
Neofusicoccum cryptomeriae sp. nov. and N. parvum Cause Stem Basal Canker of Cryptomeria japonica in China
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Fungal Isolation
2.2. Pathogenicity Tests
2.3. Morphological Identification
2.4. DNA Extraction, PCR Amplification, and Sequencing
2.5. Phylogenetic Analyses
2.6. Genealogical Concordance Phylogenetic Species Recognition Analysis
3. Results
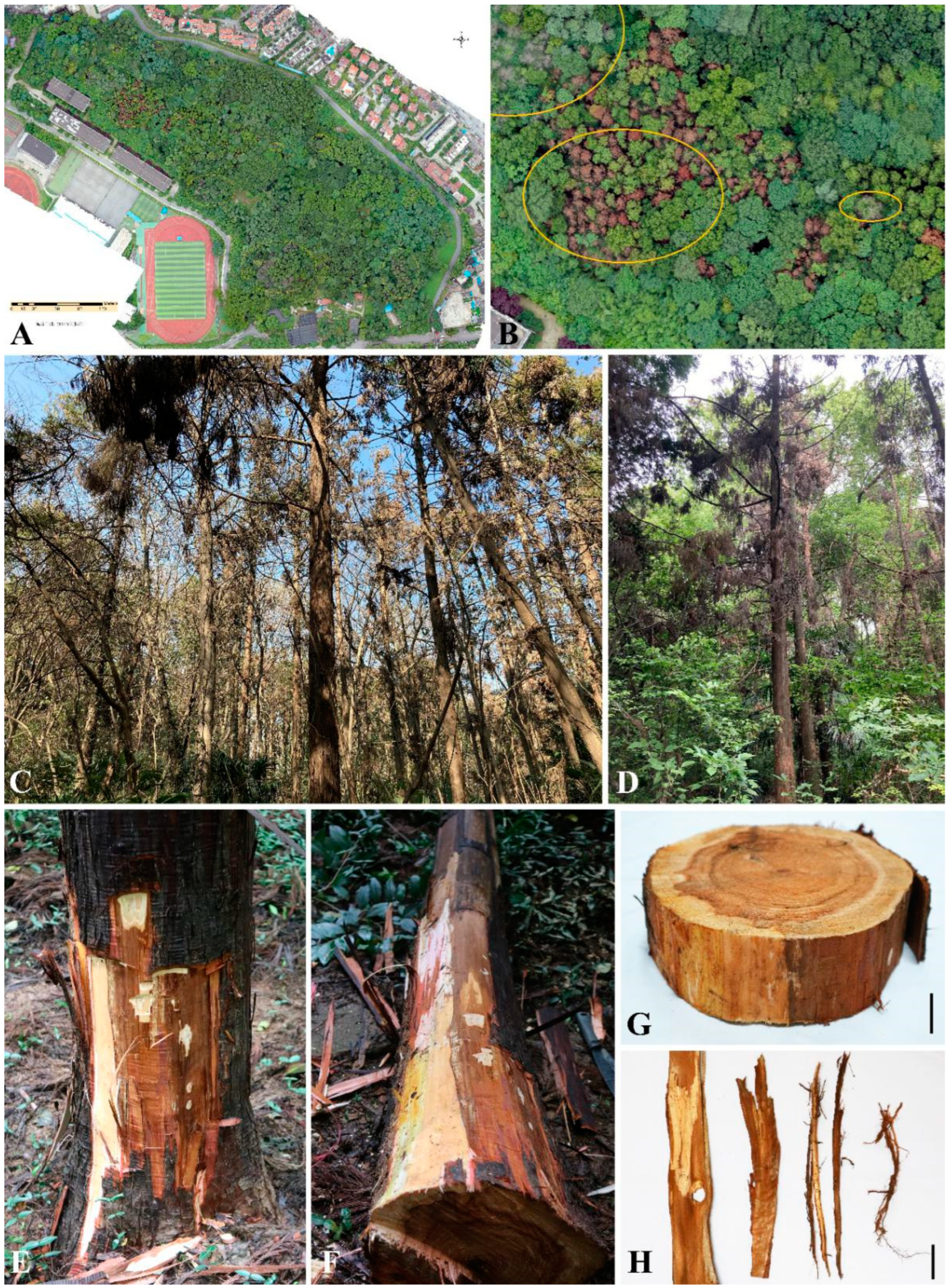
3.1. Symptoms in the Field and Fungal Isolation
3.2. Pathogenicity Assays
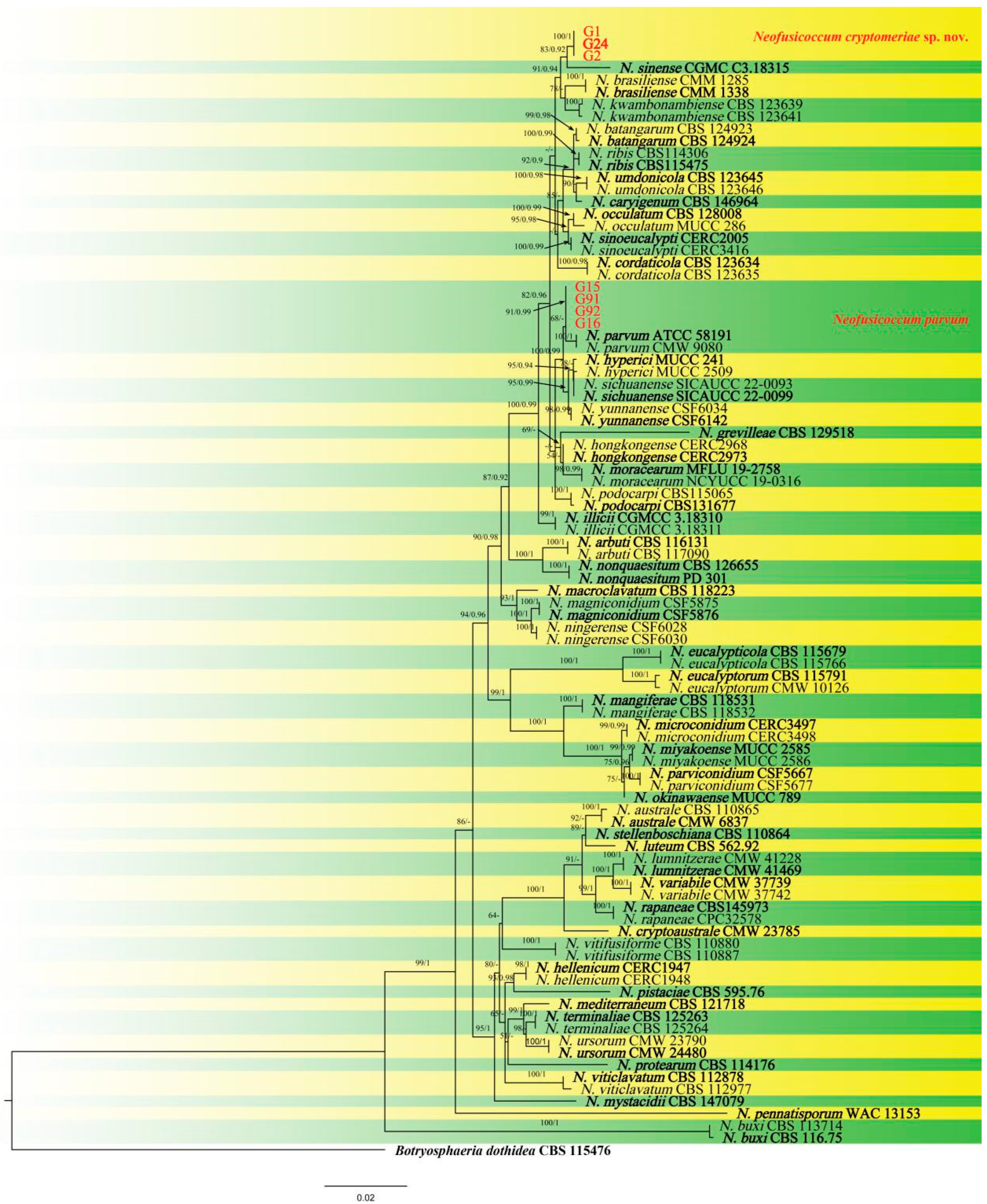
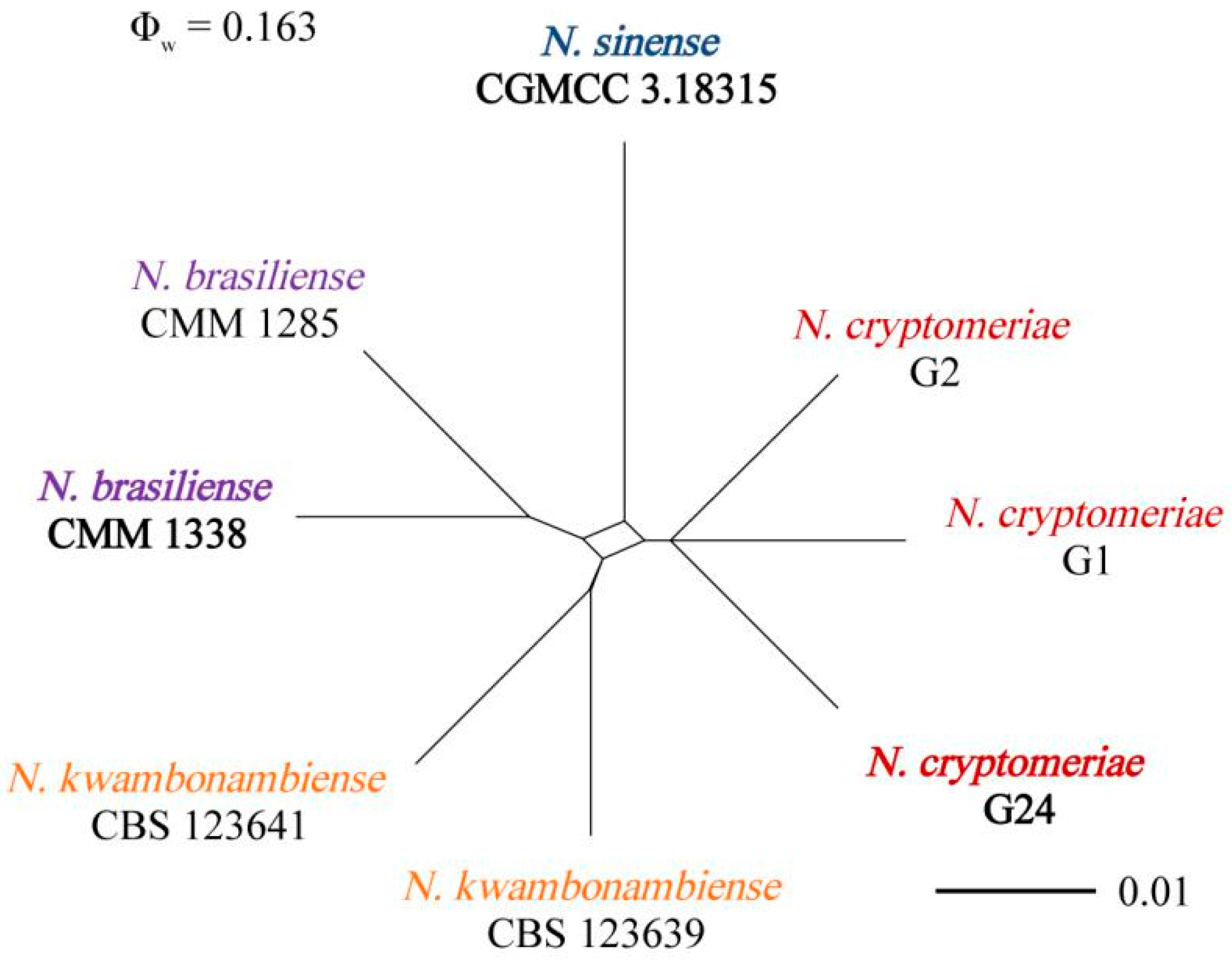
3.3. Molecular Identification of the Fungal Isolates
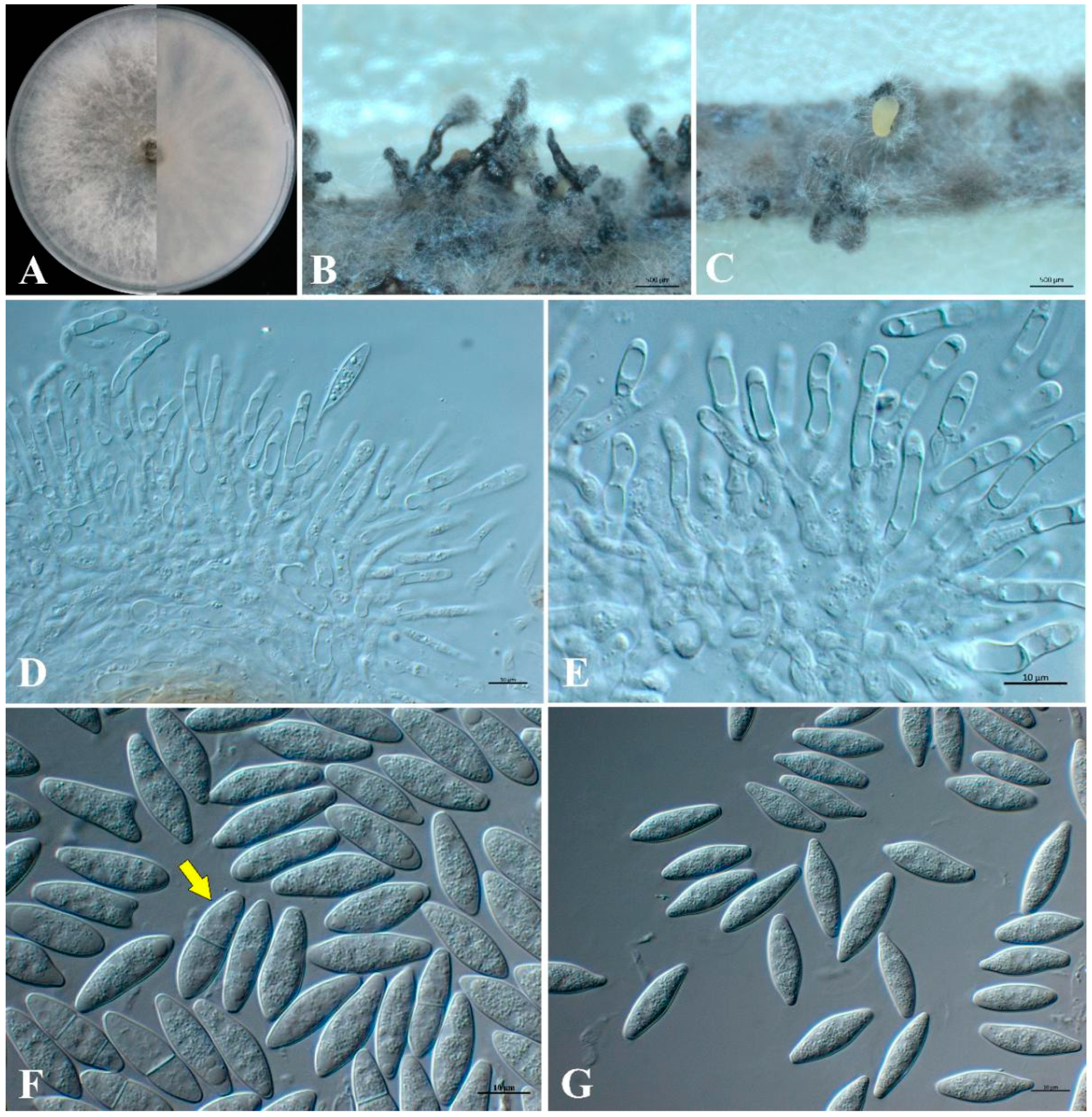
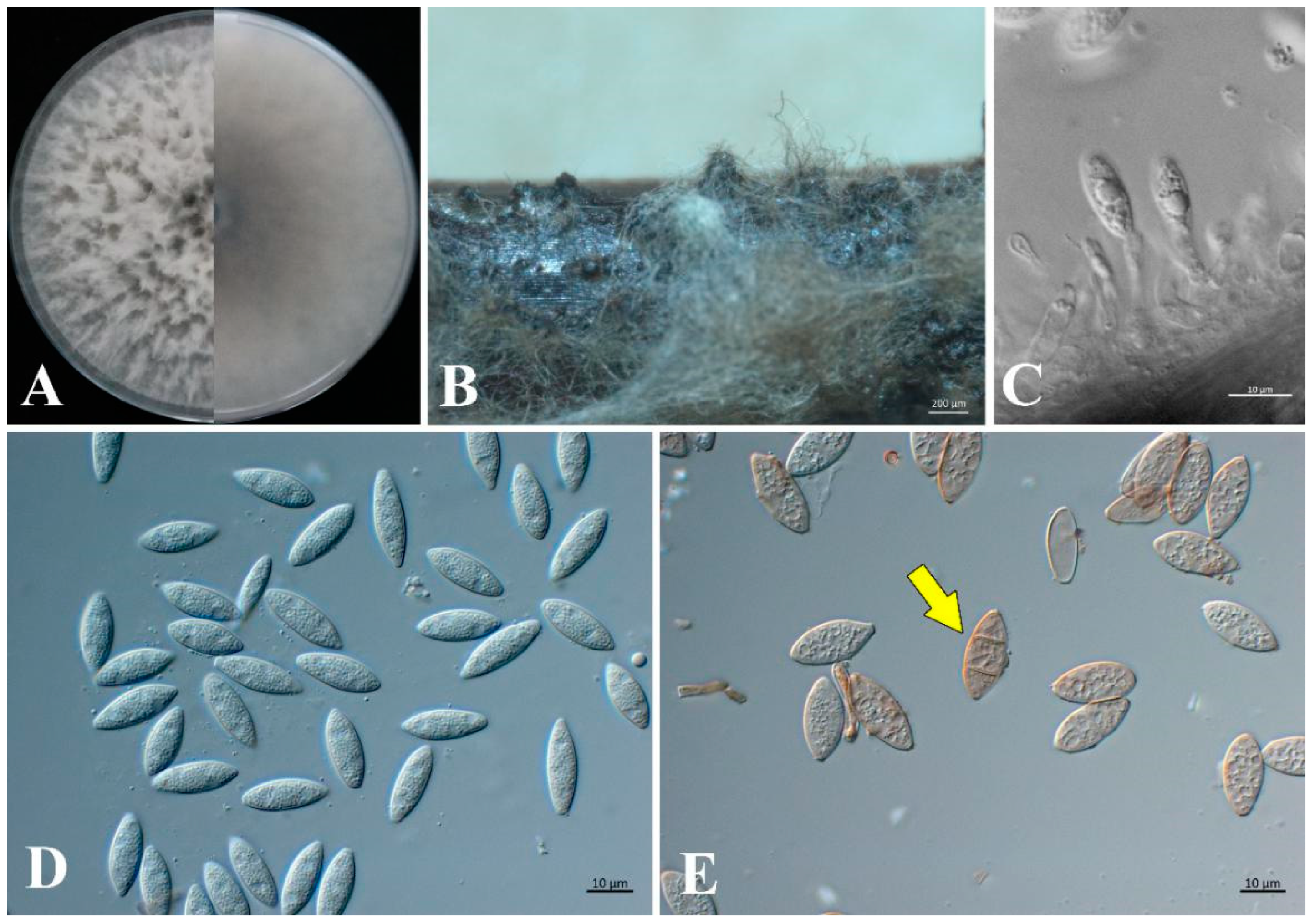
3.4. Morphology and Taxonomy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yan, Y.; Wu, X.; Wang, M.; Li, Z.; Yuan, M.; Dai, M.; Wen, Y. Complete chloroplast genomes of wild and cultivated Cryptomeria japonica var. sinensis. Biotechnol. Biotechnol. Eq. 2021, 35, 821–827. [Google Scholar] [CrossRef]
- POWO. Plants of the World Online. 2023. Available online: https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:261870-1 (accessed on 10 February 2023).
- Chen, Y.; Yang, S.Z.; Zhao, M.S.; Ni, B.Y.; Liu, L.; Chen, X.Y. Demographic Genetic Structure of Cryptomeria japonica var. sinensis in Tianmushan Nature Reserve, China. J. Integr. Plant Biol. 2008, 50, 1171–1177. [Google Scholar] [CrossRef] [PubMed]
- Zhejiang Department of Forestry Ancient and Famous Trees. Zhejiang Ancient and Famous Trees; Zhejiang Scientific and Technology Press: Hangzhou, China, 2001. [Google Scholar]
- Iizuka, K.; Toya, N.; Ohshima, J.; Ishiguri, F.; Miyamoto, N.; Aizawa, M.; Ohkubo, T.; Takenaka, C.; Yokota, S. Relationship between Cs-137 concentration and potassium content in stem wood of Japanese cedar (Cryptomeria japonica). J. Wood Sci. 2018, 64, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Hanai, Y.; Takenaka, C.; Tomioka, R.; Iizuka, K.; Ozawa, H. Cesium absorption through bark of Japanese cedar (Cryptomeria japonica). J. For. Res. -Jpn. 2016, 21, 251–258. [Google Scholar] [CrossRef]
- Lima, A.; Arruda, F.; Janeiro, A.; Medeiros, J.; Baptista, J.; Madruga, J.; Lima, E. Biological activities of organic extracts and specialized metabolites from different parts of Cryptomeria japonica (Cupressaceae)—A critical review. Phytochemistry 2023, 206, 113520. [Google Scholar] [CrossRef] [PubMed]
- Li, W.H.; Chang, S.T.; Chang, S.C.; Chang, H.T. Isolation of antibacterial diterpenoids from Cryptomeria japonica bark. Nat. Prod. Res. 2008, 22, 1085–1093. [Google Scholar] [CrossRef] [PubMed]
- Morisawa, J.; Kim, C.S.; Kashiwagi, T.; Tebayashi, S.; Horiike, M. Repellents in the Japanese cedar, Cryptomeria japonica, against the pill-bug, Armadillidium vulgare. Biosci. Biotech. Bioch. 2002, 66, 2424–2428. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, Y.; Johuchi, T.; Saijo, H.; Takahashi, K.; Ashitani, T. Inhibition of the harmful alga Microcystis aeruginosa by sugi (Cryptomeria japonica) bark. J. Wood Sci. 2020, 66, 75. [Google Scholar] [CrossRef]
- Shyur, L.F.; Huang, C.C.; Lo, C.P.; Chiu, C.Y.; Chen, Y.P.; Wang, S.Y.; Chang, S.T. Hepatoprotective phytocompounds from Cryptomeria japonica are potent modulators of inflammatory mediators. Phytochemistry 2008, 69, 1348–1358. [Google Scholar] [CrossRef]
- Trianoski, R.; Iwakiri, S.; de Matos, J.L.M.; Chies, D. Use of Cryptomeria japonica wood for the production of particleboard. Sci. For. 2013, 41, 57–64. Available online: https://www.researchgate.net/publication/290554439 (accessed on 10 February 2023).
- Ota, Y.; Kimura, M.K.; Hattori, T.; Miyuki, Y.; Endo, R. First report of trunk rot caused by Fomitiporia torreyae in Kyoto prefecture on cultivars of Japanese cedar with no relatedness to ‘Sanbu-sugi’. J. For. Res. -Jpn. 2016, 21, 105–109. [Google Scholar] [CrossRef]
- Sawada, K. Fungi inhabiting on conifers in the Tohoku district. I. Fungi on “Sugi” (Cryptomeria japonica D. Don.). Bull. Gov. For. Exp. Stn. Meguro 1950, 45, 27–53. [Google Scholar]
- Zhou, L.; Li, J.; Chen, F.; Chen, J.; Ye, J. First report of shoot blight on Cryptomeria japonica caused by Pestalotiopsis neglecta in China. Plant Dis. 2019, 103, 2140–2141. [Google Scholar] [CrossRef]
- Crous, P.W.; Slippers, B.; Wingfield, M.J.; Rheeder, J.; Marasas, W.F.O.; Philips, A.J.L.; Alves, A.; Burgess, T.; Barber, P.; Groenewald, J.Z. Phylogenetic lineages in the Botryosphaeriaceae. Stud. Mycol. 2006, 55, 235–253. [Google Scholar] [CrossRef] [Green Version]
- Li, G.Q.; Liu, F.F.; Li, J.Q.; Liu, Q.L.; Chen, S.F. Botryosphaeriaceae from Eucalyptus plantations and adjacent plants in China. Persoonia 2018, 40, 63–95. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Li, S.J.; Han, S.; Qiao, T.M.; Zhou, Y.J.; Zhu, T.H. First report of Neofusicoccum parvum causing stem and branch blight disease of Zanthoxylum bungeanum in Sichuan, China. Plant Dis. 2021, 105, 3742. [Google Scholar] [CrossRef] [PubMed]
- Seddaiu, S.; Mello, A.; Sechi, C.; Cerboneschi, A.; Linaldeddu, B.T. First report of Neofusicoccum parvum associated with chestnut nut rot in Italy. Plant Dis. 2021, 105, 3743. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Villarroel-Zeballos, M.I.; Ficheux, P.F.; Zima, H.; Dhillon, B.D.S.; Correll, J.C. First report of Neofusicoccum parvum causing dieback and canker disease on hemp in the United States. Plant Dis. 2020, 104, 3075–3076. [Google Scholar] [CrossRef]
- Diniz, I.; Batista, D.; Pena, A.R.; Rodrigues, A.S.B.; Reis, P.; Baldé, A.; Indjai, B.; Catarino, L.; Monteiro, F. First report of dieback caused by Neofusicoccum batangarum in cashew in Guinea-Bissau. Plant Dis. 2021, 105, 1215. [Google Scholar] [CrossRef] [PubMed]
- Tapia, L.; Larach, A.; Riquelme, N.; Guajardo, J.; Besoain, X. First report of Neofusicoccum luteum causing stem-end rot disease on avocado fruits in Chile. Plant Dis. 2020, 104, 2027. [Google Scholar] [CrossRef]
- Wan, Y.; Wang, X.Y.; He, L.X.; Xu, W.; Zhu, L.H. First report of Neofusicoccum mangiferae causing shoot blight of Sassafras tzumu in China. Plant Dis. 2020, 104, 2028–2029. [Google Scholar] [CrossRef] [Green Version]
- Li, G.Q.; Slippers, B.; Wingfield, M.J.; Chen, S.F. Variation in Botryosphaeriaceae from Eucalyptus plantations in YunNan Province in southwestern China across a climatic gradient. IMA Fungus. 2020, 11, 22. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Groenewald, J.Z.; Lombard, L.; Schumacher, R.K.; Phillips, A.J.L.; Crous, P.W. Evaluating species in Botryosphaeriales. Persoonia 2021, 46, 63–115. [Google Scholar] [CrossRef] [PubMed]
- Brewer, M.T.; Cameron, C.J.; Chan, C.T.; Dutta, B.; Gitaitis, R.D.; Grauke, L.J.; Brock, J.H.; Brenneman, T.D. Neofusicoccum caryigenum, a new species causing leaf dieback disease of pecan (Carya illinoinensis). Mycologia 2021, 113, 586–598. [Google Scholar] [CrossRef]
- Xu, X.L.; Wang, F.H.; Liu, C.; Yang, H.B.; Zeng, Z.; Wang, B.X.; Liu, Y.G.; Yang, C.L. Morphology and phylogeny of ascomycetes associated with walnut trees (Juglans regia) in Sichuan province, China. Front. Microbiol. 2022, 13, 1016548. [Google Scholar] [CrossRef]
- Hattori, Y.; Ando, Y.; Nakashima, C. Taxonomical re-examination of the genus Neofusicoccum in Japan. Mycoscience 2021, 62, 250–259. [Google Scholar] [CrossRef]
- Tennakoon, D.S.; Kuo, C.H.; Maharachchikumbura, S.S.N.; Thambugala, K.M.; Gentekaki, E.; Phillips, A.J.L.; Bhat, D.J.; Wanasinghe, D.N.; de Silva, N.I.; Promputtha, I.; et al. Taxonomic and phylogenetic contributions to Celtis formosana, Ficus ampelas, F. septica, Macaranga tanarius and Morus australis leaf litter inhabiting microfungi. Fungal Divers. 2021, 108, 1–215. [Google Scholar] [CrossRef]
- Crous, P.W.; Hernández-Restrepo, M.; Schumacher, R.K.; Cowan, D.A.; Maggs-Kölling, G.; Marais, E.; Wingfield, M.J.; Yilmaz, N.; Adan, O.C.G.; Akulov, A.; et al. New and Interesting Fungi. 4. Fungal Syst. Evol. 2021, 7, 255–343. [Google Scholar] [CrossRef]
- Zheng, X.R.; Zhang, M.J.; Shang, X.L.; Fang, S.Z.; Chen, F.M. Stem Canker on Cyclocarya paliurus is Caused by Botryosphaeria dothidea. Plant Dis. 2020, 104, 1032–1040. [Google Scholar] [CrossRef]
- Phillips, A.J.L.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef] [Green Version]
- Smith, H.; Wingfield, M.J.; Crous, P.W.; Coutinho, T.A. Sphaeropsis sapinea and Botryosphaeria dothidea endophytic in Pinus spp. and Eucalyptus spp. in South Africa. S. Afr. J. Bot. 1996, 62, 86–88. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Lin, S.; He, W.; Zhang, Y. Three species of Neofusicoccum (Botryosphaeriaceae, Botryosphaeriales) associated with woody plants from southern China. Mycosphere 2017, 8, 797–808. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press, Inc.: New York, NY, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microb. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Pavlic, D.; Slippers, B.; Coutinho, T.A.; Wingfield, M.J. Multiple gene genealogies and phenotypic data reveal cryptic species of the Botryosphaeriaceae: A case study on the Neofusicoccum parvum/N. ribis complex. Mol. Phylogenet. Evol. 2009, 51, 259–268. [Google Scholar] [CrossRef]
- Sakalidis, M.L.; Hardy, G.E.S.; Burgess, T.I. Use of the Genealogical Sorting Index (GSI) to delineate species boundaries in the Neofusicoccum parvum-Neofusicoccum ribis species complex. Mol. Phylogenet. Evol. 2011, 60, 333–344. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Groenewald, J.Z.; Cheewangkoon, R.; Jami, F.; Abdollahzadeh, J.; Lombard, L.; Crous, P.W. Families, genera, and species of Botryosphaeriales. Fungal Biol. 2017, 124, 322–346. [Google Scholar] [CrossRef]
- Begoude, B.A.D.; Slippers, B.; Wingfield, M.J.; Roux, J. Botryosphaeriaceae associated with Terminalia catappa in Cameroon, South Africa and Madagascar. Mycol. Prog. 2010, 9, 101–123. [Google Scholar] [CrossRef] [Green Version]
- Marques, M.W.; Lima, N.B.; de Morais, M.A.J.; Michereff, S.J.; Phillips, A.J.L.; Camara, M.P.S. Botryosphaeria, Neofusicoccum, Neoscytalidium and Pseudofusicoccum species associated with mango in Brazil. Fungal Divers. 2013, 61, 195–208. [Google Scholar] [CrossRef]
- Slippers, B.; Fourie, G.; Crous, P.W.; Coutinho, T.A.; Wingfield, B.D.; Carnegie, A.J.; Wingfield, M.J. Speciation and distribution of Botryospaeria spp. on native and introduced Eucalyptus trees in Australia and South Africa. Stud. Mycol. 2005, 50, 343–358. [Google Scholar]
- Slippers, B.; Crous, P.W.; Denman, S.; Coutinho, T.A.; Wingfield, B.D.; Wingfield, M.J. Combined multiple gene genealogies and phenotypic characters differentiate several species previously identified as Botryosphaeria dothidea. Mycologia 2004, 96, 83–101. [Google Scholar] [CrossRef] [Green Version]
- Crous, P.W.; Groenewald, J.Z.; Shivas, R.G.; Edwards, J.; Seifert, K.A.; Alfenas, A.C.; Alfenas, R.F.; Burgess, T.I.; Carnegie, A.J.; Hardy, G.E.; et al. Fungal Planet description sheets: 69–91. Persoonia 2011, 26, 108–156. [Google Scholar] [CrossRef]
- Chen, S.F.; Li, G.; Liu, F.; Michailides, T.J. Novel species of Botryosphaeriaceae associated with shoot blight of pistachio. Mycologia 2015, 107, 780–792. [Google Scholar] [CrossRef]
- Osorio, J.A.; Crous, C.J.; de Beer, Z.W.; Wingfield, M.J.; Roux, J. Endophytic Botryosphaeriaceae, including five new species, associated with mangrove trees in South Africa. Fungal Biol. 2017, 121, 361–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgess, T.I.; Barber, P.; Hardy, G.E.S.J. Botryosphaeria spp. associated with eucalypts in Western Australia including description of Fusicoccum macroclavatum sp. nov. Australas. Plant Pathol. 2005, 34, 557–567. [Google Scholar] [CrossRef]
- Inderbitzin, P.; Bostock, R.M.; Trouillas, F.P.; Michailides, T.J. A six locus phylogeny reveals high species diversity in Botryosphaeriaceae from California almond. Mycologia 2010, 102, 1350–1368. [Google Scholar] [CrossRef] [PubMed]
- Taylor, K.; Barber, P.; Hardy, G.; Burgess, T.I. Botryosphaeriaceae from tuart (Eucalyptus gomphocephala) woodland, including descriptions of four new species. Mycol. Res. 2009, 113, 337–353. [Google Scholar] [CrossRef]
- Jami, F.; Marincowitz, S.; Slippers, B.; Wingfield, M.J. New Botryosphaeriales on native red milkwood (Mimusops caffra). Australas. Plant Pathol. 2018, 47, 475–484. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Gao, F.L.; Jakovlic, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Quaedvlieg, W.; Binder, M.; Groenewald, J.Z.; Summerell, B.A.; Carnegie, A.J.; Burgess, T.I.; Crous, P.W. Introducing the consolidated species concept to resolve species in the Teratosphaeriaceae. Persoonia 2014, 33, 1–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakalidis, M.L.; Slippers, B.; Wingfield, B.D.; Hardy, G.E.S.J.; Burgess, T.I. The challenge of understanding the origin, pathways and extent of fungal invasions: Global populations of the Neofusicoccum parvum-N. ribis species complex. Divers. Distrib. 2013, 19, 873–883. [Google Scholar] [CrossRef]
- Farr, D.F.; Rossman, A.Y. Fungal Databases, U.S. National Fungus Collections, ARS, USDA. Available online: https://nt.ars-grin.gov/fungaldatabases/ (accessed on 10 February 2023).
- Bellée, A.; Comont, G.; Nivault, A.; Abou-Mansour, E.; Coppin, C.; Dufour, M.C.; Corio-Costet, M.F. Life traits of four Botryosphaeriaceae species and molecular responses of different grapevine cultivars or hybrids. Plant Pathol. 2017, 66, 763–776. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.F.; Morgan, D.P.; Hasey, J.K.; Anderson, K.; Michailides, T.J. Phylogeny, morphology, distribution, and pathogenicity of Botryosphaeriaceae and Diaporthaceae from English walnut in California. Plant Dis. 2014, 98, 636–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitt, W.M.; Huang, R.; Steel, C.C.; Savocchia, S. Pathogenicity and epidemiology of Botryosphaeriaceae species isolated from grapevines in Australia. Australas Plant Path. 2013, 42, 573–582. [Google Scholar] [CrossRef]
- Yacoub, A.; Haidar, R.; Gerbore, J.; Masson, C.; Dufour, M.C.; Guyoneaud, R.; Rey, P. Pythium oligandrum induces grapevine defence mechanisms against the trunk pathogen Neofusicoccum parvum. Phytopathol. Mediterr. 2020, 59, 565–580. [Google Scholar] [CrossRef]
- Rooney-Latham, S.; Tidwell, T.E.; Blomquist, C.L.; Peek, K.S. First report of Neofusicoccum nonquaesitum causing branch cankers on giant sequoia (Sequoiadendron giganteum) in North America. Plant Dis. 2012, 96, 905. [Google Scholar] [CrossRef]
- Haenzi, M.; Cochard, B.; Chablais, R.; Crovadore, J.; Lefort, F. Neofusicoccum parvum, A new agent of Sequoia canker and dieback identified in Geneva, Switzerland. Forests 2021, 12, 434. [Google Scholar] [CrossRef]
- Perez, S.; Guerrero, J.; Galdames, R. First report of Neofusicoccum nonquaesitum in Chile causing branch dieback and decline in Araucaria araucana. Plant Dis. 2018, 102, 1460. [Google Scholar] [CrossRef]
- Luo, Z.L.; Bhat, D.J.; Jeewon, R.; Boonmee, S.; Bao, D.F.; Zhao, Y.C.; Chai, H.M.; Su, H.Y.; Su, X.J.; Hyde, K.D. Molecular phylogeny and morphological characterization of asexual fungi (Tubeufiaceae) from freshwater habitats in Yunnan, China. Cryptogam. Mycol. 2017, 38, 27–53. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, Y.; Sun, W.; Zhao, L.; Pavlic-Zupanc, D.; Crous, P.W.; Slippers, B.; Dai, Y. Toward a natural classification of Botryosphaeriaceae: A study of the type specimens of Botryosphaeria sensu lato. Front. Microbiol. 2021, 12, 737541. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GenBank Accession Number | ||||||
|---|---|---|---|---|---|---|
| Species | Isolate | ITS c | tef1 c | tub2 c | rpb2 c | Reference |
| Neofusicoccum arbuti | CBS 116131 a | AY819720 | KF531792 | KF531793 | KX464003 | [32] |
| N. arbuti | CBS 117090 | AY819724 | KF531791 | KF531794 | N/A | [32] |
| N. australe | CBS 110865 | AY343408 | KX464661 | KX464937 | KX464005 | [40] |
| N. australe | CMW 6837 a | AY339262 | AY339270 | AY339254 | EU339573 | [40] |
| N. batangarum | CBS 124923 | FJ900608 | FJ900654 | FJ900635 | FJ900616 | [41] |
| N. batangarum | CBS 124924 a | FJ900607 | FJ900653 | FJ900634 | FJ900615 | [41] |
| N. brasiliense | CMM 1285 | JX513628 | JX513608 | KC794030 | N/A | [42] |
| N. brasiliense | CMM 1338 a | JX513630 | JX513610 | KC794031 | N/A | [42] |
| N. buxi | CBS 113714 | KX464164 | KX464677 | KX464954 | KX464009 | [40] |
| N. buxi | CBS 116.75 a | KX464165 | KX464678 | N/A | KX464010 | [40] |
| N.caryigenum | CBS 146964 a | MW405114 | MW393657 | MW393679 | MW393668 | [26] |
| N. cordaticola | CBS 123634 a | EU821898 | EU821868 | EU821838 | EU821928 | [38] |
| N. cordaticola | CBS 123635 | EU821903 | EU821873 | EU821843 | EU821933 | [38] |
| N. cryptoaustrale | CMW 23785 a | FJ752742 | FJ752713 | FJ752756 | KX464014 | [40] |
| N. cryptomeriae | G24 = CFCC 55721 a,b | ON209700 | OP056461 | OP056458 | OP056455 | this study |
| N. cryptomeriae | G1 = CFCC 55720 b | ON209698 | OP056459 | OP056456 | OP056453 | this study |
| N. cryptomeriae | G2 = CFCC 55728 b | ON209699 | OP056460 | OP056457 | OP056454 | this study |
| N. eucalypticola | CBS 115679 a | AY615141 | AY615133 | AY615125 | N/A | [43] |
| N. eucalypticola | CBS 115766 | AY615143 | AY615135 | AY615127 | N/A | [43] |
| N. eucalyptorum | CBS 115791 a | AF283686 | AY236891 | AY236920 | N/A | [44] |
| N. eucalyptorum | CMW 10126 | AF283687 | AY236892 | AY236921 | N/A | [44] |
| N. grevilleae | CBS 129518 a | JF951137 | N/A | N/A | N/A | [45] |
| N. hellenicum | CERC1947 a | KP217053 | KP217061 | KP217069 | N/A | [46] |
| N. hellenicum | CERC1948 | KP217054 | KP217062 | KP217070 | N/A | [46] |
| N. hongkongense | CERC2968 | KX278051 | KX278156 | KX278260 | KX278282 | [17] |
| N. hongkongense | CERC2973 a | KX278052 | KX278157 | KX278261 | KX278283 | [17] |
| N. hyperici | MUCC 241 a | LC589125 | LC589137 | LC589147 | LC589160 | [28] |
| N. hyperici | MUCC 2509 | LC589126 | LC589138 | LC589148 | LC589161 | [28] |
| N. illicii | CGMCC 3.18310 a | KY350149 | N/A | KY350155 | N/A | [34] |
| N. illicii | CGMCC 3.18311 | KY350150 | KY817756 | KY350156 | N/A | [34] |
| N. kwambonambiense | CBS 123639 | EU821900 | EU821870 | EU821840 | EU821930 | [38] |
| N. kwambonambiense | CBS 123641 | EU821919 | EU821889 | EU821859 | EU821949 | [38] |
| N. lumnitzerae | CMW 41228 | KP860882 | KP860725 | KP860803 | KU587926 | [47] |
| N. lumnitzerae | CMW 41469 a | KP860881 | KP860724 | KP860801 | KU587925 | [47] |
| N. luteum | CBS 562.92 a | KX464170 | KX464690 | KX464968 | KX464020 | [40] |
| N. macroclavatum | CBS 118223 a | DQ093196 | DQ093217 | DQ093206 | KX464022 | [48] |
| N. magniconidium | CSF5875 | MT028611 | MT028777 | MT028943 | MT029084 | [24] |
| N. magniconidium | CSF5876 a | MT028612 | MT028778 | MT028944 | MT029085 | [24] |
| N. mangiferae | CBS 118531 a | AY615185 | DQ093221 | AY615172 | N/A | [48] |
| N. mangiferae | CBS 118532 | AY615186 | DQ093220 | AY615173 | KX464023 | [48] |
| N. mediterraneum | CBS 121718 a | GU251176 | GU251308 | GU251836 | KX464024 | [49] |
| N. microconidium | CERC3497 a | KX278053 | KX278158 | KX278262 | MF410203 | [17] |
| N. microconidium | CERC3498 | KX278054 | KX278159 | KX278263 | MF410204 | [17] |
| N. miyakoense | MUCC 2585 a | N/A | LC589146 | LC589157 | LC589170 | [28] |
| N. miyakoense | MUCC 2586 | LC589133 | LC589144 | LC589155 | LC589168 | [28] |
| N. moracearium | MELU 19-2758 a | NR174834 | MW183808 | N/A | N/A | [29] |
| N. moracearium | MFLU 19-0316 | MW063187 | MW183809 | N/A | N/A | [29] |
| N. mystacidii | CBS 147079 a | NR173012 | MW890094 | MW890133 | MW890065 | [30] |
| N. ningerense | CSF6028 | MT028613 | MT028779 | MT028945 | MT029086 | [24] |
| N. ningerense | CSF6030 | MT028614 | MT028780 | MT028946 | MT029087 | [24] |
| N. nonquaesitum | CBS 126655 a | GU251163 | GU251295 | GU251823 | KX464025 | [49] |
| N. nonquaesitum | PD 301 | GU251164 | GU251296 | GU251824 | N/A | [49] |
| N. occulatum | CBS 128008 a | EU301030 | EU339509 | EU339472 | EU339558 | [39] |
| N. occulatum | MUCC 286 | EU736947 | EU339511 | EU339474 | EU339560 | [39] |
| N. okinawaense | MUCC 789 a | LC589134 | LC589145 | LC589156 | LC589169 | [28] |
| N. parviconidium | CSF5667 a | MT028615 | MT028781 | MT028947 | MT029088 | [24] |
| N. parviconidium | CSF5677 | MT028619 | MT028785 | MT028951 | MT029092 | [24] |
| N. parvum | ATCC 58191 a | AY236943 | AY236888 | AY236917 | EU821963 | [44] |
| N. parvum | CMW 9080 | AY236942 | AY236887 | AY236916 | EU821962 | [44] |
| N. parvum | G15 = CFCC 55724 b | ON209685 | OP095379 | OP095383 | OP095387 | this study |
| N. parvum | G16 = CFCC 55718 b | ON209686 | OP095380 | OP095384 | OP095388 | this study |
| N. parvum | G91 = CFCC 55719 b | ON209687 | OP095381 | OP095385 | OP095389 | this study |
| N. parvum | G92 = CFCC 55723 b | ON209688 | OP095382 | OP095386 | OP095390 | this study |
| N. pennatisporum | WAC 13153 a | NR136987 | EF591976 | EF591959 | N/A | [50] |
| N. pistaciae | CBS 595.76 a | KX464163 | KX464676 | KX464953 | KX464008 | [40] |
| N. podocarpi | CBS 115065 | MT587507 | MT592222 | MT592714 | MT592411 | [25] |
| N. podocarpi | CBS 131677 a | MT587508 | MT592223 | MT592715 | MT592412 | [25] |
| N. protearum | CBS 114176 a | AF452539 | KX464720 | KX465006 | KX464029 | [40] |
| N. rapaneae | CBS 145973 a | MT587511 | MT592226 | MT592718 | MT592415 | [25] |
| N. rapaneae | CPC 32578 | MT587512 | MT592227 | MT592719 | MT592416 | [25] |
| N. ribis | CBS 114306 | MT587514 | MT592229 | MT592721 | MT592418 | [25] |
| N. ribis | CBS 115475 a | AY236935 | AY236877 | AY236906 | EU821958 | [25] |
| N. sichuanense | SICAUCC22-0099 a | OP058990 | OP066336 | OP066363 | OP066355 | [27] |
| N. sichuanense | SICAUCC22-0093 | OP058984 | OP066333 | OP066357 | OP066349 | [27] |
| N. sinense | CGMCC 3.18315 a | KY350148 | KY817755 | KY350154 | N/A | [34] |
| N. sinoeucalypti | CERC2005 a | KX278061 | KX278166 | KX278270 | KX278290 | [17] |
| N. sinoeucalypti | CERC3416 | KX278064 | KX278169 | KX278273 | KX278293 | [17] |
| N. stellenboschiana | CBS 110864 a | AY343407 | AY343348 | KX465047 | KX464042 | [40] |
| N. terminaliae | CBS 125263 a | GQ471802 | GQ471780 | KX465052 | KX464045 | [40] |
| N. terminaliae | CBS 125264 | GQ471804 | GQ471782 | KX465053 | KX464046 | [40] |
| N. umdonicola | CBS 123645 a | EU821904 | EU821874 | EU821844 | EU821934 | [40] |
| N. umdonicola | CBS 123646 | EU821905 | EU821875 | EU821845 | EU821935 | [40] |
| N. ursorum | CMW 23790 | FJ752745 | FJ752708 | KX465057 | N/A | [40] |
| N. ursorum | CMW 24480 a | FJ752746 | FJ752709 | KX465056 | KX464047 | [40] |
| N. variabile | CMW 37739 a | MH558608 | N/A | MH569153 | N/A | [51] |
| N. variabile | CMW 37742 | MH558609 | MH576585 | MH569154 | N/A | [51] |
| N. viticlavatum | CBS 112878 a | AY343381 | AY343342 | KX465058 | KX464048 | [32] |
| N. viticlavatum | CBS 112977 | AY343380 | AY343341 | KX465059 | N/A | [32] |
| N. vitifusiforme | CBS 110880 | AY343382 | AY343344 | KX465008 | N/A | [40] |
| N. vitifusiforme | CBS 110887 | AY343383 | AY343343 | KX465061 | KX464049 | [40] |
| N. yunnanense | CSF6034 | MT028672 | MT028838 | MT029004 | MT029117 | [24] |
| N. yunnanense | CSF6142 a | MT028667 | MT028833 | MT028999 | MT029112 | [24] |
| Botryosphaeria dothidea | CBS 115476 a | AY236949 | AY236898 | AY236927 | EU339577 | [44] |
| Species | Isolates | Conidiogenous Cells (μm) | Conidia (μm) | Reference |
|---|---|---|---|---|
| Neofusicoccum cryptomeriae | G1 = CFCC 55720 | 16.4–24.1 × 3–3.9 | 20.5–25.7 × 7.1–8.9 | this study |
| G2 = CFCC 55728 | 15.8–24.2 × 3.2–4 | 20.1–22.6 × 6.8–8 | this study | |
| G24 = CFCC 55721 | 13.8–22.4 × 3.4–4.2 | 23–26.1 × 7–7.8 | this study | |
| N. parvum | G15 = CFCC 55724 | 13.4–18.4 ×2.6–3.8 | 17–18.7 × 5.8–6.3 | this study |
| G16 = CFCC 55718 | 11.4–19.5 × 2.1–3.6 | 16.8–18.1 × 5.7–6.3 | this study | |
| G91 = CFCC 55719 | 13.7–20.9 × 2.5–3.3 | 17.1–19.5 × 5.7–6.4 | this study | |
| G92 = CFCC 55723 | 12.2–21.4 × 2.6–3.4 | 17.1–18.8 × 5.6–6.3 | this study | |
| N. sinense | CGMCC 3.18315 | not observed | 17.6–20.4 × 7.4–8 | [34] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Si, Y.-Z.; Sun, J.-W.; Wan, Y.; Chen, Y.-N.; He, J.; Li, W.-Z.; Li, D.-W.; Zhu, L.-H. Neofusicoccum cryptomeriae sp. nov. and N. parvum Cause Stem Basal Canker of Cryptomeria japonica in China. J. Fungi 2023, 9, 404. https://doi.org/10.3390/jof9040404
Si Y-Z, Sun J-W, Wan Y, Chen Y-N, He J, Li W-Z, Li D-W, Zhu L-H. Neofusicoccum cryptomeriae sp. nov. and N. parvum Cause Stem Basal Canker of Cryptomeria japonica in China. Journal of Fungi. 2023; 9(4):404. https://doi.org/10.3390/jof9040404
Chicago/Turabian StyleSi, Yuan-Zhi, Jian-Wei Sun, Yu Wan, Yi-Na Chen, Jiao He, Wei-Zheng Li, De-Wei Li, and Li-Hua Zhu. 2023. "Neofusicoccum cryptomeriae sp. nov. and N. parvum Cause Stem Basal Canker of Cryptomeria japonica in China" Journal of Fungi 9, no. 4: 404. https://doi.org/10.3390/jof9040404
APA StyleSi, Y. -Z., Sun, J. -W., Wan, Y., Chen, Y. -N., He, J., Li, W. -Z., Li, D. -W., & Zhu, L. -H. (2023). Neofusicoccum cryptomeriae sp. nov. and N. parvum Cause Stem Basal Canker of Cryptomeria japonica in China. Journal of Fungi, 9(4), 404. https://doi.org/10.3390/jof9040404