
Characterization of Virulence Factors in Candida Species Causing Candidemia in a Tertiary Care Hospital in Bangkok, Thailand

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Isolates and Ethical Statement
2.2. Hydrolytic Enzyme Activity Assays
2.3. CSH Assay
2.4. Biofilm Formation
2.5. Candida spp. Invasion Assay
2.6. Cytotoxicity Assay
2.7. Fluorescent Staining and Confocal Microscopy
2.8. Human Monocyte Uptake of Candida Cells
2.9. Amplification and Sequencing of ERG11 Gene
2.10. Statistical Analysis
3. Results
3.1. Virulence Assessment of Invasive Candida spp. Isolates
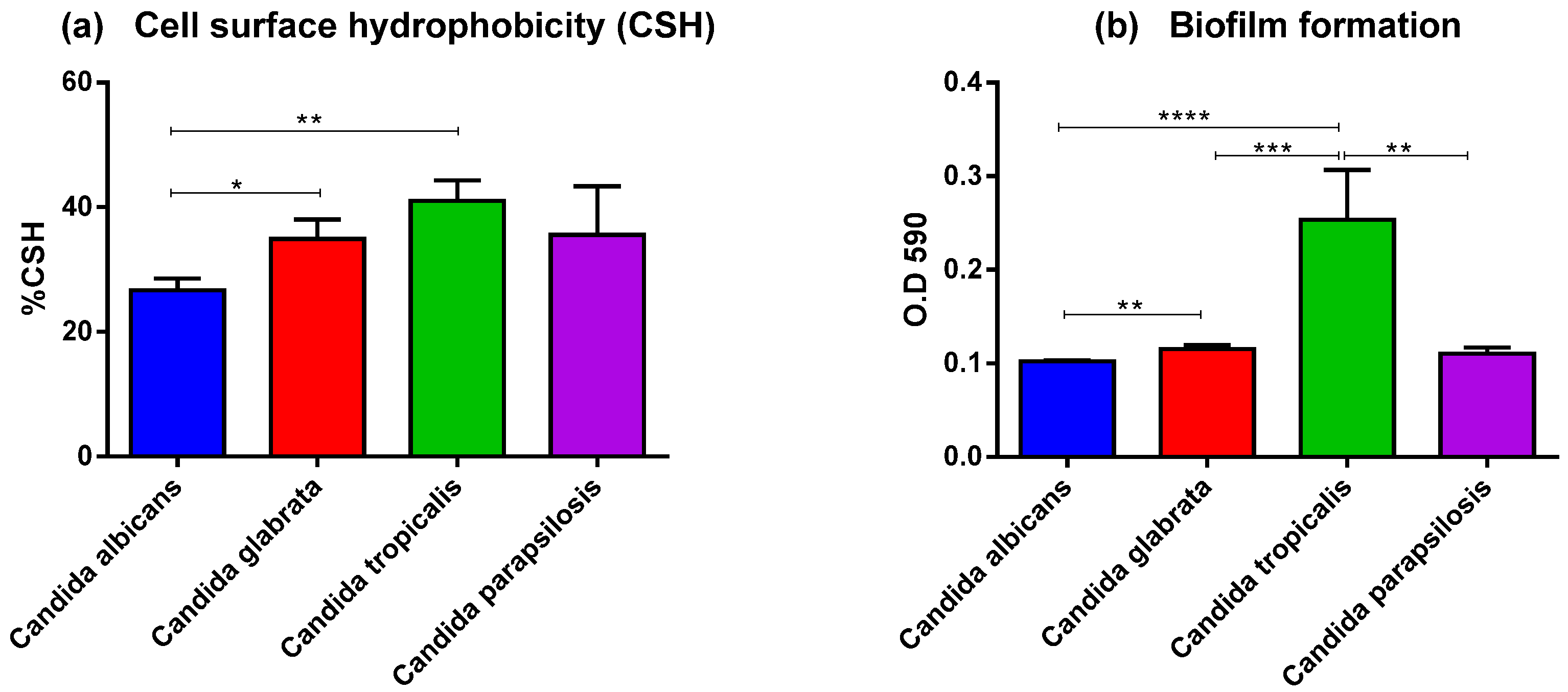
3.2. Adhesion of Invasive Candida spp. Isolates
3.3. Ability to Invade and Damage Epithelial Cells
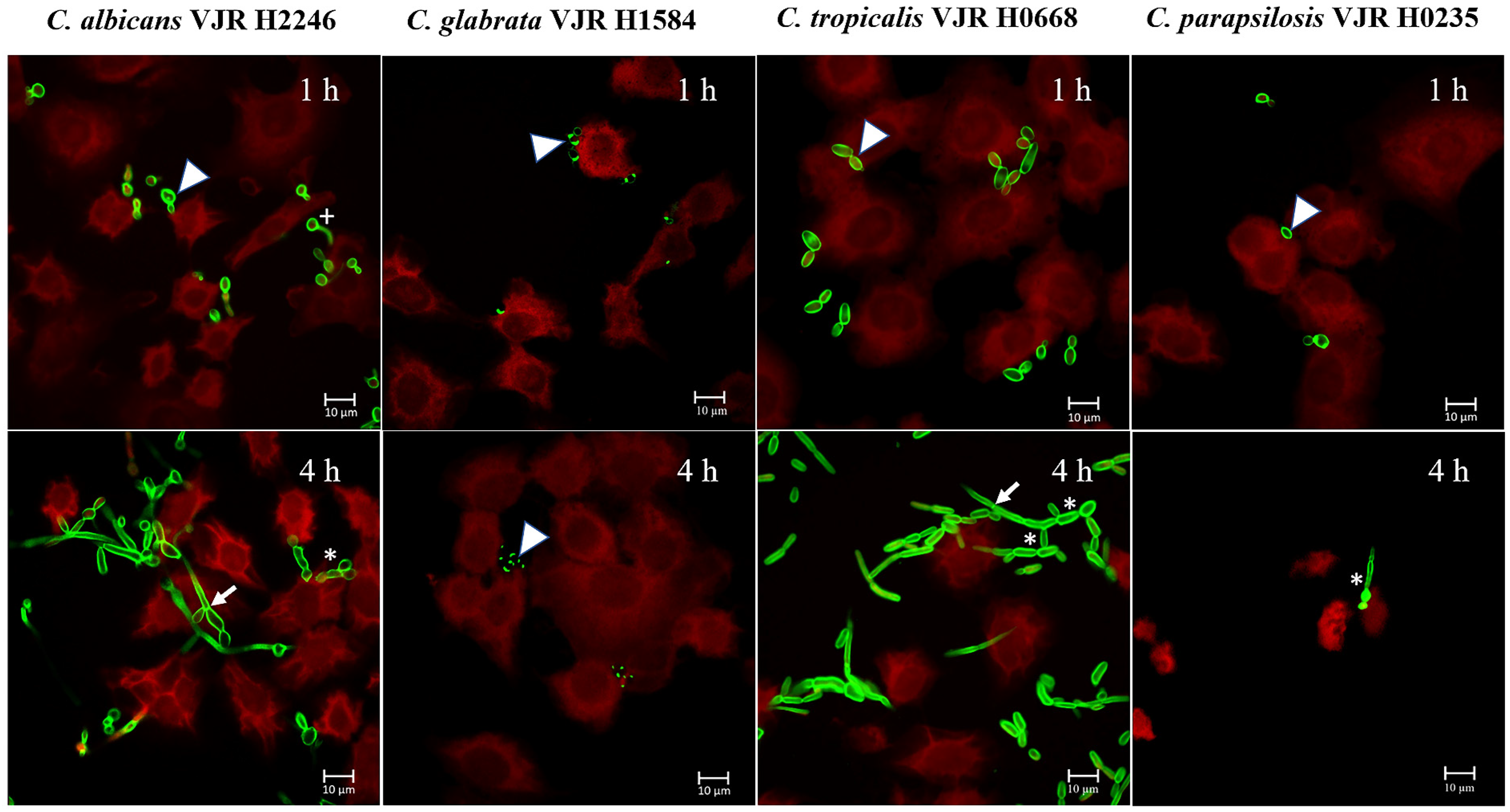
3.4. Morphologic Transition in Candida spp. Isolates
3.5. Phagocytosis Assay with Candida spp. Isolates
3.6. Mutation in ERG11 Gene
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kullberg, B.J.; Arendrup, M.C. Invasive candidiasis. N. Engl. J. Med. 2015, 373, 1445–1456. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Du, M.; Chen, L.A.; Liu, Y.; Liang, Z. Nosocomial bloodstream infection due to Candida spp. in China: Species distribution, clinical features, and outcomes. Mycopathologia 2016, 181, 485–495. [Google Scholar] [CrossRef]
- Kothalawala, M.; Jayaweera, J.; Arunan, S.; Jayathilake, A. The emergence of non-albicans candidemia and evaluation of HiChrome Candida differential agar and VITEK2 YST(R) platform for differentiation of Candida bloodstream isolates in teaching hospital Kandy, Sri Lanka. BMC Microbiol. 2019, 19, 136. [Google Scholar] [CrossRef]
- Xiao, Z.; Wang, Q.; Zhu, F.; An, Y. Epidemiology, species distribution, antifungal susceptibility and mortality risk factors of candidemia among critically ill patients: A retrospective study from 2011 to 2017 in a teaching hospital in China. Antimicrob. Resist. Infect. Control. 2019, 8, 89. [Google Scholar] [CrossRef]
- Pfaller, M.; Neofytos, D.; Diekema, D.; Azie, N.; Meier-Kriesche, H.U.; Quan, S.P.; Horn, D. Epidemiology and outcomes of candidemia in 3648 patients: Data from the prospective antifungal therapy (PATH Alliance(R)) registry, 2004–2008. Diagn Microbiol. Infect. Dis. 2012, 74, 323–331. [Google Scholar] [CrossRef]
- Boonsilp, S.; Homkaew, A.; Phumisantiphong, U.; Nutalai, D.; Wongsuk, T. Species distribution, antifungal susceptibility, and molecular epidemiology of Candida species causing candidemia in a tertiary care hospital in Bangkok, Thailand. J. Fungi 2021, 577. [Google Scholar] [CrossRef]
- Tulyaprawat, O.; Pharkjaksu, S.; Chongtrakool, P.; Ngamskulrungroj, P. An Association of an eBURST group with triazole resistance of Candida tropicalis blood isolates. Front. Microbiol. 2020, 11, 934. [Google Scholar] [CrossRef]
- Doi, A.M.; Pignatari, A.C.; Edmond, M.B.; Marra, A.R.; Camargo, L.F.; Siqueira, R.A.; da Mota, V.P.; Colombo, A.L. Epidemiology and microbiologic characterization of nosocomial candidemia from a Brazilian national surveillance program. PLoS ONE 2016, 11, e0146909. [Google Scholar] [CrossRef]
- Khairat, S.M.; Sayed, A.M.; Nabih, M.; Soliman, N.S.; Hassan, Y.M. Prevalence of Candida blood stream infections among children in tertiary care hospital: Detection of species and antifungal susceptibility. Infect. Drug Resist. 2019, 12, 2409–2416. [Google Scholar] [CrossRef] [Green Version]
- Whaley, S.G.; Berkow, E.L.; Rybak, J.M.; Nishimoto, A.T.; Barker, K.S.; Rogers, P.D. Azole antifungal resistance in Candida albicans and emerging non-albicans Candida Species. Front. Microbiol. 2016, 7, 2173. [Google Scholar] [CrossRef] [Green Version]
- Zuza-Alves, D.L.; Silva-Rocha, W.P.; Chaves, G.M. An Update on Candida tropicalis based on basic and clinical approaches. Front. Microbiol. 2017, 8, 1927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, L.; Cao, Z.; Wang, Q.; Wang, Y.; Wang, X.; Chen, H.; Wang, H. MDR1 overexpression combined with ERG11 mutations induce high-level fluconazole resistance in Candida tropicalis clinical isolates. BMC Infect. Dis. 2018, 18, 162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Negri, M.; Silva, S.; Henriques, M.; Oliveira, R. Insights into Candida tropicalis nosocomial infections and virulence factors. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 1399–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mora Carpio, A.L.; Climaco, A. Fungemia Candidiasis. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Chin, V.K.; Foong, K.J.; Maha, A.; Rusliza, B.; Norhafizah, M.; Ng, K.P.; Chong, P.P. Candida albicans isolates from a Malaysian hospital exhibit more potent phospholipase and haemolysin activities than non-albicans Candida isolates. Trop. Biomed. 2013, 30, 654–662. [Google Scholar]
- Mattei, A.S.; Alves, S.H.; Severo, C.B.; Guazzelli Lda, S.; Oliveira Fde, M.; Severo, L.C. Determination of germ tube, phospholipase, and proteinase production by bloodstream isolates of Candida albicans. Rev. Soc. Bras. Med. Trop. 2013, 46, 340–342. [Google Scholar] [CrossRef] [Green Version]
- Neji, S.; Hadrich, I.; Trabelsi, H.; Abbes, S.; Cheikhrouhou, F.; Sellami, H.; Makni, F.; Ayadi, A. Virulence factors, antifungal susceptibility and molecular mechanisms of azole resistance among Candida parapsilosis complex isolates recovered from clinical specimens. J. Biomed. Sci. 2017, 24, 67. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarti, A.; Nayak, N.; Talwar, P. In vitro proteinase production by Candida species. Mycopathologia 1991, 114, 163–168. [Google Scholar] [CrossRef]
- Tsang, C.S.P.; Chu, F.C.S.; Leung, W.K.; Jin, L.J.; Samaranayake, L.P.; Siu, S.C. Phospholipase, proteinase and haemolytic activities of Candida albicans isolated from oral cavities of patients with type 2 diabetes mellitus. J. Med. Microbiol. 2007, 56 Pt 10, 1393–1398. [Google Scholar] [CrossRef] [Green Version]
- Luo, G.; Samaranayake, L.P.; Yau, J.Y. Candida species exhibit differential in vitro hemolytic activities. J. Clin. Microbiol. 2001, 39, 2971–2974. [Google Scholar] [CrossRef] [Green Version]
- Sachin, C.R.K.; Santosh, S. In vitro evaluation of proteinase, phospholipase and haemolysin activities of Candida species isolated from clinical specimens. Int. J. Med. Biomed. Res. 2012, 1, 153–157. [Google Scholar] [CrossRef]
- Rosenberg, M. Bacterial adherence to hydrocarbon: A useful technique for studying cell surface hydrophobicity. FEMS Microbiol. Lett. 1984, 22, 289–295. [Google Scholar] [CrossRef]
- Ramage, G.; Vande Walle, K.; Wickes, B.L.; Lopez-Ribot, J.L. Standardized method for in vitro antifungal susceptibility testing of Candida albicans biofilms. Antimicrob. Agents Chemother. 2001, 45, 2475–2479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correia, I.; Prieto, D.; Roman, E.; Wilson, D.; Hube, B.; Alonso-Monge, R.; Pla, J. Cooperative role of MAPK pathways in the interaction of Candida albicans with the host epithelium. Microorganisms 2019, 8, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saiprom, N.; Sangsri, T.; Tandhavanant, S.; Sengyee, S.; Phunpang, R.; Preechanukul, A.; Surin, U.; Tuanyok, A.; Lertmemongkolchai, G.; Chantratita, W.; et al. Genomic loss in environmental and isogenic morphotype isolates of Burkholderia pseudomallei is associated with intracellular survival and plaque-forming efficiency. PLoS Negl. Trop. Dis. 2020, 14, e0008590. [Google Scholar] [CrossRef]
- Dagher, Z.; Xu, S.; Negoro, P.E.; Khan, N.S.; Feldman, M.B.; Reedy, J.L.; Tam, J.M.; Sykes, D.B.; Mansour, M.K. Fluorescent tracking of yeast division clarifies the essential role of spleen tyrosine kinase in the intracellular control of Candida glabrata in macrophages. Front. Immunol. 2018, 9, 1058. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, S.; Yamabe, M.; Yamaguchi, Y.; Kobayashi, Y.; Konno, T.; Tada, K. Establishment and characterization of a human acute monocytic leukemia cell line (THP-1). Int. J. Cancer 1980, 26, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Villasenor-Cardoso, M.I.; Frausto-Del-Rio, D.A.; Ortega, E. Aminopeptidase N (CD13) is involved in phagocytic processes in human dendritic cells and macrophages. Biomed. Res. Int. 2013, 2013, 562984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, S.; Kannan, I.; Mohanram, K. Extensive ERG11 mutations associated with fluconazole-resistant Candida albicans isolated from HIV-infected patients. Curr. Med. Mycol. 2019, 5, 1–6. [Google Scholar] [CrossRef]
- Ngamchokwathana, C.; Chongtrakool, P.; Waesamaae, A.; Chayakulkeeree, M. Risk factors and outcomes of non-albicans Candida bloodstream infection in patients with candidemia at Siriraj Hospital-Thailand’s largest national tertiary referral hospital. J. Fungi 2021, 7, 269. [Google Scholar] [CrossRef]
- Ying, Y.; Zhao, Y.; Hu, X.; Cai, Z.; Liu, X.; Jin, G.; Zhang, J.; Zhang, J.; Liu, J.; Huang, X. In vitro fluconazole susceptibility of 1,903 clinical isolates of Candida albicans and the identification of ERG11 mutations. Microb. Drug Resist. 2013, 19, 266–273. [Google Scholar] [CrossRef]
- Morio, F.; Loge, C.; Besse, B.; Hennequin, C.; Le Pape, P. Screening for amino acid substitutions in the Candida albicans Erg11 protein of azole-susceptible and azole-resistant clinical isolates: New substitutions and a review of the literature. Diagn. Microbiol. Infect. Dis. 2010, 66, 373–384. [Google Scholar] [CrossRef]
- Jiang, C.; Dong, D.; Yu, B.; Cai, G.; Wang, X.; Ji, Y.; Peng, Y. Mechanisms of azole resistance in 52 clinical isolates of Candida tropicalis in China. J. Antimicrob. Chemother. 2013, 68, 778–785. [Google Scholar] [CrossRef] [PubMed]
- Chew, K.L.; Cheng, J.W.; Jureen, R.; Lin, R.T.; Teo, J.W. ERG11 mutations are associated with high-level azole resistance in clinical Candida tropicalis isolates, a Singapore study. Mycoscience 2017, 58, 111–115. [Google Scholar] [CrossRef]
- Paul, S.; Shaw, D.; Joshi, H.; Singh, S.; Chakrabarti, A.; Rudramurthy, S.M.; Ghosh, A.K. Mechanisms of azole antifungal resistance in clinical isolates of Candida tropicalis. PLoS ONE 2022, 17, e0269721. [Google Scholar] [CrossRef] [PubMed]
- Forastiero, A.; Mesa-Arango, A.C.; Alastruey-Izquierdo, A.; Alcazar-Fuoli, L.; Bernal-Martinez, L.; Pelaez, T.; Lopez, J.F.; Grimalt, J.O.; Gomez-Lopez, A.; Cuesta, I.; et al. Candida tropicalis antifungal cross-resistance is related to different azole target (Erg11p) modifications. Antimicrob. Agents Chemother. 2013, 57, 4769–4781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, J.; Zhang, J.; Chen, W.; Sun, Y.; Wan, Z.; Li, R.; Liu, W. The A395T mutation in ERG11 gene confers fluconazole resistance in Candida tropicalis causing candidemia. Mycopathologia 2015, 179, 213–218. [Google Scholar] [CrossRef]
- Klotz, S.A.; Drutz, D.J.; Zajic, J.E. Factors governing adherence of Candida species to plastic surfaces. Infect. Immun. 1985, 50, 97–101. [Google Scholar] [CrossRef]
- Panagoda, G.J.; Ellepola, A.N.; Samaranayake, L.P. Adhesion of Candida parapsilosis to epithelial and acrylic surfaces correlates with cell surface hydrophobicity. Mycoses 2001, 44, 29–35. [Google Scholar] [CrossRef]
- Galan-Ladero, M.A.; Blanco-Blanco, M.T.; Hurtado, C.; Perez-Giraldo, C.; Blanco, M.T.; Gomez-Garcia, A.C. Determination of biofilm production by Candida tropicalis isolated from hospitalized patients and its relation to cellular surface hydrophobicity, plastic adherence and filamentation ability. Yeast 2013, 30, 331–339. [Google Scholar] [CrossRef]
- Dabiri, S.; Shams-Ghahfarokhi, M.; Razzaghi-Abyaneh, M. Comparative analysis of proteinase, phospholipase, hydrophobicity and biofilm forming ability in Candida species isolated from clinical specimens. J. Mycol. Med. 2018, 28, 437–442. [Google Scholar] [CrossRef]
- Silva-Dias, A.; Miranda, I.M.; Branco, J.; Monteiro-Soares, M.; Pina-Vaz, C.; Rodrigues, A.G. Adhesion, biofilm formation, cell surface hydrophobicity, and antifungal planktonic susceptibility: Relationship among Candida spp. Front. Microbiol. 2015, 6, 205. [Google Scholar] [CrossRef] [Green Version]
- Cavalheiro, M.; Teixeira, M.C. Candida biofilms: Threats, challenges, and promising strategies. Front. Med. 2018, 5, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sriphannam, C.; Nuanmuang, N.; Saengsawang, K.; Amornthipayawong, D.; Kummasook, A. Anti-fungal susceptibility and virulence factors of Candida spp. isolated from blood cultures. J. Mycol. Med. 2019, 29, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Nouraei, H.; Pakshir, K.; ZareShahrabadi, Z.; Zomorodian, K. High detection of virulence factors by Candida species isolated from bloodstream of patients with candidemia. Microb. Pathog. 2020, 149, 104574. [Google Scholar] [CrossRef] [PubMed]
- Moyes, D.L.; Richardson, J.P.; Naglik, J.R. Candida albicans-epithelial interactions and pathogenicity mechanisms: Scratching the surface. Virulence 2015, 6, 338–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaller, M.; Borelli, C.; Korting, H.C.; Hube, B. Hydrolytic enzymes as virulence factors of Candida albicans. Mycoses 2005, 48, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Linares, C.E.; de Loreto, E.S.; Silveira, C.P.; Pozzatti, P.; Scheid, L.A.; Santurio, J.M.; Alves, S.H. Enzymatic and hemolytic activities of Candida dubliniensis strains. Rev. Inst. Med. Trop. Sao Paulo 2007, 49, 203–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furlaneto, M.C.; Favero, D.; Franca, E.J.; Furlaneto-Maia, L. Effects of human blood red cells on the haemolytic capability of clinical isolates of Candida tropicalis. J. Biomed. Sci. 2015, 22, 13. [Google Scholar] [CrossRef] [Green Version]
- Vieira de Melo, A.P.; Zuza-Alves, D.L.; da Silva-Rocha, W.P.; Ferreira Canario de Souza, L.B.; Francisco, E.C.; Salles de Azevedo Melo, A.; Maranhao Chaves, G. Virulence factors of Candida spp. obtained from blood cultures of patients with candidemia attended at tertiary hospitals in Northeast Brazil. J. Mycol. Med. 2019, 29, 132–139. [Google Scholar] [CrossRef]
- Ghannoum, M.A. Potential role of phospholipases in virulence and fungal pathogenesis. Clin. Microbiol. Rev. 2000, 13, 122–143, table of contents. [Google Scholar] [CrossRef]
- Canela, H.M.S.; Cardoso, B.; Vitali, L.H.; Coelho, H.C.; Martinez, R.; Ferreira, M. Prevalence, virulence factors and antifungal susceptibility of Candida spp. isolated from bloodstream infections in a tertiary care hospital in Brazil. Mycoses 2018, 61, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Sharma, Y.; Chumber, S.K.; Kaur, M. Studying the prevalence, species distribution, and detection of in vitro production of phospholipase from Candida isolated from cases of invasive candidiasis. J. Glob. Infect. Dis. 2017, 9, 8–11. [Google Scholar] [CrossRef]
- Borst, A.; Fluit, A.C. High levels of hydrolytic enzymes secreted by Candida albicans isolates involved in respiratory infections. J. Med. Microbiol. 2003, 52 Pt 11, 971–974. [Google Scholar] [CrossRef] [Green Version]
- Naglik, J.R.; Challacombe, S.J.; Hube, B. Candida albicans secreted aspartyl proteinases in virulence and pathogenesis. Microbiol. Mol. Biol. Rev. 2003, 67, 400–428. [Google Scholar] [CrossRef] [Green Version]
- Silva, S.; Hooper, S.J.; Henriques, M.; Oliveira, R.; Azeredo, J.; Williams, D.W. The role of secreted aspartyl proteinases in Candida tropicalis invasion and damage of oral mucosa. Clin. Microbiol. Infect. 2011, 17, 264–272. [Google Scholar] [CrossRef] [Green Version]
- Tay, S.T.; Abidin, I.A.; Hassan, H.; Ng, K.P. Proteinase, phospholipase, biofilm forming abilities and antifungal susceptibilities of Malaysian Candida isolates from blood cultures. Med. Mycol. 2011, 49, 556–560. [Google Scholar]
- Sariguzel, F.M.; Berk, E.; Koc, A.N.; Sav, H.; Demir, G. Investigation of the relationship between virulence factors and genotype of Candida spp. isolated from blood cultures. J. Infect. Dev. Ctries 2015, 9, 857–864. [Google Scholar] [CrossRef] [PubMed]
- Dalle, F.; Wachtler, B.; L’Ollivier, C.; Holland, G.; Bannert, N.; Wilson, D.; Labruere, C.; Bonnin, A.; Hube, B. Cellular interactions of Candida albicans with human oral epithelial cells and enterocytes. Cell Microbiol. 2010, 12, 248–271. [Google Scholar] [CrossRef] [PubMed]
- Wachtler, B.; Citiulo, F.; Jablonowski, N.; Forster, S.; Dalle, F.; Schaller, M.; Wilson, D.; Hube, B. Candida albicans-epithelial interactions: Dissecting the roles of active penetration, induced endocytosis and host factors on the infection process. PLoS ONE 2012, 7, e36952. [Google Scholar] [CrossRef] [Green Version]
- Silva, S.; Henriques, M.; Oliveira, R.; Azeredo, J.; Malic, S.; Hooper, S.J.; Williams, D.W. Characterization of Candida parapsilosis infection of an in vitro reconstituted human oral epithelium. Eur. J. Oral Sci. 2009, 117, 669–675. [Google Scholar] [CrossRef]
- Zupancic, M.L.; Frieman, M.; Smith, D.; Alvarez, R.A.; Cummings, R.D.; Cormack, B.P. Glycan microarray analysis of Candida glabrata adhesin ligand specificity. Mol. Microbiol. 2008, 68, 547–559. [Google Scholar] [CrossRef] [PubMed]
- De Groot, P.W.; Kraneveld, E.A.; Yin, Q.Y.; Dekker, H.L.; Gross, U.; Crielaard, W.; de Koster, C.G.; Bader, O.; Klis, F.M.; Weig, M. The cell wall of the human pathogen Candida glabrata: Differential incorporation of novel adhesin-like wall proteins. Eukaryot. Cell 2008, 7, 1951–1964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Netea, M.G.; Brown, G.D.; Kullberg, B.J.; Gow, N.A. An integrated model of the recognition of Candida albicans by the innate immune system. Nat. Rev. Microbiol. 2008, 6, 67–78. [Google Scholar] [CrossRef]
- Heilmann, C.J.; Sorgo, A.G.; Siliakus, A.R.; Dekker, H.L.; Brul, S.; de Koster, C.G.; de Koning, L.J.; Klis, F.M. Hyphal induction in the human fungal pathogen Candida albicans reveals a characteristic wall protein profile. Microbiology 2011, 157 Pt 8, 2297–2307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebanks, R.O.; Chisholm, K.; McKinnon, S.; Whiteway, M.; Pinto, D.M. Proteomic analysis of Candida albicans yeast and hyphal cell wall and associated proteins. Proteomics 2006, 6, 2147–2156. [Google Scholar] [CrossRef] [PubMed]
- Gantner, B.N.; Simmons, R.M.; Underhill, D.M. Dectin-1 mediates macrophage recognition of Candida albicans yeast but not filaments. EMBO J. 2005, 24, 1277–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borg-von Zepelin, M.; Meyer, I.; Thomssen, R.; Wurzner, R.; Sanglard, D.; Telenti, A.; Monod, M. HIV-protease inhibitors reduce cell adherence of Candida albicans strains by inhibition of yeast secreted aspartic proteases. J. Investig. Dermatol. 1999, 113, 747–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bektic, J.; Lell, C.P.; Fuchs, A.; Stoiber, H.; Speth, C.; Lass-Florl, C.; Borg-von Zepelin, M.; Dierich, M.P.; Wurzner, R. HIV protease inhibitors attenuate adherence of Candida albicans to epithelial cells in vitro. FEMS Immunol. Med. Microbiol. 2001, 31, 65–71. [Google Scholar] [CrossRef]
- Tsang, C.S.; Hong, I. HIV protease inhibitors differentially inhibit adhesion of Candida albicans to acrylic surfaces. Mycoses 2010, 53, 488–494. [Google Scholar] [CrossRef]
- De Bernardis, F.; Mondello, F.; Scaravelli, G.; Pachi, A.; Girolamo, A.; Agatensi, L.; Cassone, A. High aspartyl proteinase production and vaginitis in human immunodeficiency virus-infected women. J. Clin. Microbiol. 1999, 37, 1376–1380. [Google Scholar] [CrossRef] [Green Version]
- Cassone, A.; De Bernardis, F.; Torosantucci, A.; Tacconelli, E.; Tumbarello, M.; Cauda, R. In vitro and in vivo anticandidal activity of human immunodeficiency virus protease inhibitors. J. Infect. Dis. 1999, 180, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Cassone, A.; Tacconelli, E.; De Bernardis, F.; Tumbarello, M.; Torosantucci, A.; Chiani, P.; Cauda, R. Antiretroviral therapy with protease inhibitors has an early, immune reconstitution-independent beneficial effect on Candida virulence and oral candidiasis in human immunodeficiency virus-infected subjects. J. Infect. Dis. 2002, 185, 188–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cauda, R.; Tacconelli, E.; Tumbarello, M.; Morace, G.; De Bernardis, F.; Torosantucci, A.; Cassone, A. Role of protease inhibitors in preventing recurrent oral candidosis in patients with HIV infection: A prospective case-control study. J. Acquir. Immune. Defic. Syndr. 1999, 21, 20–25. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saiprom, N.; Wongsuk, T.; Oonanant, W.; Sukphopetch, P.; Chantratita, N.; Boonsilp, S. Characterization of Virulence Factors in Candida Species Causing Candidemia in a Tertiary Care Hospital in Bangkok, Thailand. J. Fungi 2023, 9, 353. https://doi.org/10.3390/jof9030353
Saiprom N, Wongsuk T, Oonanant W, Sukphopetch P, Chantratita N, Boonsilp S. Characterization of Virulence Factors in Candida Species Causing Candidemia in a Tertiary Care Hospital in Bangkok, Thailand. Journal of Fungi. 2023; 9(3):353. https://doi.org/10.3390/jof9030353
Chicago/Turabian StyleSaiprom, Natnaree, Thanwa Wongsuk, Worrapoj Oonanant, Passanesh Sukphopetch, Narisara Chantratita, and Siriphan Boonsilp. 2023. "Characterization of Virulence Factors in Candida Species Causing Candidemia in a Tertiary Care Hospital in Bangkok, Thailand" Journal of Fungi 9, no. 3: 353. https://doi.org/10.3390/jof9030353
APA StyleSaiprom, N., Wongsuk, T., Oonanant, W., Sukphopetch, P., Chantratita, N., & Boonsilp, S. (2023). Characterization of Virulence Factors in Candida Species Causing Candidemia in a Tertiary Care Hospital in Bangkok, Thailand. Journal of Fungi, 9(3), 353. https://doi.org/10.3390/jof9030353