
An Integrative Study of Mycobiome in Different Habitats from a High Arctic Region: Diversity, Distribution, and Functional Role


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Area
2.2. DNA Extraction, PCR Amplification, and Illumina Sequencing
2.3. Sequencing Data Processing
2.4. Statistical Analyses
3. Results
3.1. Sequence Data
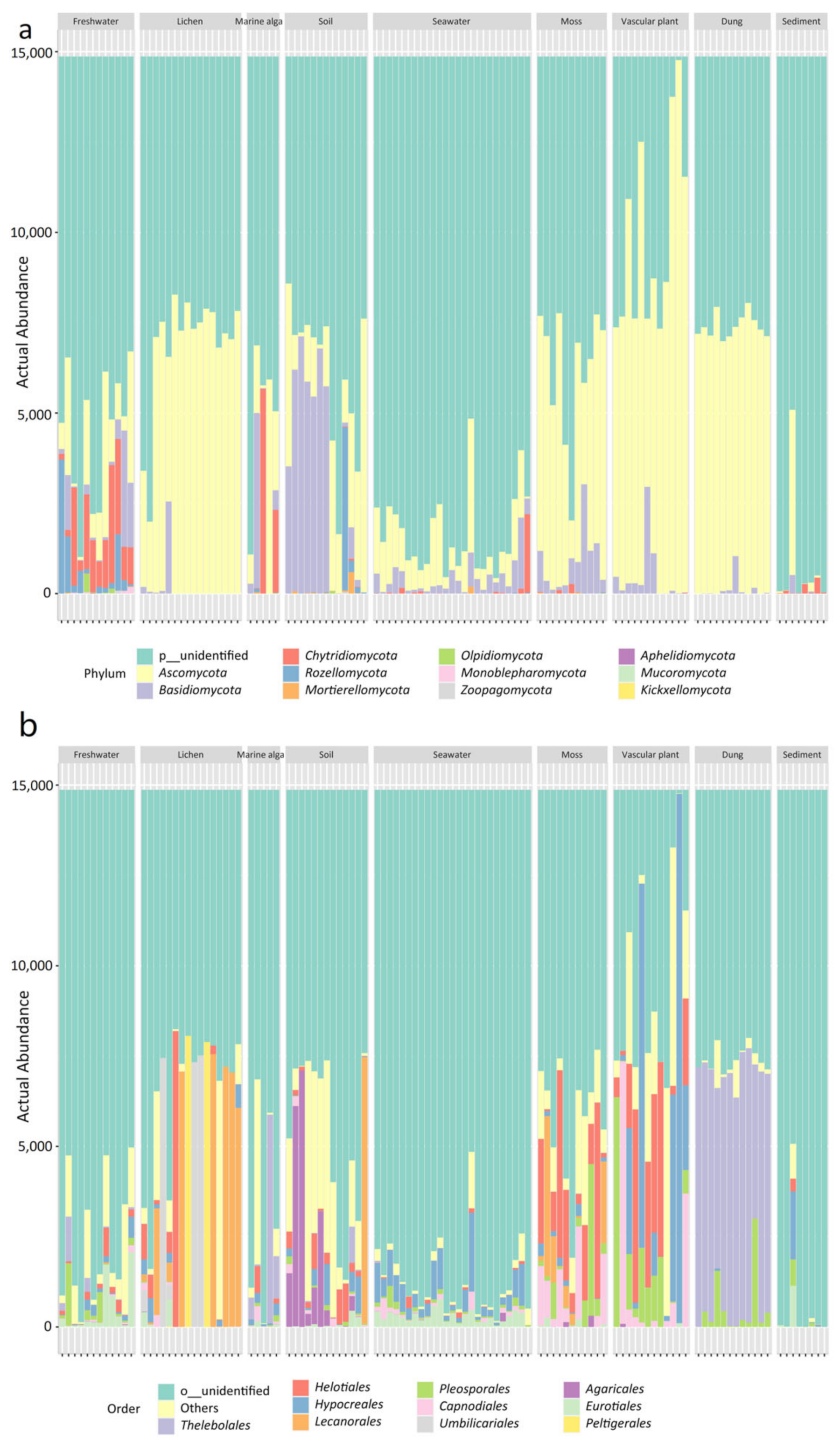
3.2. Diversity and Composition of the Mycobiome in the Nine Habitats
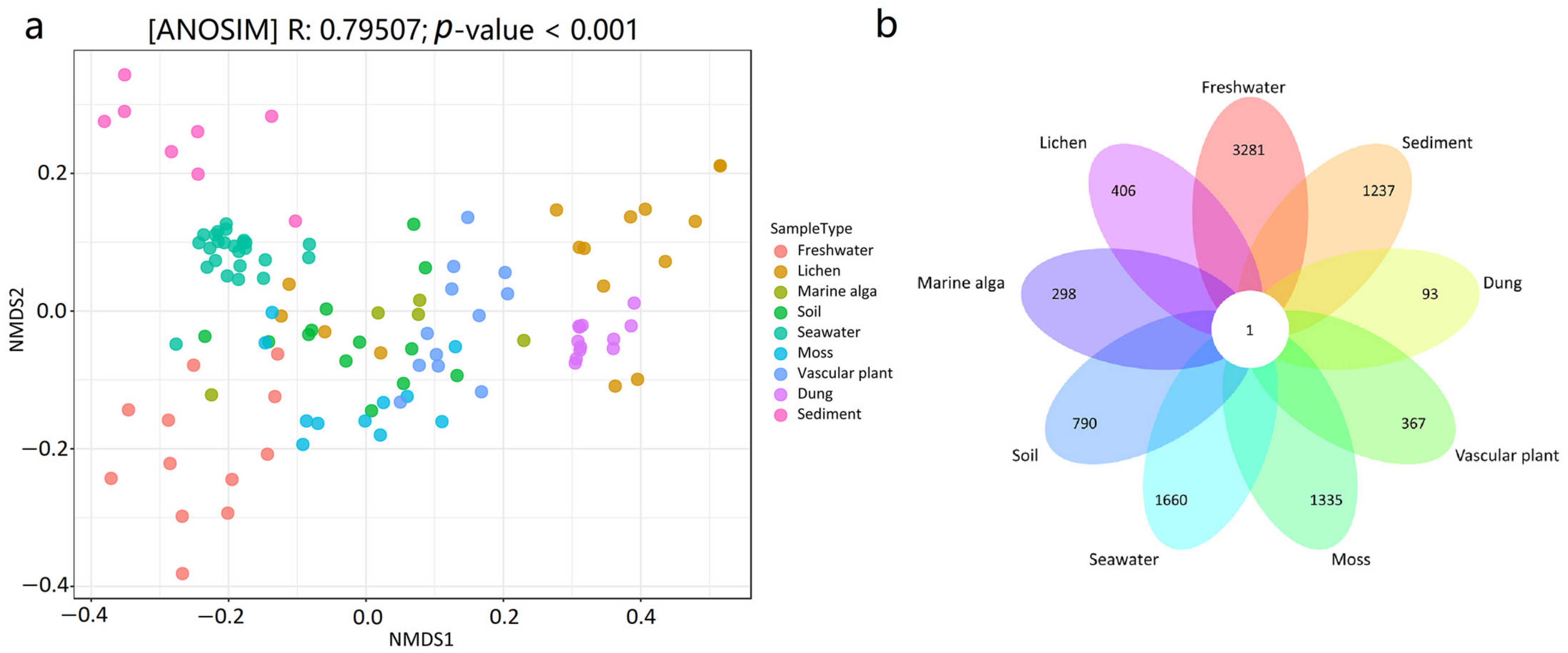
3.3. Dissimilarity of the Mycobiome among the Nine Habitats
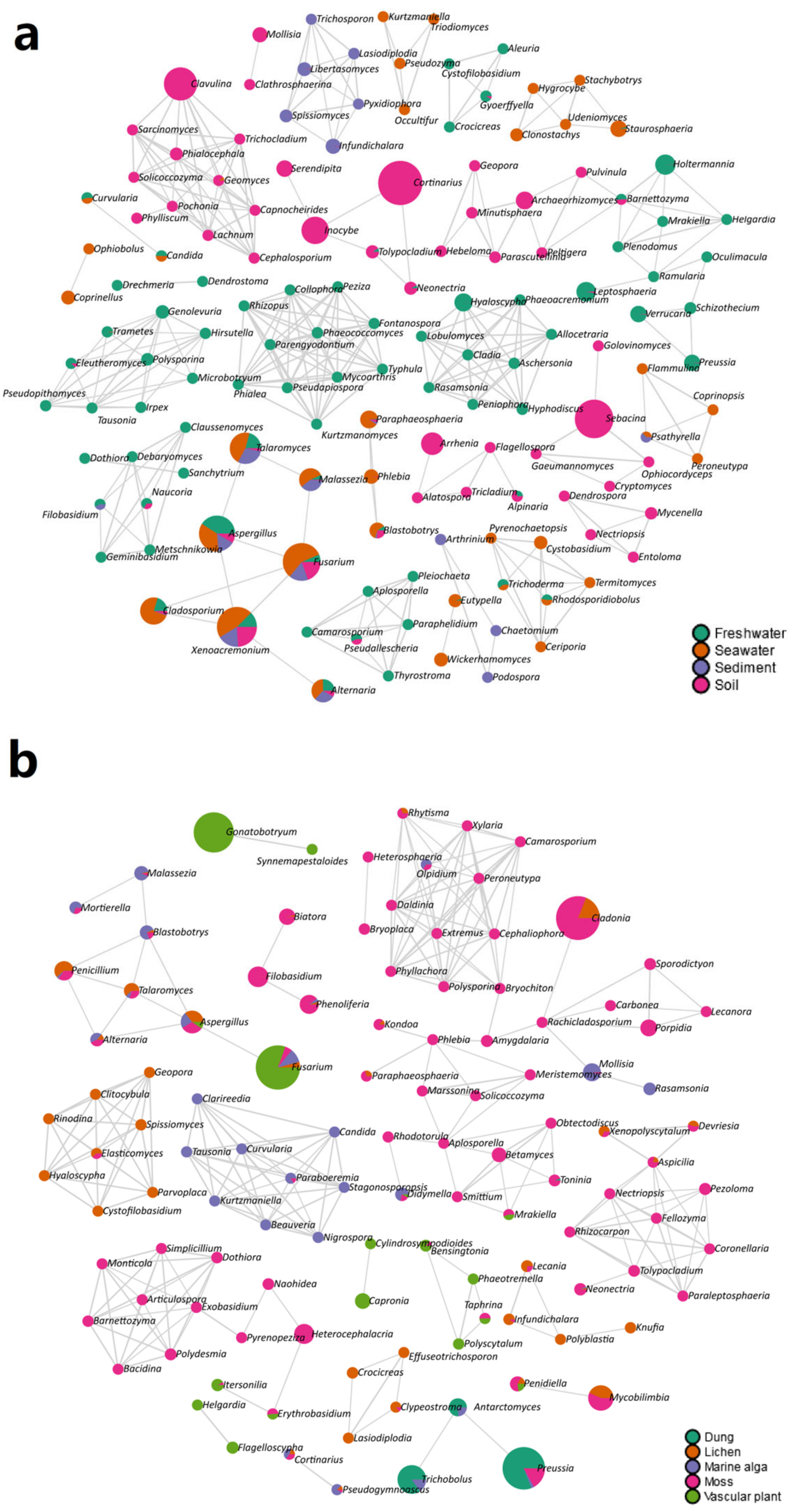
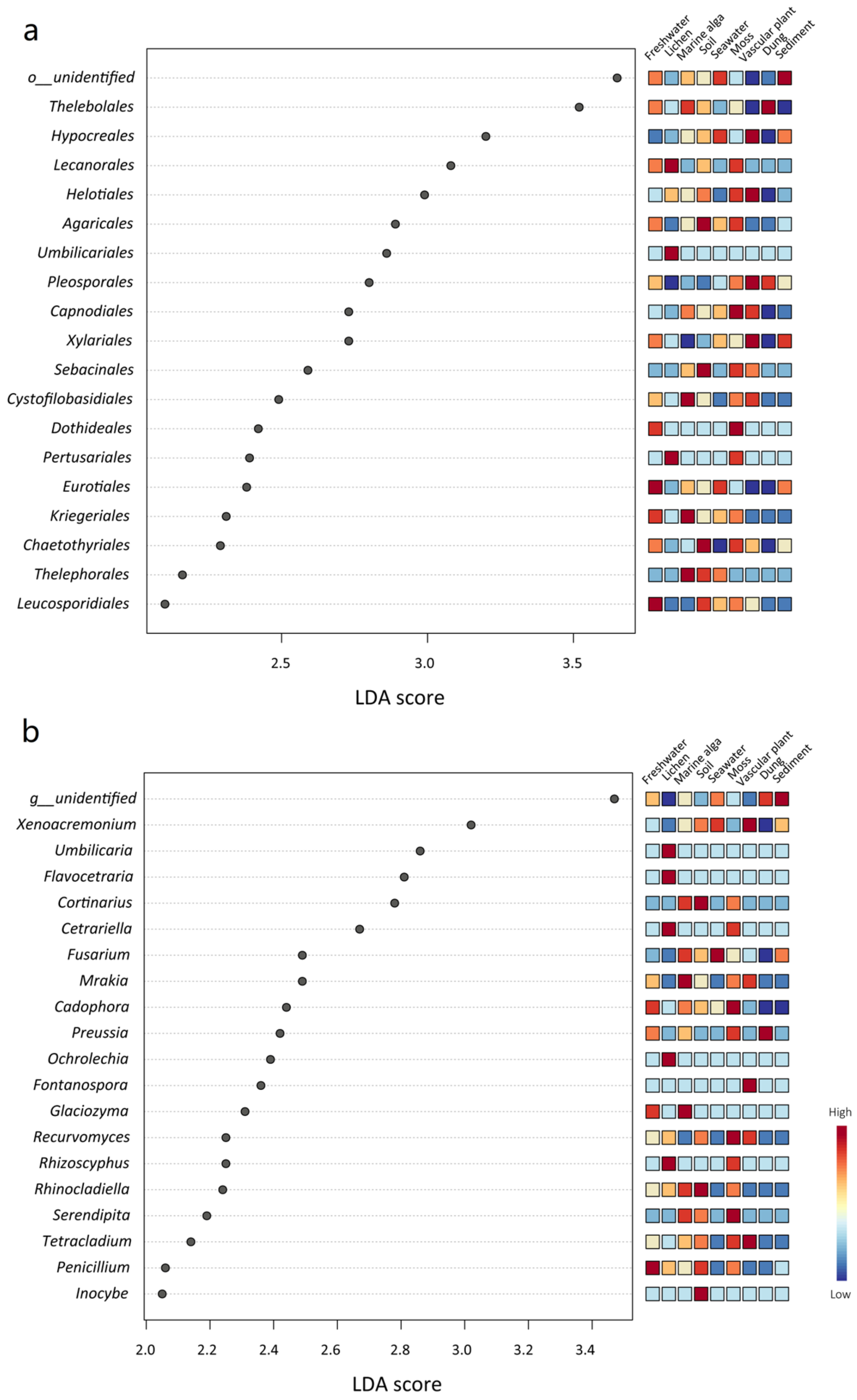
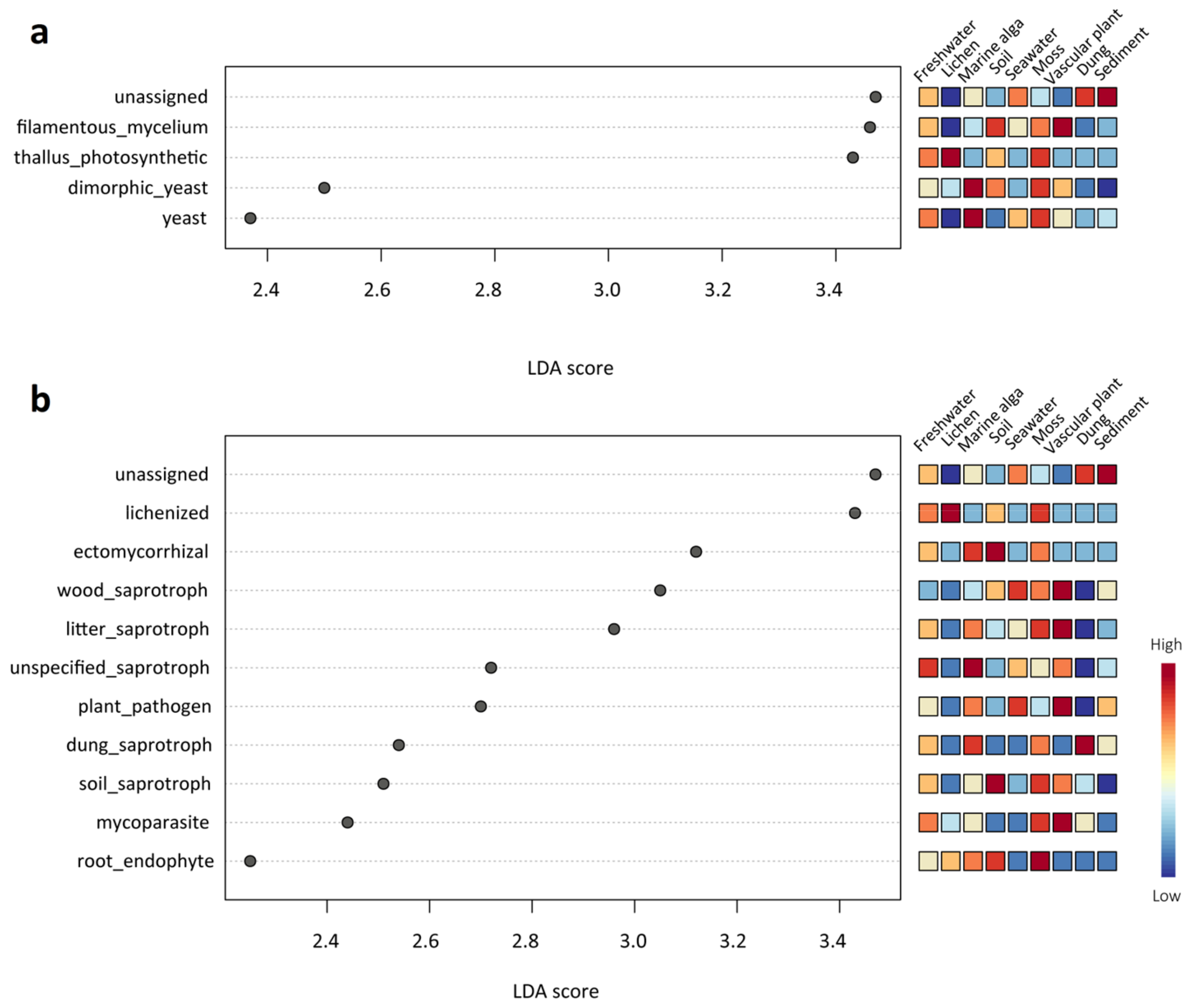
3.4. Ecological Role of the Mycobiome in the Nine Habitats
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Biskaborn, B.K.; Smith, S.L.; Noetzli, J.; Matthes, H.; Vieira, G.; Streletskiy, D.A.; Schoeneich, P.; Romanovsky, V.E.; Lewkowicz, A.G.; Abramov, A.; et al. Permafrost is warming at a global scale. Nat. Commun. 2019, 10, 264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassett, B.T.; Gradinger, R. Chytrids dominate arctic marine fungal communities. Environ. Microbiol. 2016, 18, 2001–2009. [Google Scholar] [CrossRef] [PubMed]
- Tape, K.E.N.; Sturm, M.; Racine, C. The evidence for shrub expansion in Northern Alaska and the Pan-Arctic. Glob. Chang. Biol. 2006, 12, 686–702. [Google Scholar] [CrossRef]
- Deslippe, J.R.; Hartmann, M.; Simard, S.W.; Mohn, W.W. Long-term warming alters the composition of Arctic soil microbial communities. FEMS Microbiol. Ecol. 2012, 82, 303–315. [Google Scholar] [CrossRef] [Green Version]
- Morgado, L.N.; Semenova, T.A.; Welker, J.M.; Walker, M.D.; Smets, E.; Geml, J. Summer temperature increase has distinct effects on the ectomycorrhizal fungal communities of moist tussock and dry tundra in Arctic Alaska. Glob. Chang. Biol. 2015, 21, 959–972. [Google Scholar] [CrossRef] [Green Version]
- Timling, I.; Taylor, D.L. Peeking through a frosty window: Molecular insights into the ecology of Arctic soil fungi. Fungal Ecol. 2012, 5, 419–429. [Google Scholar] [CrossRef]
- Blaud, A.; Lerch, T.Z.; Phoenix, G.K.; Osborn, A.M. Arctic soil microbial diversity in a changing world. Res. Microbiol. 2015, 166, 796–813. [Google Scholar] [CrossRef]
- Hafizah, S.; Alias, S.; Siang, H.; Smykla, J.; Pang, K.-L.; Guo, S.-Y.; Convey, P. Studies on diversity of soil microfungi in the Hornsund area, Spitsbergen. Pol. Polar Res. 2013, 34, 39–54. [Google Scholar] [CrossRef]
- Singh, S.; Singh, S.; Yadav, L.S.; Singh, P.; Ravindra, R. Filamentous Soil Fungi from Ny-Ålesund, Spitsbergen, and Screening for Extracellular Enzymes. ARCTIC 2012, 65, 45–55. [Google Scholar] [CrossRef]
- Zhang, T.; Wei, X.L.; Wei, Y.Z.; Liu, H.Y.; Yu, L.Y. Diversity and distribution of cultured endolichenic fungi in the Ny-Alesund Region, Svalbard (High Arctic). Extremophiles 2016, 20, 461–470. [Google Scholar] [CrossRef]
- Zhang, T.; Zhao, L.; Yu, C.; Wei, T.; Yu, L. Diversity and bioactivity of cultured aquatic fungi from the High Arctic region. Adv. Polar Sci. 2017, 28, 29–42. [Google Scholar]
- Borzęcka, J.; Suchodolski, J.; Dudek, B.; Matyaszczyk, L.; Spychała, K.; Ogórek, R. The first comprehensive biodiversity study of culturable fungal communities inhabiting cryoconite holes in Werenskiold Glacier on Spitsbergen (Svalbard Archipelago, Arctic). Biology 2022, 11, 1224. [Google Scholar] [CrossRef]
- Grau, O.; Geml, J.; Pérez-Haase, A.; Ninot, J.M.; Semenova-Nelsen, T.A.; Peñuelas, J. Abrupt changes in the composition and function of fungal communities along an environmental gradient in the high Arctic. Mol. Ecol. 2017, 26, 4798–4810. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Wang, N.; Yu, L. Soil fungal community composition differs significantly among the Antarctic, Arctic, and Tibetan Plateau. Extremophiles 2020, 24, 821–829. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, N.F.; Liu, H.Y.; Zhang, Y.Q.; Yu, L.Y. Soil pH is a Key Determinant of Soil Fungal Community Composition in the Ny-Alesund Region, Svalbard (High Arctic). Front. Microbiol. 2016, 7, 227. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, N.F.; Yu, L.Y. Geographic Distance and Habitat Type Influence Fungal Communities in the Arctic and Antarctic Sites. Microb. Ecol. 2021, 82, 224–232. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, N.F.; Zhang, Y.Q.; Liu, H.Y.; Yu, L.Y. Diversity and distribution of fungal communities in the marine sediments of Kongsfjorden, Svalbard (High Arctic). Sci. Rep. 2015, 5, 14524. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Wei, X.L.; Zhang, Y.Q.; Liu, H.Y.; Yu, L.Y. Diversity and distribution of lichen-associated fungi in the Ny-Alesund Region (Svalbard, High Arctic) as revealed by 454 pyrosequencing. Sci. Rep. 2015, 5, 14850. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Wang, N.; Yu, L. Host-specificity of moss-associated fungal communities in the Ny-Ålesund region (Svalbard, High Arctic) as revealed by amplicon pyrosequencing. Fungal Ecol. 2021, 53, 101092. [Google Scholar] [CrossRef]
- Blaalid, R.; Davey, M.L.; Kauserud, H.; Carlsen, T.; Halvorsen, R.; Hoiland, K.; Eidesen, P.B. Arctic root-associated fungal community composition reflects environmental filtering. Mol. Ecol. 2014, 23, 649–659. [Google Scholar] [CrossRef]
- Botnen, S.S.; Thoen, E.; Eidesen, P.B.; Krabberod, A.K.; Kauserud, H. Community composition of arctic root-associated fungi mirrors host plant phylogeny. FEMS Microbiol. Ecol. 2020, 96, fiaa185. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Yao, Y.F. Endophytic Fungal Communities Associated with Vascular Plants in the High Arctic Zone Are Highly Diverse and Host-Plant Specific. PLoS ONE 2015, 10, e0130051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassett, B.T.; Ducluzeau, A.L.; Collins, R.E.; Gradinger, R. Spatial distribution of aquatic marine fungi across the western Arctic and sub-arctic. Environ. Microbiol. 2017, 19, 475–484. [Google Scholar] [CrossRef]
- Rämä, T.; Hassett, B.T.; Bubnova, E. Arctic marine fungi: From filaments and flagella to operational taxonomic units and beyond. Bot. Mar. 2017, 60, 433–452. [Google Scholar] [CrossRef]
- Kluge, M.; Wauthy, M.; Clemmensen, K.E.; Wurzbacher, C.; Hawkes, J.A.; Einarsdottir, K.; Rautio, M.; Stenlid, J.; Peura, S. Declining fungal diversity in Arctic freshwaters along a permafrost thaw gradient. Glob. Chang. Biol. 2021, 27, 5889–5906. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, N.F.; Zhang, Y.Q.; Liu, H.Y.; Yu, L.Y. Diversity and Distribution of Aquatic Fungal Communities in the Ny-Alesund Region, Svalbard (High Arctic): Aquatic Fungi in the Arctic. Microb. Ecol. 2016, 71, 543–554. [Google Scholar] [CrossRef]
- Mundra, S.; Halvorsen, R.; Kauserud, H.; Bahram, M.; Tedersoo, L.; Elberling, B.; Cooper, E.J.; Eidesen, P.B. Ectomycorrhizal and saprotrophic fungi respond differently to long-term experimentally increased snow depth in the High Arctic. Microbiologyopen 2016, 5, 856–869. [Google Scholar] [CrossRef]
- Canini, F.; Zucconi, L.; Pacelli, C.; Selbmann, L.; Onofri, S.; Geml, J. Vegetation, pH and Water Content as Main Factors for Shaping Fungal Richness, Community Composition and Functional Guilds Distribution in Soils of Western Greenland. Front. Microbiol. 2019, 10, 2348. [Google Scholar] [CrossRef] [Green Version]
- Geml, J.; Semenova, T.A.; Morgado, L.N.; Welker, J.M. Changes in composition and abundance of functional groups of arctic fungi in response to long-term summer warming. Biol. Lett. 2016, 12, 20160503. [Google Scholar] [CrossRef] [Green Version]
- Hassett, B.T.; Borrego, E.J.; Vonnahme, T.R.; Rama, T.; Kolomiets, M.V.; Gradinger, R. Arctic marine fungi: Biomass, functional genes, and putative ecological roles. ISME J. 2019, 13, 1484–1496. [Google Scholar] [CrossRef] [Green Version]
- Perini, L.; Mogrovejo, D.C.; Tomazin, R.; Gostincar, C.; Brill, F.H.H.; Gunde-Cimerman, N. Phenotypes Associated with Pathogenicity: Their Expression in Arctic Fungal Isolates. Microorganisms 2019, 7, 600. [Google Scholar] [CrossRef] [Green Version]
- Maturilli, M.; Herber, A.; König-Langlo, G. Climatology and time series of surface meteorology in Ny-Ålesund, Svalbard. Earth Syst. Sci. Data 2013, 5, 155–163. [Google Scholar] [CrossRef] [Green Version]
- Gardes, M.; Bruns, T.D. ITS Primers with enhanced specificity for Basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phy-logenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahe, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [Green Version]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2′s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef]
- Abarenkov, K.; Zirk, A.; Piirmann, T.; Pöhönen, R.; Ivanov, F.; Nilsson, R.H.; Kõljalg, U. UNITE QIIME release for Fungi 2. Version 10.05.2021. UNITE Community 2021, 7, 1264763. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. MicrobiomeAnalyst: A web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017, 45, W180–W188. [Google Scholar] [CrossRef] [PubMed]
- Põlme, S.; Abarenkov, K.; Nilsson, R.H.; Lindahl, B.; Clemmensen, K.; Kauserud, H.; Nguyen, N.; Kjøller, R.; Bates, S.; Baldrian, P.; et al. FungalTraits: A user-friendly traits database of fungi and fungus-like stramenopiles. Fungal Divers. 2020, 105, 1–16. [Google Scholar] [CrossRef]
- De Hoog, G.S.; Guarro, J.; Gené, J.; Ahmed, S.A.; Al-Hatmi, A.M.S.; Figueras, M.J.; Vitale, R.G. Atlas of Clinical Fungi, 4th ed.; CBS: Utrecht, The Netherlands, 2020. [Google Scholar]
- Walker, J.; Aldrich-Wolfe, L.; Riffel, A.; Barbare, H.; Simpson, N.; Trowbridge, J.; Jumpponen, A. Diverse Helotiales associated with the roots of three species of Arctic Ericaceae provide no evidence for host specificity. New Phytol. 2011, 191, 515–527. [Google Scholar] [CrossRef] [PubMed]
- He, C.Y.; Feng, J.J.; Zhang, T.; Yu, L.Y. Species diversity and floristic analysis of macrofungi in Ny-Ålesund Region, High Arctic. Chin. J. Polar Res. 2022, 34, 139–148. [Google Scholar] [CrossRef]
- Baldrian, P.; Valaskova, V. Degradation of cellulose by basidiomycetous fungi. FEMS Microbiol. Rev. 2008, 32, 501–521. [Google Scholar] [CrossRef] [Green Version]
- Geml, J.; Timling, I.; Robinson, C.; Lennon, N.; Nusbaum, C.; Brochmann, C.; Noordeloos, M.; Taylor, D.L. An arctic community of symbiotic fungi assembled by long-distance dispersers: Phylogenetic diversity of ectomycorrhizal basidiomycetes in Svalbard based on soil and sporocarp DNA. J. Biogeogr. 2011, 39, 74–88. [Google Scholar] [CrossRef]
- Prasad, R.; Chhabra, S.; Gill, S.; Kumar Singh, P.; Tuteja, N. The Microbial Symbionts: Potential for Crop Improvement in Changing Environments. In Advancement in Crop Improvement Techniques; Tuteja, N., Tuteja, R., Passricha, N., Saifi, S.K., Eds.; Woodhead Publishing: London, UK, 2020; pp. 233–240. [Google Scholar] [CrossRef]
- Pusz, W.; Urbaniak, J. Airborne fungi in Longyearbyen area (Svalbard, Norway)—Case study. Environ. Monit. Assess. 2021, 193, 290. [Google Scholar] [CrossRef]
- Wu, R.; Trubl, G.; Taş, N.; Jansson, J.K. Permafrost as a potential pathogen reservoir. One Earth 2022, 5, 351–360. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habitat Type | Soil | Freshwater | Seawater | Lichen | Sediment | Dung | Marine alga | Moss | Vascular Plant |
|---|---|---|---|---|---|---|---|---|---|
| Soil | - | 0.67664 *** | 0.84645 *** | 0.48733 *** | 0.97115 *** | 0.97516 *** | 0.47797 ** | 0.37873 *** | 0.58004 *** |
| Freshwater | 0.67664 *** | - | 0.9078 *** | 0.77907 *** | 0.97385 *** | 1 *** | 0.76711 ** | 0.65965 *** | 0.94234 *** |
| Seawater | 0.84645 *** | 0.9078 *** | - | 0.84193 *** | 0.90845 *** | 1 *** | 0.94132 *** | 0.92049 *** | 0.99311 *** |
| Lichen | 0.48733 *** | 0.77907 *** | 0.84193 *** | - | 0.82897 *** | 0.40703 *** | 0.43981 ** | 0.50916 *** | 0.48796 *** |
| Sediment | 0.97115 *** | 0.97385 *** | 0.90845 *** | 0.82897 *** | - | 1 *** | 0.94868 *** | 0.96906 *** | 1 *** |
| Dung | 0.97516 *** | 1 *** | 1 *** | 0.40703 *** | 1 *** | - | 0.90614 ** | 0.95968 *** | 0.92961 *** |
| Marine alga | 0.47797 ** | 0.76711 ** | 0.94132 *** | 0.43981 ** | 0.94868 *** | 0.90614 ** | - | 0.61455 *** | 0.42719 ** |
| Moss | 0.37873 *** | 0.65965 *** | 0.92049 *** | 0.50916 *** | 0.96906 *** | 0.95968 *** | 0.61455 *** | - | 0.60594 *** |
| Vascular plant | 0.58004 *** | 0.94234 *** | 0.99311 *** | 0.48796 *** | 1 *** | 0.92961 *** | 0.42719 ** | 0.60594 *** | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Yan, D.; Yu, L.; Zhang, T. An Integrative Study of Mycobiome in Different Habitats from a High Arctic Region: Diversity, Distribution, and Functional Role. J. Fungi 2023, 9, 437. https://doi.org/10.3390/jof9040437
Chen X, Yan D, Yu L, Zhang T. An Integrative Study of Mycobiome in Different Habitats from a High Arctic Region: Diversity, Distribution, and Functional Role. Journal of Fungi. 2023; 9(4):437. https://doi.org/10.3390/jof9040437
Chicago/Turabian StyleChen, Xiufei, Dong Yan, Liyan Yu, and Tao Zhang. 2023. "An Integrative Study of Mycobiome in Different Habitats from a High Arctic Region: Diversity, Distribution, and Functional Role" Journal of Fungi 9, no. 4: 437. https://doi.org/10.3390/jof9040437
APA StyleChen, X., Yan, D., Yu, L., & Zhang, T. (2023). An Integrative Study of Mycobiome in Different Habitats from a High Arctic Region: Diversity, Distribution, and Functional Role. Journal of Fungi, 9(4), 437. https://doi.org/10.3390/jof9040437