
Virulence and Genetic Types of Blumeria graminis f. sp. hordei in Tibet and Surrounding Areas


Abstract
:1. Introduction
2. Materials and Methods
2.1. Lines and Varieties of Barley
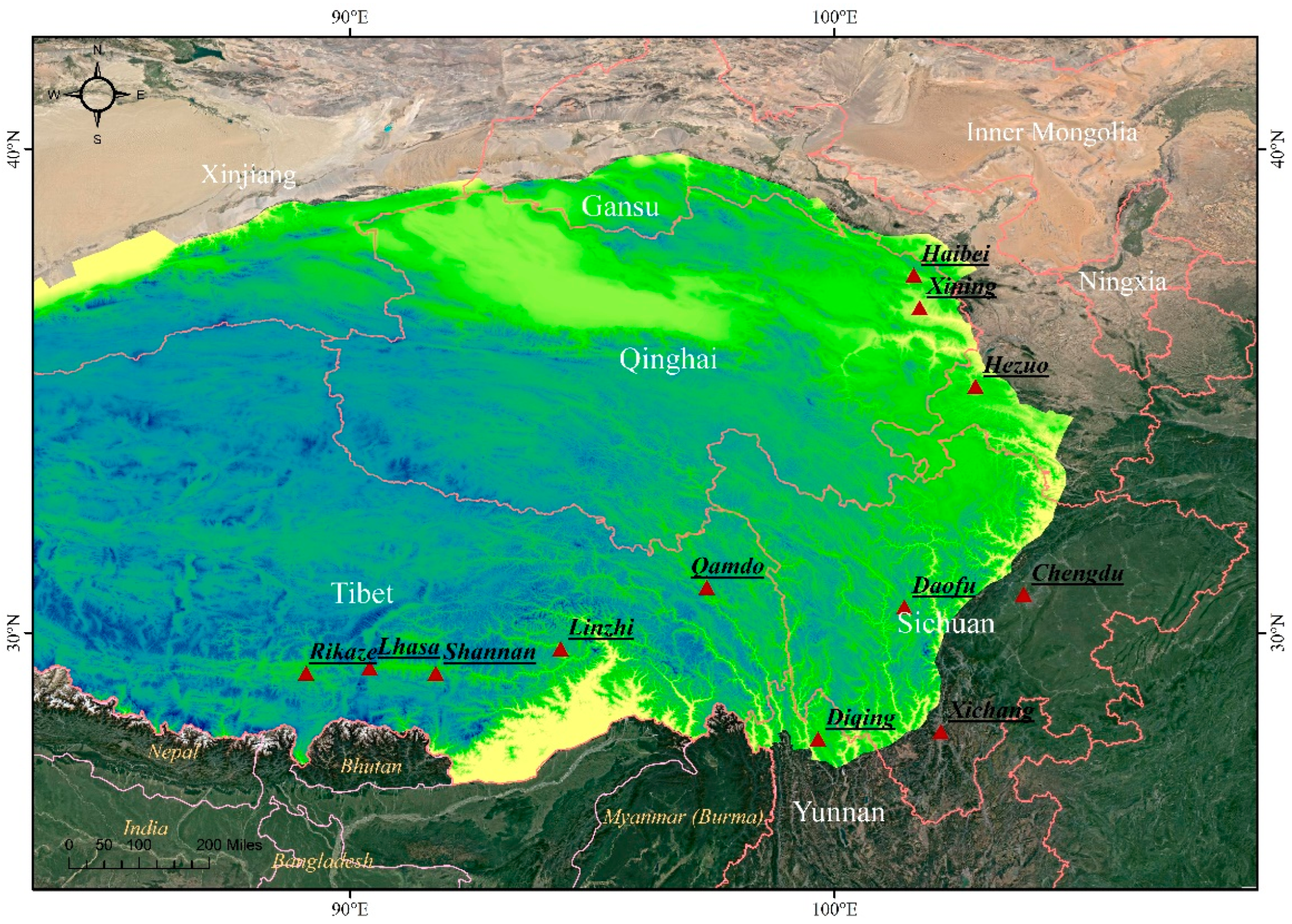
2.2. Disease Samples and Bgh Isolates
2.3. Spore Preparation
2.4. Virulence Typing
2.5. DNA Extraction and PCR Amplification
2.6. Data Processing
3. Results
3.1. Virulence Diversity of Bgh Populations
3.1.1. Virulence Frequencies of Bgh
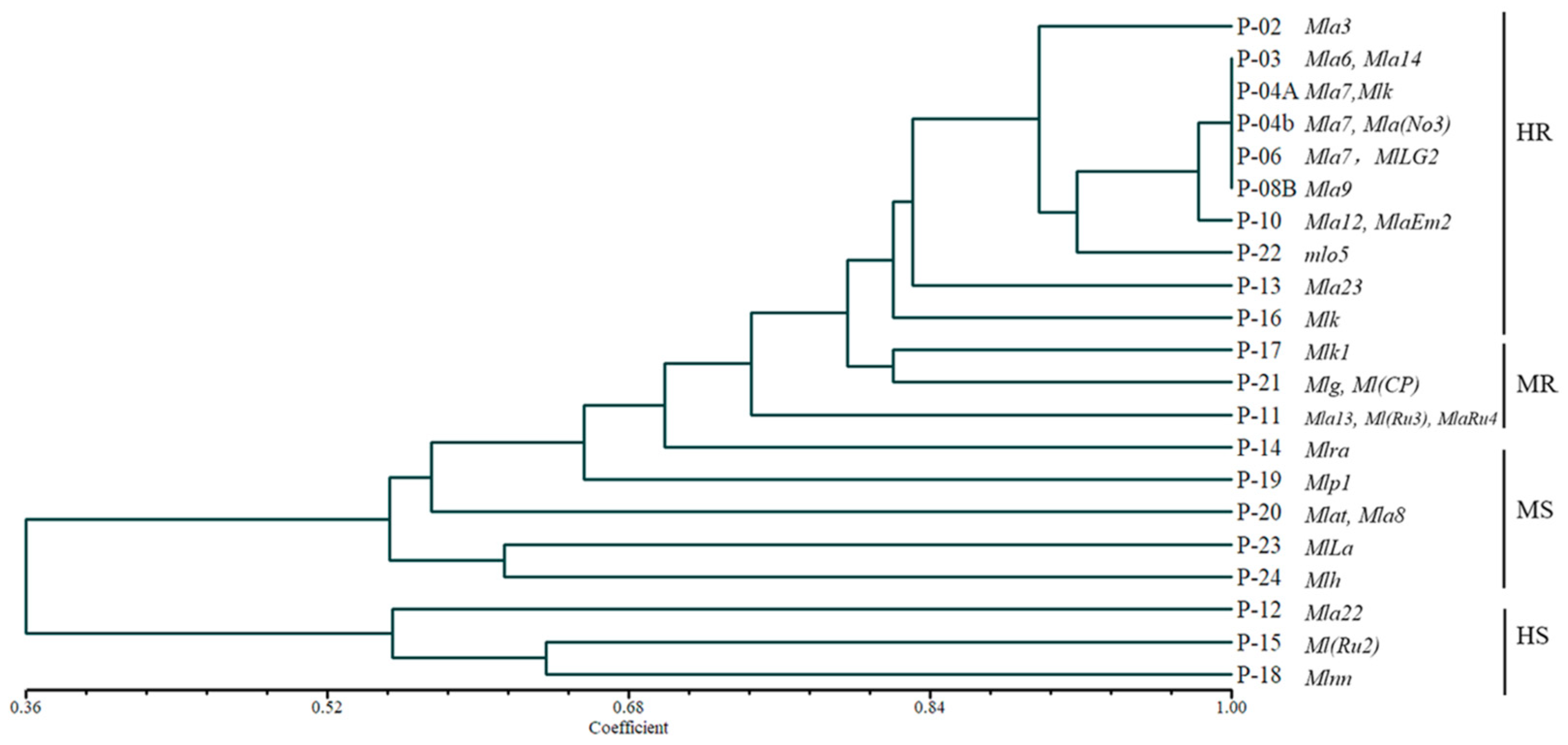
3.1.2. Virulence Types of Bgh
3.1.3. Virulence Diversity of Bgh
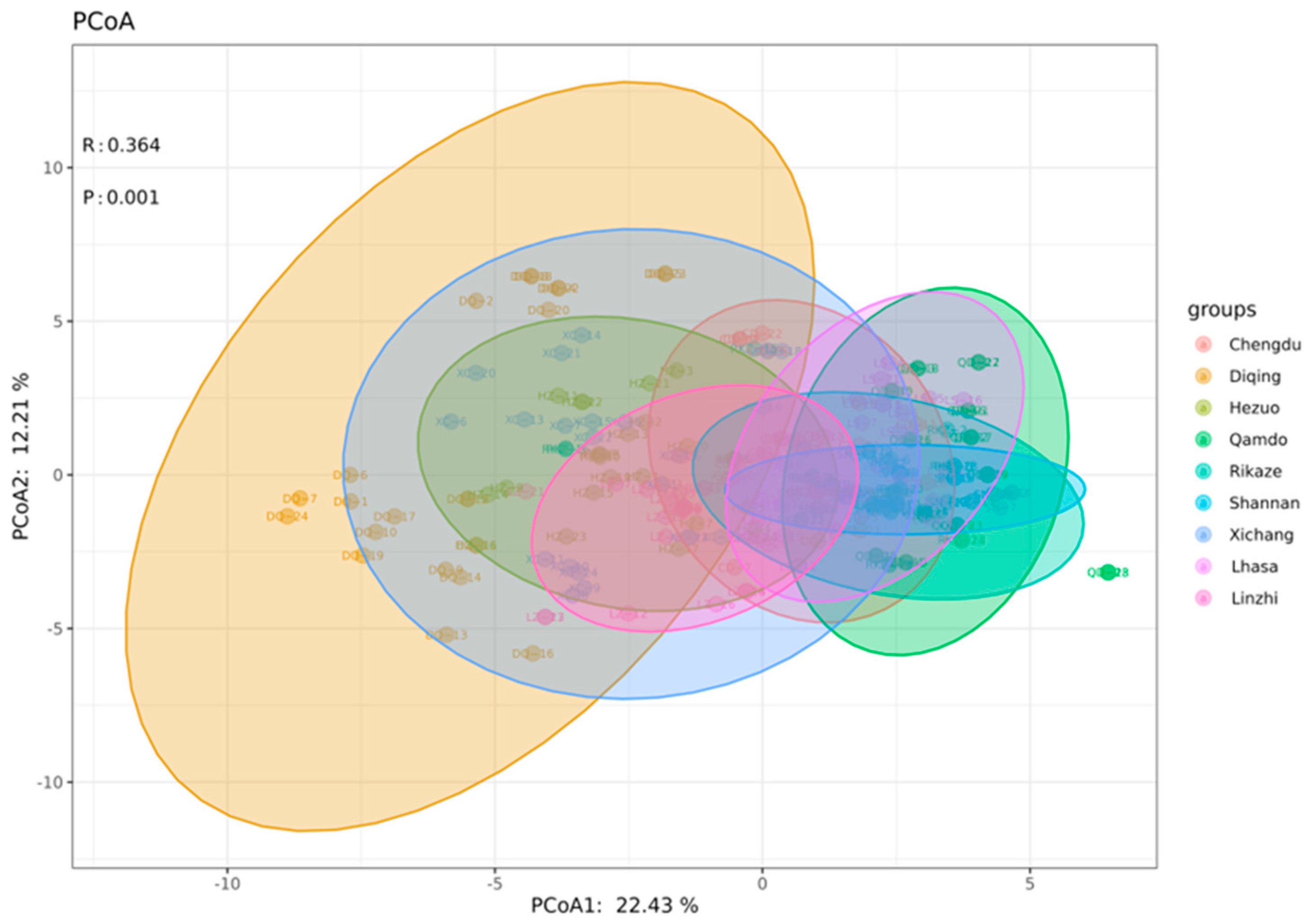
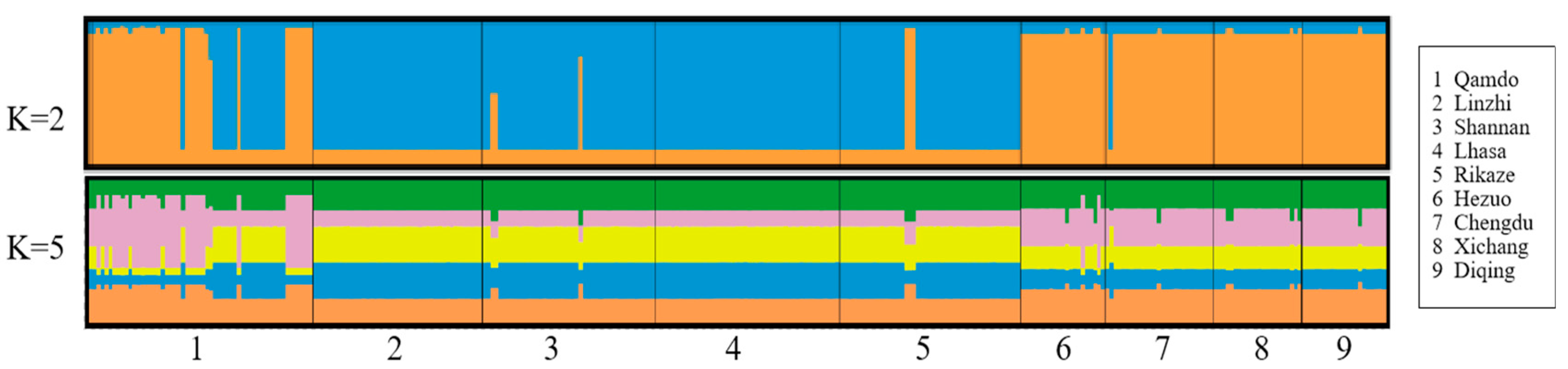
3.2. Molecular Genetic Structure of the Bgh Populations
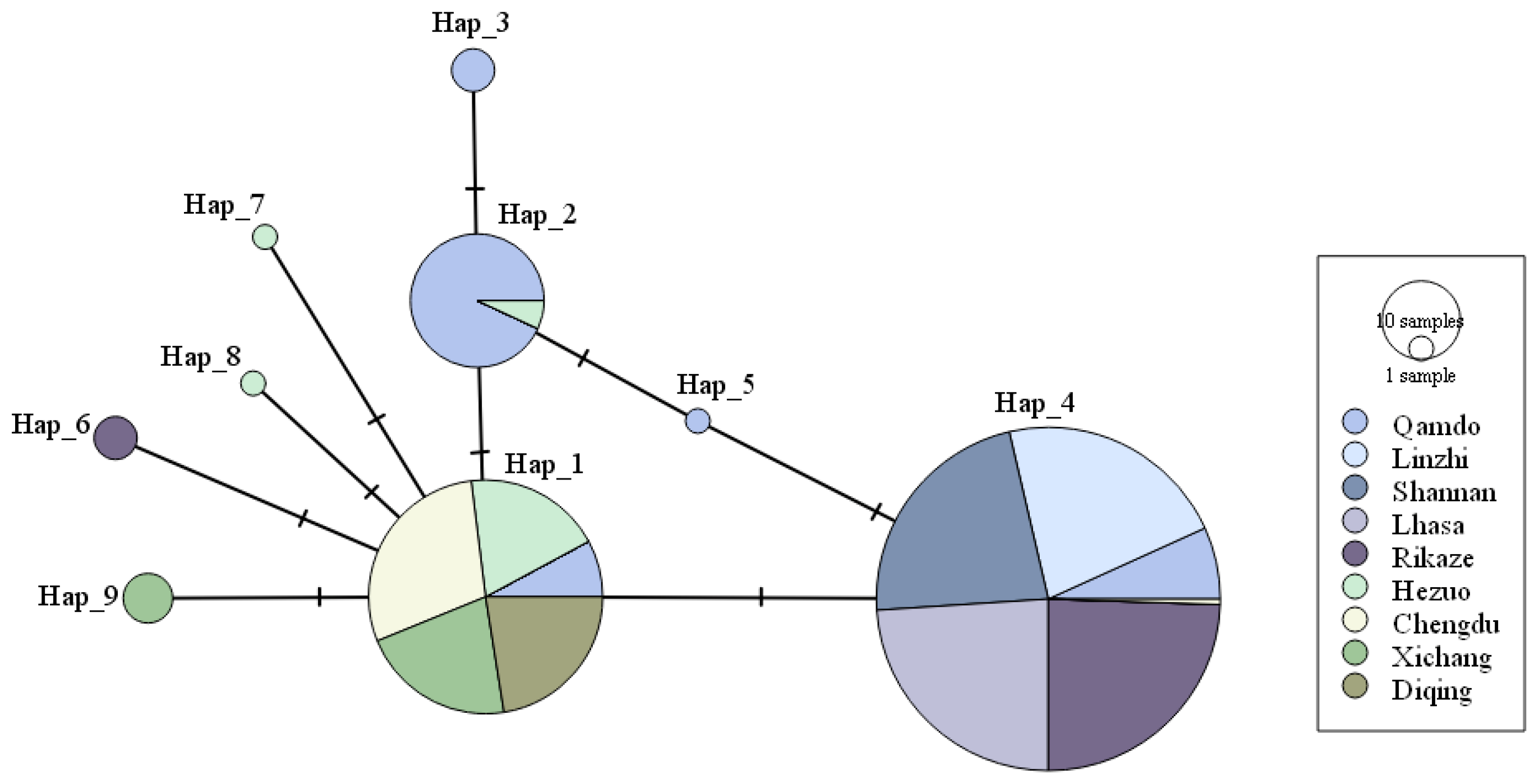
3.2.1. Haplotype Composition of Different Bgh Population
3.2.2. Genotype Diversity of Bgh Populations
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Variety | Resistance Genes | Origin |
|---|---|---|---|
| 1 | P01 | Mla1, Mla(Al2) | Denmark |
| 2 | P02 | Mla3 | Denmark |
| 3 | P03 | Mla6, Mla14 | Denmark |
| 4 | P04A | Mla7,Mlk | Denmark |
| 5 | P04B | Mla7, Mla(No3) | Denmark |
| 6 | P05 | / | Denmark |
| 7 | P06 | Mla7Ml(Lg2) | Denmark |
| 8 | P07 | Mla9, Mlk | Denmark |
| 9 | P08A | Mla9, Mlk | Denmark |
| 10 | P08B | Mla9 | Denmark |
| 11 | P09 | Mla10, MlaDu2 | Denmark |
| 12 | P10 | Mla12, MlaEm2 | Denmark |
| 13 | P11 | Mla13, MlRu3, MlaRu4 | Denmark |
| 14 | P12 | Mla22 | Denmark |
| 15 | P13 | Mla23 | Denmark |
| 16 | P14 | Mlra | Denmark |
| 17 | P15 | Ml(Ru2) | Denmark |
| 18 | P16 | Mlk | Denmark |
| 19 | P17 | Mlk1 | Denmark |
| 20 | P18 | Mlnn | Denmark |
| 21 | P19 | Mlp1 | Denmark |
| 22 | P20 | Mlat, Mla8 | Denmark |
| 23 | P21 | Mlg, Ml(CP) | Denmark |
| 24 | P22 | mlo5 | Denmark |
| 25 | P23 | MlLa | Denmark |
| 26 | P24 | Mlh | Denmark |
| 27 | P29 | / | Denmark |
| 28 | P30 | / | Denmark |
| 29 | P31 | / | Denmark |
| 30 | Pallas | Mla8 | Sweden |
| Pathotypes | |||||
|---|---|---|---|---|---|
| 1.0.1.0.0.0.0 | 3.0.0.0.0.0.0 | 3.4.3.2.3.0.0 | 5.6.6.1.1.0.0 | 6.5.1.5.6.0.0 | 7.2.0.0.0.0.0 |
| 1.0.1.4.0.0.0 | 3.0.0.4.0.0.0 | 3.5.0.0.0.0.0 | 5.7.0.0.0.0.0 | 6.5.4.0.0.0.0 | 7.2.1.0.0.0.0 |
| 1.2.0.0.0.0.0 | 3.0.2.0.0.0.0 | 3.5.1.0.1.0.0 | 6.0.0.0.0.0.0 | 6.6.4.0.0.0.0 | 7.2.4.0.0.0.0 |
| 1.2.0.0.1.0.0 | 3.0.3.0.0.0.0 | 3.5.1.2.2.0.0 | 6.0.1.3.0.0.0 | 6.7.3.3.2.0.0 | 7.2.5.5.4.0.0 |
| 1.2.2.0.1.0.0 | 3.0.3.2.0.0.0 | 3.5.1.4.5.0.0 | 6.0.2.0.0.0.0 | 7.0.0.0.0.0.0 | 7.2.6.5.0.0.0 |
| 1.3.0.0.0.0.0 | 3.0.3.4.1.0.0 | 3.5.3.2.2.0.0 | 6.0.4.0.0.0.0 | 7.0.0.2.0.0.0 | 7.3.0.0.0.0.0 |
| 1.3.0.0.2.0.0 | 3.0.3.5.1.0.0 | 4.0.3.4.2.0.0 | 6.1.2.0.0.0.0 | 7.0.1.0.0.0.0 | 7.3.3.0.0.0.0 |
| 1.3.1.6.0.0.0 | 3.0.3.6.1.0.0 | 4.0.4.0.0.0.0 | 6.1.5.0.0.0.0 | 7.0.1.0.4.0.0 | 7.3.4.0.0.0.0 |
| 1.4.0.0.0.0.0 | 3.0.3.7.1.0.0 | 4.4.2.0.0.0.0 | 6.1.7.1.4.0.0 | 7.0.1.1.4.0.0 | 7.3.4.1.0.0.0 |
| 1.5.4.0.0.0.0 | 3.1.0.0.0.0.0 | 4.6.0.0.0.0.0 | 6.2.5.0.0.0.0 | 7.0.1.3.2.0.0 | 7.3.6.5.0.0.0 |
| 1.5.4.1.0.0.0 | 3.1.1.0.0.0.0 | 4.7.0.0.0.0.0 | 6.2.5.3.1.0.0 | 7.0.2.0.0.0.0 | 7.4.0.0.0.0.0 |
| 1.7.0.0.0.0.0 | 3.1.1.2.3.0.0 | 5.0.0.0.5.1.0 | 6.2.5.7.0.0.0 | 7.0.3.2.0.0.0 | 7.4.1.3.0.0.0 |
| 2.0.0.0.0.0.0 | 3.1.3.2.2.0.0 | 5.0.1.0.4.0.0 | 6.2.7.3.1.0.0 | 7.0.3.4.1.0.0 | 7.4.1.3.2.0.0 |
| 2.0.4.6.0.0.0 | 3.2.3.3.1.0.0 | 5.0.2.0.4.0.0 | 6.3.2.2.5.0.0 | 7.0.4.0.0.0.0 | 7.4.2.0.0.0.0 |
| 2.0.5.6.0.0.0 | 3.2.4.0.0.0.0 | 5.1.2.0.0.0.0 | 6.3.4.2.2.0.0 | 7.1.0.0.0.0.0 | 7.4.3.2.0.0.0 |
| 2.0.7.0.0.0.0 | 3.2.4.0.0.1.0 | 5.1.2.0.4.0.0 | 6.3.5.7.0.0.0 | 7.1.0.0.4.0.0 | 7.4.3.6.0.0.0 |
| 2.0.7.2.0.0.0 | 3.3.0.0.0.0.0 | 5.1.2.0.5.1.0 | 6.4.2.0.1.0.0 | 7.1.1.0.0.0.0 | 7.5.0.0.0.0.0 |
| 2.1.0.0.1.0.0 | 3.3.3.5.1.0.0 | 5.1.3.0.0.0.0 | 6.4.4.0.0.0.0 | 7.1.1.0.4.0.0 | 7.5.0.0.1.0.0 |
| 2.2.2.0.0.0.0 | 3.3.3.7.0.0.0 | 5.3.0.0.0.0.0 | 6.4.6.0.1.0.0 | 7.1.2.0.0.0.0 | 7.5.0.0.3.0.0 |
| 2.5.0.0.0.0.0 | 3.3.3.7.1.0.0 | 5.3.0.1.0.0.0 | 6.5.0.0.0.0.0 | 7.1.2.0.1.0.0 | 7.6.0.0.0.0.0 |
| 2.6.4.0.0.0.0 | 3.4.0.0.0.0.0 | 5.4.0.0.0.0.0 | 6.5.0.1.0.0.0 | 7.1.3.1.1.0.0 | 7.7.0.0.0.0.0 |
| 2.7.0.0.1.0.0 | 3.4.3.0.3.0.0 | 5.6.2.1.0.0.0 | 6.5.1.4.0.0.0 | 7.1.4.6.0.0.0 | 7.7.0.0.1.0.0 |
References
- IGC. 2019. Available online: https://www.igc.int/en/default.aspx (accessed on 5 August 2022).
- Çelik, O.; Karakaya, A. Genetic diversity of barley foliar fungal pathogens. Agronomy 2021, 11, 434. [Google Scholar] [CrossRef]
- Dreiseitl, A.; Yang, J. Powdery mildew resistance in a collection of Chinese barley varieties. Genet. Resour. Crop Evol. 2007, 54, 2. [Google Scholar] [CrossRef]
- Jia, X.; Sun, Z.; Li, X. Layout of barley production and its comparative Advantage in China. Agric. Outlook 2017, 13, 40–46. [Google Scholar]
- Zeng, Y.; Pu, X.; Zhang, J. Synthetic research and utilization on industrial development of barley in Southwestern China. J. Agric. Sci. Technol. 2013, 15, 48–56. [Google Scholar]
- Qiang, X.; Chi, D.; Feng, J. Status of the production and development of highland barley in the Qinghai-Tibet plateau region. Tibet Sci. Technol. 2008, 3, 11–17. [Google Scholar]
- Nyima, T.; Yu, D.; Tang, Y.; Bian, B.; Zeng, X. Great improvement of the yield of highland barley to ensure the safety of highland barley. Tibet Sci. Technol. 2013, 2, 5–7. [Google Scholar]
- Yuan, H.; Zeng, X.; Yang, Q.; Xu, Q.; Wang, Y.; Jabu, D.; Sang, Z.; Tashi, N. Gene coexpression network analysis combined with metabonomics reveals the resistance responses to powdery mildew in Tibetan hulless barley. Sci. Rep. 2018, 8, 14928. [Google Scholar] [CrossRef] [Green Version]
- Mathre, D.E. Compendium of Barley Disease, 2nd ed.; American Phytopathological Society: St. Paul, MN, USA, 1997. [Google Scholar]
- Yahyaoui, A.; Reinhold, M.; Scharen, A. Virulence spectrum in populations of the barley powdery mildew pathogen Erysiphe graminis f. sp. hordei in Tunisia and Morocco in 1992. Plant Pathol. 1997, 46, 139–146. [Google Scholar]
- Dreiseitl, A. Pathogenic divergence of central European and Australian populations of Blumeria graminis f. sp. hordei. Ann. Appl. Biol. 2014, 165, 364–372. [Google Scholar] [CrossRef]
- Zhu, J.; Zhou, Y.; Yang, J. Advances in the study of genetics of Blumeria graminis f. sp. hordei. J. Plant Prot. 2014, 41, 109–117. [Google Scholar]
- Chaure, P.; Gurr, S.; Spanu, P. Stable transformation of Erysiphe graminis an obligate biotrophic pathogen of barley. Nat. Biotechnol. 2000, 18, 205–207. [Google Scholar] [CrossRef]
- Jørgensen, J.; Jensen, H. Powdery mildew resistance in barley landrace material. I. Screening for resistance. Euphytica 1997, 97, 227–233. [Google Scholar] [CrossRef]
- Dreiseitl, A. Specific resistance of barley to powdery mildew, Its use and beyond. A concise critical review. Genes 2020, 11, 971. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Li, Y.; Gao, Q.; Qin, X.; Ma, X.; Liu, G. Climate change trend and its impact on yield of highland barley in Tibet, China. J. Agric. Resour. Environ. 2018, 35, 374–380. [Google Scholar]
- Liu, G. Effect of climate change on Tibetan agricultural production. Tibet J. Agric. Sci. 2019, 41, 49–54. [Google Scholar]
- Reinstädler, A.; Müller, J.; Czembor, J.; Piffanelli, P.; Panstruga, R. Novel induced mlo mutant alleles in combination with site-directed mutagenesis reveal functionally important domains in the heptahelical barley Mlo protein. BMC Plant Biol. 2010, 10, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parks, R.; Carbone, I.; Murphy, J.; Cowger, C. Population genetic analysis of an eastern U.S. wheat powdery mildew population reveals geographic subdivision and recent common ancestry with U.K. and Israeli populations. Phytopathology 2009, 99, 840–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Duan, X.; Zhou, Y. Population genetic analysis of Blumeria graminis f. sp. tritici in Qinghai Province, China. J. Integr. Agric. 2014, 13, 1952–1961. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, G.; Mu, W.; Lin, R. Virulence variability and genetic diversity in Blumeria graminis f. sp. hoedei in southeastern and southwestern China. Plant Dis. 2022, 8, 10. [Google Scholar]
- Jensen, H.; Christensen, E.; Jørgensen, J. Powdery Mildew Resistance Genes in 127 Northwest European Spring Barley Varieties. Plant Breed. 1992, 108, 210–228. [Google Scholar] [CrossRef]
- Gilmour, J. Octal notation for designating physiologic races of plant pathogens. Nature 1973, 242, 620. [Google Scholar] [CrossRef]
- Limpert, E.; Müller, K. Designation of pathotypes of plant pathogens. J. Phytopathol. 1994, 140, 346–358. [Google Scholar] [CrossRef]
- Cowger, C.; Parks, R.; Kosman, E. Structure and migration in U.S. Blumeria graminis f. sp. tritici populations. Phytopathology 2016, 106, 295–304. [Google Scholar] [PubMed] [Green Version]
- Schachtel, G.; Dinoor, A.; Herrmann, A.; Kosman, E. Comprehensive evaluation of virulence and resistance data: A new analysis tool. Plant Dis. 2012, 96, 1060–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Earl, D.A.; vonHoldt, B.M. Structure Harvester: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [Green Version]
- Excoffier, L.; Lischer, H. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Dupanloup, I.; Schneider, S.; Excoffier, L. A simulated annealing approach to define the genetic structure of populations. Mol. Ecol. 2002, 11, 2571–2581. [Google Scholar] [CrossRef]
- Wang, J.; Dreiseitl, A. Virulence and diversity of Blumeria graminis f. sp. hordei in East China. Eur. J. Plant Pathol. 2007, 117, 357–368. [Google Scholar]
- Dreiseitl, A. Great pathotype diversity and reduced virulence complexity in a central European population of Blumeria graminis f. sp. hordei in 2015–2017. Eur. J. Plant Pathol. 2019, 153, 801–811. [Google Scholar] [CrossRef]
- Dreiseitl, A. Genotype heterogeneity in accessions of a winter barley core collection assessed on postulated specific powdery mildew resistance genes. Agronomy 2021, 11, 513. [Google Scholar] [CrossRef]
- Helen, R.; Dreiseitl, A.; Sadiki, M.; Daniel, J. High diversity, low spatial structure and rapid pathotype evolution in Moroccan populations of Blumeria graminis f. sp. hordei. Eur. J. Plant Pathol. 2013, 136, 323–336. [Google Scholar]
- Zeng, X.; Guo, Y.; Xu, Q.; Mascher, M.; Guo, G.; Li, S.; Mao, L.; Liu, Q.; Xia, Z.; Zhou, J.; et al. Origin and evolution of qingke barley in Tibet. Nat. Commun. 2018, 9, 5433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S. Diseases, Pests and Weeds in Tibet Agriculture; The Tibet People’s Publisher: Lhasa, China, 1987; pp. 393–394. [Google Scholar]





| Variety | Resistance Gene(s) | Virulence Frequency (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| QD a | LZ | SN | LS | RKZ | HZ | CD | XC | DQ | Total | ||
| P01 | Mla1, Mla(Al2) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| P02 | Mla3 | 0 | 35.71 | 0 | 0 | 0 | 16.67 | 11.54 | 25.00 | 0 | 9.88 |
| P03 | Mla6, Mla14 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| P04A | Mla7, Mlk | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| P04B | Mla7, Mla(No3) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| P05 | / | 0 | 21.43 | 0 | 0 | 0 | 16.67 | 7.69 | 41.67 | 58.33 | 16.20 |
| P06 | Mla7, Ml(Lg2) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| P07 | Mla9, Mlk | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| P08A | Mla9, Mlk | 0 | 0 | 0 | 4.55 | 0 | 0 | 0 | 0 | 0 | 0.50 |
| P08B | Mla9 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| P09 | Mla10, MlaDu2 | 60.00 | 100 | 66.67 | 100 | 70.00 | 91.67 | 76.92 | 100 | 100 | 85.03 |
| P10 | Mla12, MlaEm2 | 6.67 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.74 |
| P11 | Mla13, Ru3, Ru4 | 40.00 | 0 | 0 | 54.54 | 3.70 | 0 | 23.08 | 54.00 | 33.33 | 23.18 |
| P12 | Mla22 | 53.33 | 100 | 83.33 | 63.64 | 77.78 | 75.00 | 65.38 | 33.00 | 58.33 | 67.75 |
| P-13 | Mla23 | 0 | 21.43 | 16.67 | 13.64 | 0 | 12.50 | 19.23 | 4.17 | 54.17 | 15.76 |
| P-14 | Mlra | 0 | 21.43 | 0 | 0 | 0 | 54.17 | 65.38 | 46.00 | 66.67 | 28.18 |
| P15 | Ml(Ru2) | 60.00 | 78.57 | 83.33 | 81.82 | 44.44 | 83.33 | 76.92 | 92.00 | 91.67 | 76.90 |
| P16 | Mlk | 0 | 21.43 | 0 | 0 | 7.41 | 0 | 15.38 | 33.00 | 6.25 | 9.27 |
| P17 | Mlk1 | 0 | 7.14 | 0 | 0 | 3.70 | 62.50 | 0 | 37.50 | 75.00 | 20.65 |
| P18 | Mlnn | 60.00 | 64.29 | 71.00 | 72.73 | 48.15 | 83.33 | 84.62 | 50.00 | 41.67 | 63.98 |
| P19 | Mlp1 | 0 | 75.00 | 0 | 0 | 7.41 | 8.75 | 7.69 | 71.00 | 91.67 | 29.06 |
| P20 | Mlat, Mla8 | 73.33 | 21.43 | 66.67 | 72.73 | 14.81 | 25.00 | 23.08 | 16.67 | 16.67 | 36.71 |
| P21 | Mlg, Ml(CP) | 0 | 0 | 0 | 18.18 | 0 | 54.17 | 23.08 | 33.00 | 58.33 | 20.75 |
| P22 | mlo5 | 0 | 0 | 0 | 4.55 | 22.22 | 16.67 | 0 | 21.00 | 12.50 | 8.55 |
| P23 | MlLa | 46.67 | 71.43 | 91.67 | 50.00 | 40.74 | 29.17 | 15.38 | 54.00 | 33.33 | 48.04 |
| P24 | Mlh | 33.33 | 27.43 | 66.67 | 36.36 | 59.26 | 12.5 | 30.77 | 0 | 58.33 | 36.07 |
| P29 | / | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| P30 | / | 73.33 | 92.86 | 91.67 | 81.82 | 85.19 | 100 | 92.31 | 100 | 100 | 90.80 |
| P31 | / | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Pallas | Mla8 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| QD | LZ | SN | LS | RKZ | HZ | CD | XC | DQ | |
|---|---|---|---|---|---|---|---|---|---|
| Number of isolates | 30 | 28 | 24 | 22 | 27 | 24 | 26 | 24 | 24 |
| Number of pathotypes | 13 | 18 | 10 | 18 | 15 | 16 | 21 | 22 | 19 |
| Average virulence complexity per isolate (Ci) | 3.733 | 5.393 | 5.000 | 4.682 | 3.370 | 6.042 | 4.808 | 5.625 | 7.625 |
| Relative virulence complexity per isolate (Rci) | 0.178 | 0.257 | 0.238 | 0.223 | 0.160 | 0.288 | 0.229 | 0.268 | 0.363 |
| Genotypic diversity (sh) | 0.742 | 0.846 | 0.670 | 0.903 | 0.797 | 0.836 | 0.912 | 0.964 | 0.891 |
| Gene diversity (HS) | 0.162 | 0.165 | 0.107 | 0.173 | 0.165 | 0.209 | 0.205 | 0.261 | 0.256 |
| Genetic diversity (KWm) | 0.248 | 0.248 | 0.143 | 0.264 | 0.250 | 0.298 | 0.267 | 0.385 | 0.385 |
| Populations | QD | LZ | SN | LS | RKZ | HZ | CD | XC | DQ |
|---|---|---|---|---|---|---|---|---|---|
| QD | - | 0.184 2 | 0.103 | 0.184 | 0.111 | 0.237 | 0.144 | 0.18 | 0.282 |
| LZ | 0.105 1 | - | 0.147 | 0.182 | 0.157 | 0.151 | 0.162 | 0.144 | 0.234 |
| SN | 0.044 | 0.08 | - | 0.082 | 0.119 | 0.220 | 0.170 | 0.240 | 0.292 |
| LS | 0.017 | 0.102 | 0.041 | - | 0.139 | 0.151 | 0.162 | 0.169 | 0.234 |
| RKZ | 0.048 | 0.074 | 0.057 | 0.062 | - | 0.201 | 0.156 | 0.184 | 0.237 |
| HZ | 0.14 | 0.07 | 0.143 | 0.119 | 0.114 | - | 0.157 | 0.127 | 0.151 |
| CD | 0.072 | 0.085 | 0.098 | 0.06 | 0.071 | 0.083 | - | 0.152 | 0.206 |
| XC | 0.101 | 0.061 | 0.134 | 0.086 | 0.093 | 0.052 | 0.061 | - | 0.155 |
| DQ | 0.185 | 0.119 | 0.192 | 0.158 | 0.151 | 0.07 | 0.112 | 0.057 | - |
| Haplotype | AOX a | PKA | PPA | QD | LZ | SN | LS | RKZ | HZ | CD | XC | DQ | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| H1 | A b | G | A | G | A | C | C | 6 | 20 | 23 | 22 | 24 | ||||
| H2 | G | 21 | 2 | |||||||||||||
| H3 | G | G | 3 | |||||||||||||
| H4 | A | 15 | 43 | 45 | 44 | 39 | 1 | |||||||||
| H5 | G | A | 1 | |||||||||||||
| H6 | T | 3 | ||||||||||||||
| H7 | G | 1 | ||||||||||||||
| H8 | A | 1 | ||||||||||||||
| H9 | A | 4 | ||||||||||||||
| Total | 46 | 43 | 45 | 44 | 42 | 24 | 24 | 26 | 24 | |||||||
| Populations | Number of Isolates | Haplotypes Diversityjof (Hd) | Nucleotide Diversity (π) | Average Number of Difference (K) |
|---|---|---|---|---|
| QD | 46 | 0.6485 | 0.00049 | 1.00735 |
| LZ | 43 | 0.000 | 0.00000 | 0.00000 |
| SN | 45 | 0.000 | 0.00000 | 0.00000 |
| LS | 44 | 0.000 | 0.00000 | 0.00000 |
| RKZ | 45 | 0.1151 | 0.00001 | 0.23020 |
| HZ | 24 | 0.3477 | 0.00017 | 0.37143 |
| CD | 24 | 0.3117 | 0.00007 | 0.07407 |
| XC | 26 | 0.1453 | 0.00014 | 0.31169 |
| DQ | 24 | 0.0000 | 0.00000 | 0.00000 |
| Total | 321 | 0.5780 | 0.00034 | 0.74034 |
| Source of Variance | d.f | Sum of Squares | Variance Components | Percentage of Variation (%) | Fixation Indices |
|---|---|---|---|---|---|
| Among groups | 1 | 45.268 | 0.31240 Va | 54.68 | FCT = 0.54681 |
| Among populations | 8 | 35.626 | 0.13655 Vb | 23.90 | FST = 0.69714 |
| Within populations | 313 | 38.302 | 0.12237 Vc | 21.42 | |
| Total | 322 | 119.195 | 0.57132 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zhuoma, Q.; Xu, Z.; Peng, Y.; Wang, M. Virulence and Genetic Types of Blumeria graminis f. sp. hordei in Tibet and Surrounding Areas. J. Fungi 2023, 9, 363. https://doi.org/10.3390/jof9030363
Wang Y, Zhuoma Q, Xu Z, Peng Y, Wang M. Virulence and Genetic Types of Blumeria graminis f. sp. hordei in Tibet and Surrounding Areas. Journal of Fungi. 2023; 9(3):363. https://doi.org/10.3390/jof9030363
Chicago/Turabian StyleWang, Yunjing, Qucuo Zhuoma, Zhi Xu, Yunliang Peng, and Mu Wang. 2023. "Virulence and Genetic Types of Blumeria graminis f. sp. hordei in Tibet and Surrounding Areas" Journal of Fungi 9, no. 3: 363. https://doi.org/10.3390/jof9030363
APA StyleWang, Y., Zhuoma, Q., Xu, Z., Peng, Y., & Wang, M. (2023). Virulence and Genetic Types of Blumeria graminis f. sp. hordei in Tibet and Surrounding Areas. Journal of Fungi, 9(3), 363. https://doi.org/10.3390/jof9030363