 18.60 ± 1.06 b | 19.84 ± 1.17 c |
| C. etunicatum | 93.34 ± 3.15 a | 103.82 ± 3.32 a | 112.50 ± 1.23 a | 11.02 ± 0.34 a | 12.12 ± 0.05 a | 12.26 ± 0.14 a | 22.50 ± 2.54 a | 26.21 ± 1.82 a | 30.10 ± 1.07 a | |
| F. mosseae | 84.98 ± 4.69 ab | 92.46 ± 4.53 bc | 103.62 ± 2.22 b | 10.29 ± 0.37 ab | 11.41 ± 0.17 ab | 12.10 ± 0.23 ab | 19.79 ± 0.74 a | 21.93 ± 0.93 ab | 24.93 ± 0.60 abc | |
| Co-inoculation | 77.08 ± 2.46 b | 87.48 ± 2.63 c | 100.70 ± 1.41 bc | 8.85 ± 0.60 bc | 10.49 ± 0.43 bc | 11.22 ± 0.38 bc | 22.30 ± 2.64 a | 25.44 ± 2.36 a | 28.52 ± 2.27 a | |
| Significance | ||||||||||
| AMF | ** | ** | ** | ** | ** | ** | ns | * | ** | |
| DS | ns | ns | ** | ns | * | * | ns | ns | ns | |
| AMF×DS | ns | ns | ns | ns | ns | ns | ns | ns | ns | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, X.; Liao, X.; Yan, Q.; Xie, Y.; Chen, J.; Liang, G.; Chen, M.; Xiao, S.; Chen, Y.; Liu, J. Arbuscular Mycorrhizal Fungi Improve the Growth, Water Status, and Nutrient Uptake of Cinnamomum migao and the Soil Nutrient Stoichiometry under Drought Stress and Recovery. J. Fungi 2023, 9, 321. https://doi.org/10.3390/jof9030321
Xiao X, Liao X, Yan Q, Xie Y, Chen J, Liang G, Chen M, Xiao S, Chen Y, Liu J. Arbuscular Mycorrhizal Fungi Improve the Growth, Water Status, and Nutrient Uptake of Cinnamomum migao and the Soil Nutrient Stoichiometry under Drought Stress and Recovery. Journal of Fungi. 2023; 9(3):321. https://doi.org/10.3390/jof9030321
Chicago/Turabian StyleXiao, Xuefeng, Xiaofeng Liao, Qiuxiao Yan, Yuangui Xie, Jingzhong Chen, Gelin Liang, Meng Chen, Shengyang Xiao, Yuan Chen, and Jiming Liu. 2023. "Arbuscular Mycorrhizal Fungi Improve the Growth, Water Status, and Nutrient Uptake of Cinnamomum migao and the Soil Nutrient Stoichiometry under Drought Stress and Recovery" Journal of Fungi 9, no. 3: 321. https://doi.org/10.3390/jof9030321
APA StyleXiao, X., Liao, X., Yan, Q., Xie, Y., Chen, J., Liang, G., Chen, M., Xiao, S., Chen, Y., & Liu, J. (2023). Arbuscular Mycorrhizal Fungi Improve the Growth, Water Status, and Nutrient Uptake of Cinnamomum migao and the Soil Nutrient Stoichiometry under Drought Stress and Recovery. Journal of Fungi, 9(3), 321. https://doi.org/10.3390/jof9030321