
Mechanisms of Surfactin from Bacillus subtilis SF1 against Fusarium foetens: A Novel Pathogen Inducing Potato Wilt

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Pathogen Isolation and Pathogenicity Tests
2.2. Pathogen Identification
2.3. The Antifungal Activity of B. subtilis SF1 against F. foetens In Vitro
2.4. Effect of B. subtilis SF1 on Fusarium Wilt Incidence
2.5. Hydrolytic Enzymes Production
2.6. Plant Growth Promoting-Determinants Produced by B. subtilis SF1
2.7. Extraction, Purification, and Identification of Surfactin Produced by B. subtilis SF1
2.8. Effect of Surfactin on the Growth and Morphology of F. foetens
2.9. Effect of Surfactin on the Nucleic Acids and Proteins of F. foetens
2.10. Effect of Surfactin on Transcription Level of F. foetens
2.11. Effect of Surfactin on GSH Production of F. foetens
2.12. DNA Binding Assay
2.13. Statistical Analysis
3. Results
3.1. Molecular and Morphological Characteristics of the F. foetens Pathogen
3.1.1. Isolation of the Pathogen and Molecular Identification
3.1.2. Morphological Characteristics of F. foetens
3.1.3. Pathogenicity Test
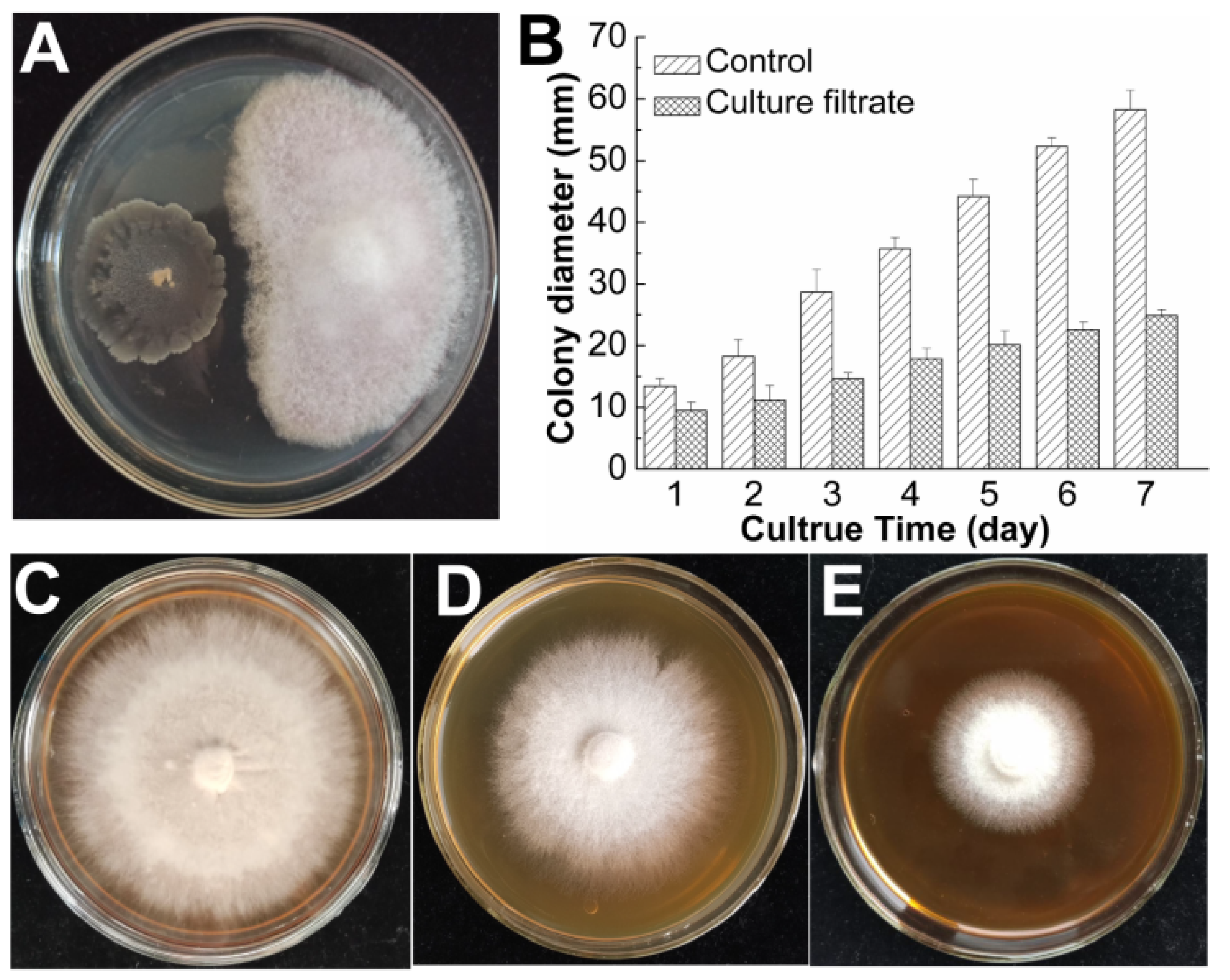
3.2. Evaluation of the Antifungal and Plant Growth-Promoting Properties of B. subtilis SF1
3.2.1. The Antifungal Activity of B. subtilis SF1 against F. foetens In Vitro
3.2.2. Effect of B. subtilis SF1 on FW Incidence In Vivo
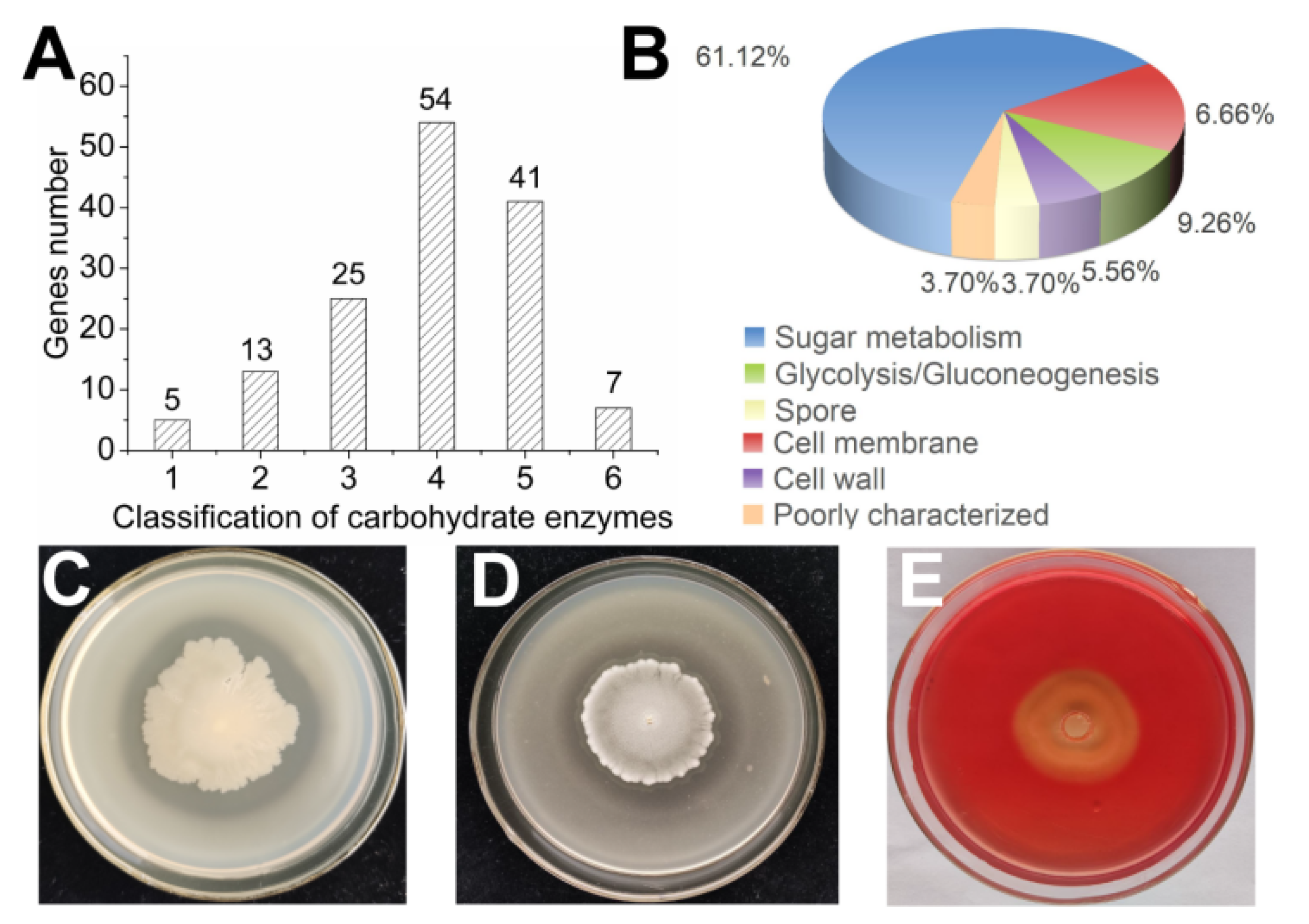
3.2.3. Production of Hydrolytic Enzymes
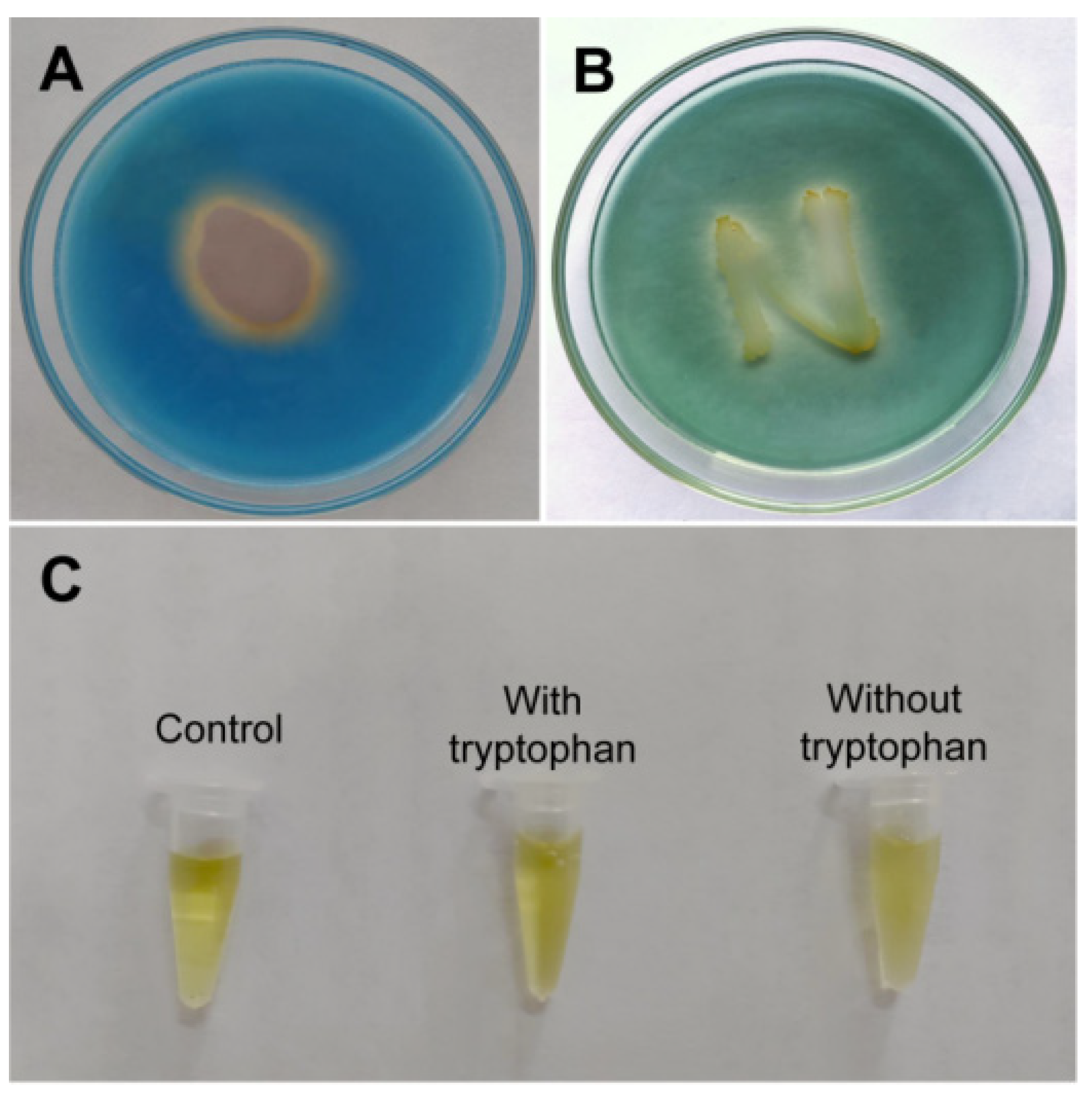
3.2.4. Plant Growth-Promoting Determinants Produced by B. subtilis SF1
3.3. Purification, Identification and Antifungal Mechanisms of the Surfactin from B. subtilis SF1
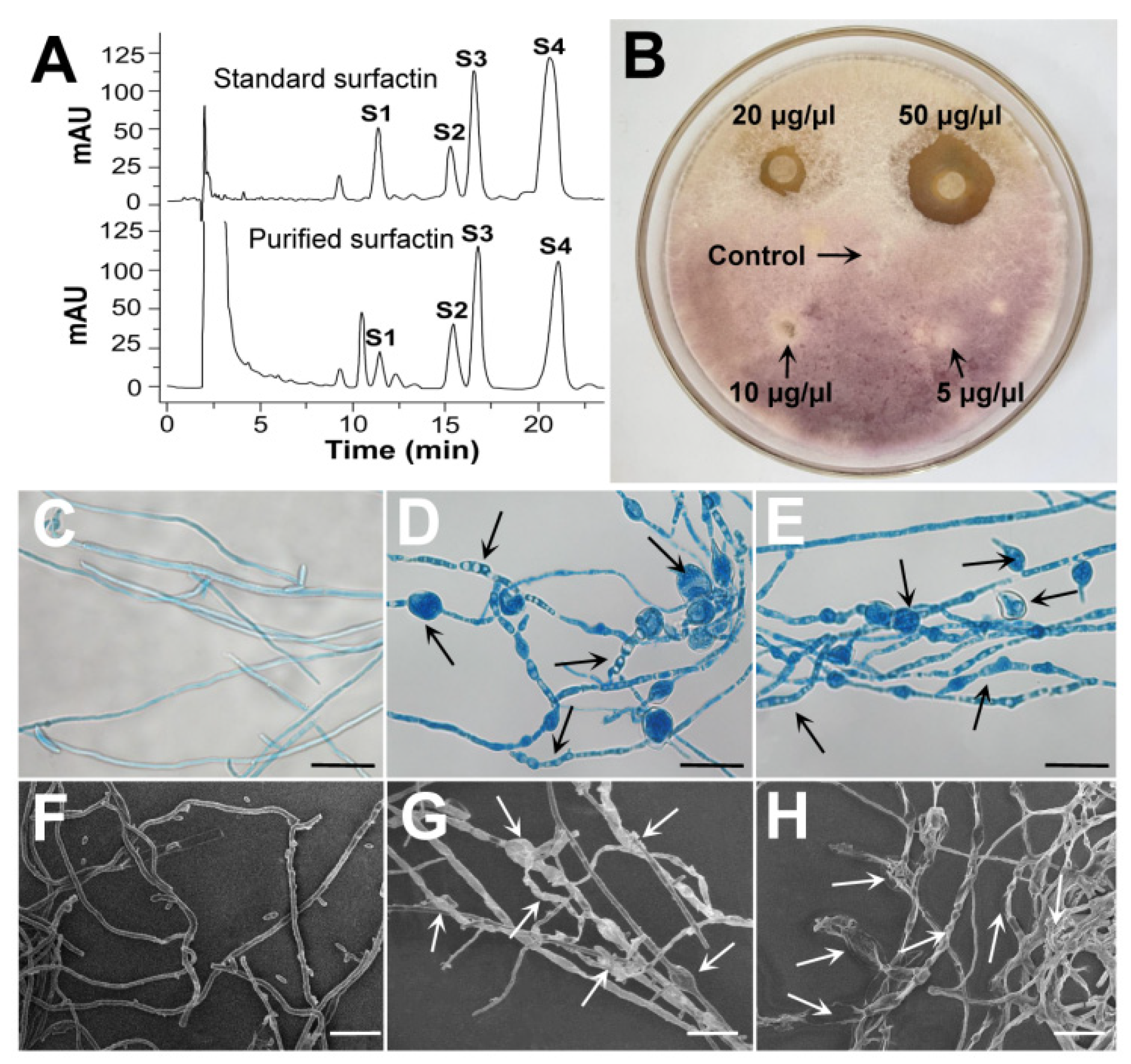
3.3.1. Purification and Identification of the Surfactin Produced by B. subtilis SF1
3.3.2. Effect of Surfactin on the Growth and Morphology of F. foetens
3.3.3. Effect of Surfactin on F. foetens Membrane Integrity
3.3.4. Effect of Surfactin on F. foetens Protein Expression
3.3.5. Effect of Surfactin on GSH Production by F. foetens
3.3.6. Effect of Surfactin on F. foetens Transcription Levels
3.3.7. Interaction between Surfactin and ff-DNA by Electrophoresis, UV-Visible Spectroscopy and Fluorescence Spectroscopy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Devaux, A.; Kromann, P.; Ortiz, O. Potatoes for sustainable global food security. Potato Res. 2014, 57, 185–199. [Google Scholar] [CrossRef]
- Tiwari, R.K.; Kumar, R.; Sharma, S.; Sagar, V.; Aggarwal, R.; Naga, K.C.; Kumar, M. Potato dry rot disease: Current status, pathogenomics and management. 3 Biotech 2020, 10, 503. [Google Scholar] [CrossRef] [PubMed]
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2005, 144, 31–43. [Google Scholar] [CrossRef]
- Larkin, R.P.; Halloran, J.M. Management effects of disease-suppressive rotation crops on potato yield and soilborne disease and their economic implications in potato production. Am. J. Potato Res. 2014, 91, 429–439. [Google Scholar] [CrossRef]
- Xia, S.Y.; Niu, Z.M.; Li, Q.Q.; Zhang, L.J.; Sheng, W.M. Research progress and control measures of Fusarium wilt of potato. Heilongjiang Agric. Sci. 2022, 2, 89–94. [Google Scholar]
- Nxumalo, N.N. Occurrence, Identification and a Potential Management Strategy of Fusarium Species Causing wilt of Potatoes in South Africa. Dissertation. Univ. Pretoria 2013. Available online: http://hdl.handle.net/2263/33188 (accessed on 12 November 2022).
- Mejdoub-Trabelsi, B.; Touihri, S.; Ammar, N.; Riahi, A.; Daami-Remadi, M. Effect of chitosan for the control of potato diseases caused by Fusarium species. J. Phytopathol. 2019, 168, 18–27. [Google Scholar] [CrossRef]
- Paul, N.C.; Park, W.; Lee, S.; Chung, M.N.; Lee, H.U.; Yang, J.W. Occurrence of sweetpotato (Ipomoea batatas) wilt and surface rot disease and determining resistance of selected varieties to the pathogen in Korea. Plants 2020, 9, 497. [Google Scholar] [CrossRef] [PubMed]
- Ayed, F.; Daami-Remadi, M.; Jabnoun-Khiareddine, H.; Hibar, K.; Mahjoub, M.E. Evaluation of fungicides for control of Fusarium wilt of potato. Plant Pathol. J. 2006, 5, 239–243. [Google Scholar] [CrossRef] [Green Version]
- Ayed, F.; Daami-Remadi, M.; Jabnoun-Khiareddine, H.; Hibar, K.; Mahjoub, M.E. Effect of potato cultivars on incidence of Fusaium oxysporum f. sp. tuberosi and its transmission to progeny tubers. Agron. J. 2006, 5, 430–434. [Google Scholar] [CrossRef] [Green Version]
- Khedher, S.B.; Mejdoub-Trabelsi, B.; Tounsi, S. Biological potential of Bacillus subtilis V26 for the control of Fusarium wilt and tuber dry rot on potato caused by Fusarium species and the promotion of plant growth. Biol. Control 2020, 152, 104444. [Google Scholar] [CrossRef]
- Gouda, S.; Kerry, R.G.; Das, G.; Paramithiotis, S.; Shin, H.S.; Patra, J.K. Revitalization of plant growth promoting rhizobacteria for sustainable development in agriculture. Microbiol. Res. 2018, 206, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Arrebola, E.; Sivakumar, D.; Bacigalupo, R.; Korsten, L. Combined application of antagonist Bacillus amyloliquefaciens and essential oils for the control of peach postharvest diseases. Crop Prot. 2010, 29, 369–377. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Lu, Y.; Shan, M.; Zhao, H.; Lu, Z.; Lu, Y. A mini-review: Mechanism of antimicrobial action and application of surfactin. World J. Microbiol. Biotechnol. 2022, 38, 143. [Google Scholar] [CrossRef]
- Dimkić, I.; Živković, S.; Berić, T.; Ivanović, Ž.; Gavrilović, V.; Stanković, S.; Fira, D. Characterization and evaluation of two Bacillus strains, SS-12.6 and SS-13.1, as potential agents for the control of phytopathogenic bacteria and fungi. Biol. Control 2013, 65, 312–321. [Google Scholar] [CrossRef]
- Yuan, B.; Xu, P.Y.; Zhang, Y.J.; Wang, P.P.; Yu, H.; Jiang, J.H. Synthesis of biocontrol macromolecules by derivative of chitosan with surfactin and antifungal evaluation. Int. J. Biol. Macromol. 2014, 66, 7–14. [Google Scholar] [CrossRef]
- Jiang, J.; Gao, L.; Bie, X.; Lu, Z.; Liu, H.; Zhang, C.; Zhao, H. Identification of novel surfactin derivatives from NRPS modification of Bacillus subtilis and its antifungal activity against Fusarium moniliforme. BMC Microbiol. 2016, 16, 31. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, M.; Dheeman, S.; Dubey, R.C.; Kumar, P.; Maheshwari, D.K.; Bajpai, V.K. Differential antagonistic responses of Bacillus pumilus MSUA3 against Rhizoctonia solani and Fusarium oxysporum causing fungal diseases in Fagopyrum esculentum moench. Microbiol. Res. 2017, 205, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, N.; Velramar, B.; Velu, R.K. Investigation of antifungal activity of surfactin against mycotoxigenic phytopathogenic fungus Fusarium moniliforme and its impact in seed germination and mycotoxicosis. Pestic. Biochem. Physiol. 2019, 155, 101–107. [Google Scholar] [CrossRef]
- Elmer, W.H.; Vossbrinck, C.; Geiser, D.M. First report of a wilt disease of Hiemalis begonias caused by Fusarium foetens in the United States. Plant Dis. 2004, 88, 1287. [Google Scholar] [CrossRef]
- Schroers, H.J.; Baayen, R.P.; Meffert, J.P.; de Gruyter, J.; Hooftman, M.; O’Donnell, K. Fusarium foetens, a new species pathogenic to begonia elatior hybrids (Begonia × hiemalis) and the sister taxon of the Fusarium oxysporum species complex. Mycologia 2004, 96, 393–406. [Google Scholar] [CrossRef]
- Tian, X.L.; Dixon, M.; Zheng, Y. First report of Hiemalis begonias wilt disease caused by Fusarium foetens in Canada. Plant Dis. 2010, 94, 1261. [Google Scholar] [CrossRef] [PubMed]
- Saurat, C.; Fourrier, C.; Wilson, V.; Casset, C.; Ioos, R. First report of begonia elatior wilt disease caused by Fusarium foetens in France. Plant Dis. 2013, 97, 144. [Google Scholar] [CrossRef] [PubMed]
- Lamprecht, S.C.; Tewoldemedhin, Y.T. Fusarium species associated with damping-off of rooibos seedlings and the potential of compost as soil amendment for disease suppression. S. Afr. J. Bot. 2017, 110, 110–117. [Google Scholar] [CrossRef]
- Amobonye, A.; Bhagwat, P.; Ranjith, D.; Mohanlall, V.; Pillai, S. Characterisation, pathogenicity and hydrolytic enzyme profiling of selected Fusarium species and their inhibition by novel coumarins. Arch. Microbiol. 2021, 203, 3495–3508. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.L.; Zhang, Y.J.; Guo, L.Z.; Liu, L. First report of Colletotrichum gloeosporioides causing leaf spot on Cyclobalanopsis glauca in China. Plant Dis. 2021, 105, 3303. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Baayen, R.P.; O’Donnell, K.; Bonants, P.J.M.; Cigelnik, E.; Kroon, L.P.N.M.; Roebroeck, E.J.A.; Waalwijk, C. Gene genealogies and AFLP analyses in the Fusarium oxysporum complex identify monophyletic and nonmonophyletic formae speciales causing wilt and rot disease. Phytopathology 2000, 90, 891–900. [Google Scholar] [CrossRef] [Green Version]
- Sarwar, A.; Hassan, M.N.; Imran, M.; Iqbal, M.; Majeed, S.; Brader, G.; Hafeez, F.Y. Biocontrol activity of surfactin A purified from Bacillus NH-100 and NH-217 against rice bakanae disease. Microbiol. Res. 2018, 209, 1–13. [Google Scholar] [CrossRef]
- Li, S.S. Field Application of Bacillus amyloliquefaciens S3-1 and Technology Optimization of Surfactin Fermentation. Master’s Thesis, Shanghai Normal University, Shanghai, China, 2017. Available online: https://kns.cnki.net/KCMS/detail/detail.aspx?dbname=CMFD201801&filename=1017161589.nh (accessed on 2 January 2022).
- Yokota, K.; Hayakawa, H. Impact of antimicrobial lipopeptides from Bacillus sp. on suppression of fusarium yellows of tatsoi. Microbes Environ. 2015, 30, 281–283. [Google Scholar] [CrossRef] [Green Version]
- Teather, R.M.; Wood, P.J. Use of congo red-polysaccharide interactions in enumeration and characterization of cellulolytic bacteria from the bovine rumen. Appl. Environ. Microbiol. 1982, 43, 777–780. [Google Scholar] [CrossRef] [Green Version]
- Dinesh, R.; Anandaraj, M.; Kumar, A.; Bini, Y.K.; Subila, K.P.; Aravind, R. Isolation, characterization, and evaluation of multi-trait plant growth promoting rhizobacteria for their growth promoting and disease suppressing effects on ginger. Microbiol. Res. 2015, 173, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Frey-Klett, P.; Chavatte, M.; Clausse, M.L.; Courrier, S.; Roux, C.L.; Raaijmakers, J.; Garbaye, J. Ectomycorrhizal symbiosis affects functional diversity of rhizosphere fluorescent pseudomonads. New Phytol. 2004, 65, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Bric, J.M.; Bostock, R.M.; Silverstone, S.E. Rapid in situ assay for indoleacetic acid production by bacteria immobilized on a nitrocellulose membrane. Appl. Environ. Microbiol. 1991, 57, 535–538. [Google Scholar] [CrossRef] [Green Version]
- Luo, C.; Liu, X.; Zhou, H.; Wang, X.; Chen, Z. Nonribosomal peptide synthase gene clusters for lipopeptide biosynthesis in Bacillus subtilis 916 and their phenotypic functions. Appl. Environ. Microbiol. 2015, 81, 422–431. [Google Scholar] [CrossRef] [Green Version]
- Jones, R.N.; Ballow, C.H.; Biedenbach, D.J. Multi-laboratory assessment of the linezolid spectrum of activity using the Kirby-Bauer disk diffusion method: Report of the Zyvox® Antimicrobial Potency Study (ZAPS) in the United States. Diagn. Microbiol. Infect. Dis. 2001, 40, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lu, J.; Sun, J.; Lu, F.; Bie, X.; Lu, Z. Membrane disruption and DNA binding of Fusarium graminearum cell induced by C16-Fengycin A produced by Bacillus amyloliquefaciens. Food Control 2019, 102, 206–213. [Google Scholar] [CrossRef]
- Ahmadi, F.; Alizadeh, A.A.; Bakhshandeh-Saraskanrood, F.; Jafari, B.; Khodadadian, M. Experimental and computational approach to the rational monitoring of hydrogen-bonding interaction of 2-Imidazolidinethione with DNA and guanine. Food Chem. Toxicol. 2010, 48, 29–36. [Google Scholar] [CrossRef]
- Oguzcan, E.; Koksal, Z.; Taskin-Tok, T.; Uzgoren-Baran, A.; Akbay, N. Spectroscopic and molecular modeling methods to investigate the interaction between psycho-stimulant modafinil and calf thymus dna using ethidium bromide as a fluorescence probe. Spectrochim. Acta Part A 2022, 270, 120787. [Google Scholar] [CrossRef]
- Rehman, S.U.; Sarwar, T.; Husain, M.A.; Ishqi, H.M.; Tabish, M. Studying non-covalent drug-DNA interactions. Arch. Biochem. Biophys. 2015, 576, 49–60. [Google Scholar] [CrossRef]
- Cao, Y.; He, X. Studies of interaction between Safranine T and double helix DNA by spectral methods. Spectrochim. Acta Part A 1998, 54, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Marty, R.; N’soukpoe-Kossi, C.N.; Charbonneau, D.; Weinert, C.M.; Kreplak, L.; Tajmir-Riahi, H.A. Structural analysis of DNA complexation with cationic lipids. Nucleic Acids Res. 2008, 37, 849–857. [Google Scholar] [CrossRef]
- Ni, Y.; Wang, Y.; Kokot, S. Study of the interaction between 10-hydroxycamptothecine and DNA with the use of ethidium bromide dye as a fluorescence probe. Sens. Actuators B 2011, 1561, 290–297. [Google Scholar] [CrossRef]
- Howe-Grant, M.; Wu, K.C.; Bauer, W.R.; Lippard, S.J. Binding of platinum and palladium metallointercalation reagents and antitumor drugs to closed and open DNAs. Biochemistry 1976, 15, 4339–4346. [Google Scholar] [CrossRef] [PubMed]
- Sehlstedt, U.; Kim, S.K.; Carter, P.; Goodisman, J.; Vollano, J.F.; Norden, B.; Dabrowiak, J.C. Interaction of cationic porphyrins with DNA. Biochemistry 1994, 33, 417–426. [Google Scholar] [CrossRef]
- Tsan, P.; Volpon, L.; Besson, F.; Lancelin, J.M. Structure and dynamics of surfactin studied by NMR in micellar media. J. Am. Chem. Soc. 2007, 129, 1968–1977. [Google Scholar] [CrossRef] [PubMed]
- Bojanowski, A.; Avis, T.J.; Pelletier, S.; Tweddell, R.J. Management of potato dry rot. Postharvest Biol. Technol. 2013, 84, 99–109. [Google Scholar] [CrossRef]
- Chung, S.; Kong, H.; Buyer, J.S.; Lakshman, D.K.; Lydon, J.; Kim, S.D.; Roberts, D.P. Isolation and partial characterization of Bacillus subtilis ME488 for suppression of soilborne pathogens of cucumber and pepper. Appl. Microbiol. Biotechnol. 2008, 80, 115–123. [Google Scholar] [CrossRef]
- Yu, X.; Ai, C.; Xin, L.; Zhou, G. The siderophore-producing bacterium, Bacillus subtilis CAS15, has a biocontrol effect on Fusarium wilt and promotes the growth of pepper. Eur. J. Soil Biol. 2011, 47, 138–145. [Google Scholar] [CrossRef]
- Zhang, L.; Khabbaz, S.E.; Wang, A.; Li, H.; Abbasi, P.A. Detection and characterization of broad-spectrum antipathogen activity of novel rhizobacterial isolates and suppression of Fusarium crown and root rot disease of tomato. J. Appl. Microbiol. 2015, 118, 685–703. [Google Scholar] [CrossRef]
- Zhao, Q.; Ran, W.; Wang, H.; Li, X.; Shen, Q.; Shen, S.; Xu, Y. Biocontrol of Fusarium wilt disease in muskmelon with Bacillus subtilis Y-IVI. Bio. Control 2012, 58, 283–292. [Google Scholar] [CrossRef]
- De Weerdt, M.; Zijlstra, C.; Van Brouwershaven, I.R.; Van Leeuwen, G.C.M.; De Gruyter, J.; Kox, L.F.F. Molecular detection of Fusarium foetens in Begonia. J. Phytopathol. 2006, 154, 694–700. [Google Scholar] [CrossRef]
- Chang, W.T.; Chen, M.L.; Wang, S.L. An antifungal chitinase produced by Bacillus subtilis using chitin waste as a carbon source. World J. Microbiol. Biotechnol. 2010, 26, 945–950. [Google Scholar] [CrossRef]
- Luo, Y.; Sun, L.; Zhu, Z.; Ran, W.; Shen, Q. Identification and characterization of an anti-fungi Fusarium oxysporum f. sp. cucumerium protease from the Bacillus subtilis strain N7. J. Microbiol. 2013, 51, 359–366. [Google Scholar] [CrossRef]
- Podile, A.R.; Prakash, A.P. Lysis and biological control of Aspergillus niger by Bacillus subtilis AF 1. Can. J. Microbiol. 1996, 42, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Aktuganov, G.E.; Galimzyanova, N.F.; Melent’ev, A.I.; Kuz’mina, L.Y. Extracellular hydrolases of strain Bacillus sp. 739 and their involvement in the lysis of micromycete cell walls. Microbiology 2007, 76, 413–420. [Google Scholar] [CrossRef]
- Siahmoshteh, F.; Hamidi-Esfahani, Z.; Spadaro, D.; Shams-Ghahfarokhi, M.; Razzaghi-Abyaneh, M. Unraveling the mode of antifungal action of Bacillus subtilis and Bacillus amyloliquefaciens as potential biocontrol agents against aflatoxigenic Aspergillus parasiticus. Food Control 2018, 89, 300–307. [Google Scholar] [CrossRef]
- Hu, L.B.; Shi, Z.Q.; Zhang, T.; Yang, Z.M. Fengycin antibiotics isolated from B-FS01 culture inhibit the growth of Fusarium moniliforme Sheldon ATCC 38932. FEMS Microbiol. Lett. 2007, 272, 91–98. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.Z.; Zheng, Q.W.; Wei, T.; Zhang, Z.Q.; Zhao, C.F.; Zhong, H.; Guo, L.Q. Isolation and characterization of fengycins produced by Bacillus amyloliquefaciens JFL21 and its broad-spectrum antimicrobial potential against multidrug-resistant foodborne pathogens. Front. Microbiol. 2020, 11, 3319. [Google Scholar] [CrossRef]
- Zhao, Z.; Wang, Q.; Wang, K.; Brian, K.; Liu, C.; Gu, Y. Study of the antifungal activity of Bacillus vallismortis ZZ185 in vitro and identification of its antifungal components. Bioresour. Technol. 2010, 101, 292–297. [Google Scholar] [CrossRef]
- Sheppard, J.D.; Jumarie, C.; Cooper, D.G.; Laprade, R. Ionic channels induced by surfactin in planar lipid bilayer membranes. Biochim. Biophys. Acta Biomembr. 1991, 1064, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, C.; Teruel, J.A.; Aranda, F.J.; Ortiz, A. Molecular mechanism of membrane permeabilization by the peptide antibiotic surfactin. Biochim. Biophys. Acta Biomembr. 2003, 1611, 91–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Tan, T.; Shen, A.; Yang, X.; Yu, Y.; Gao, C.; Zeng, L. Biocontrol potential of Bacillus subtilis IBFCBF-4 against Fusarium wilt of watermelon. J. Plant Pathol. 2020, 102, 433–441. [Google Scholar] [CrossRef]
- Backhouse, D.; Stewart, A. Interactions between Bacillus species and sclerotia of Sclerotium cepivorum. Soil Biol. Biochem. 1989, 21, 173–176. [Google Scholar] [CrossRef]
- Benoit, I.; van den Esker, M.H.; Patyshakuliyeva, A.; Mattern, D.J.; Blei, F.; Zhou, M.; Kovács, Á.T. Bacillus subtilis attachment to Aspergillus nigerhyphae results in mutually altered metabolism. Environ. Microbiol. 2014, 17, 2099–2113. [Google Scholar] [CrossRef] [PubMed]
- Ágnes, J.; Fruzsina, K.; Noémi, B.; Zoltán, T.; Ágota, R.; Zsófi, S.; Kinga, C.; Csaba, N.K.; Dániel, N.; Ildikó, B.; et al. Physiological and transcriptional profiling of surfactin exerted antifungal effect against Candida albicans. Biomed. Pharmacother. 2022, 152, 0753–3322. [Google Scholar] [CrossRef]
- Wu, S.; Liu, G.; Zhou, S.; Sha, Z.; Sun, C. Characterization of antifungal lipopeptide biosurfactants produced by marine bacterium Bacillus sp. CS30. Mar. Drugs 2019, 17, 199. [Google Scholar] [CrossRef] [Green Version]
- Pócsi, I.; Prade, R.A.; Penninckx, M.J. Glutathione, altruistic metabolite in fungi. Adv. Microb. Physiol. 2004, 49, 1–76. [Google Scholar] [CrossRef]
- Boer, D.R.; Canals, A.; Coll, M. DNA-binding drugs caught in action: The latest 3D pictures of drug-DNA complexes. Dalton Trans. 2009, 399–414. [Google Scholar] [CrossRef] [Green Version]
- Turner, P.R.; Denny, W.A. The mutagenic properties of DNA minor-groove binding ligands. Microbiol. Res. 1996, 355, 141–169. [Google Scholar] [CrossRef]
- Nelson, S.M.; Ferguson, L.R.; Denny, W.A. Non-covalent ligand/DNA interactions: Minor groove binding agents. Mutat. Res. 2007, 623, 24–40. [Google Scholar] [CrossRef] [PubMed]
- Cuellar-Gaviria, T.Z.; González-Jaramillo, L.M.; Villegas-Escobar, V. Role of Bacillus tequilensis EA-CB0015 cells and lipopeptides in the biological control of black Sigatoka disease. Biol. Control 2021, 155, 104523. [Google Scholar] [CrossRef]
- Dong, L.H.; Wang, P.P.; Zhao, W.S.S.; Su, Z.H.; Zhang, X.Y.; Lu, X.Y.; Li, S.Z.; Ma, P.; Guo, Q.G. Surfactin and fengycin contribute differentially to the biological activity of Bacillus subtilis NCD-2 against cotton verticillium wilt. Biol. Control 2022, 174, 104999. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, L.; Jin, X.; Lu, X.; Guo, L.; Lu, P.; Yu, H.; Lv, B. Mechanisms of Surfactin from Bacillus subtilis SF1 against Fusarium foetens: A Novel Pathogen Inducing Potato Wilt. J. Fungi 2023, 9, 367. https://doi.org/10.3390/jof9030367
Liu L, Jin X, Lu X, Guo L, Lu P, Yu H, Lv B. Mechanisms of Surfactin from Bacillus subtilis SF1 against Fusarium foetens: A Novel Pathogen Inducing Potato Wilt. Journal of Fungi. 2023; 9(3):367. https://doi.org/10.3390/jof9030367
Chicago/Turabian StyleLiu, Lin, Xiaofan Jin, Xiuhua Lu, Lizhong Guo, Peiwei Lu, Hao Yu, and Beibei Lv. 2023. "Mechanisms of Surfactin from Bacillus subtilis SF1 against Fusarium foetens: A Novel Pathogen Inducing Potato Wilt" Journal of Fungi 9, no. 3: 367. https://doi.org/10.3390/jof9030367
APA StyleLiu, L., Jin, X., Lu, X., Guo, L., Lu, P., Yu, H., & Lv, B. (2023). Mechanisms of Surfactin from Bacillus subtilis SF1 against Fusarium foetens: A Novel Pathogen Inducing Potato Wilt. Journal of Fungi, 9(3), 367. https://doi.org/10.3390/jof9030367