
Effect of Ectomycorrhizal Fungi on the Drought Resistance of Pinus massoniana Seedlings

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Fungal Preparation and Inoculation
2.3. Experimental Design
2.4. Index Determination
2.4.1. Gas Exchange
2.4.2. Seedling Growth
2.4.3. Colonization, Biomass, Root Vitality, and NSCs Content
2.5. Statistical Analysis
3. Results
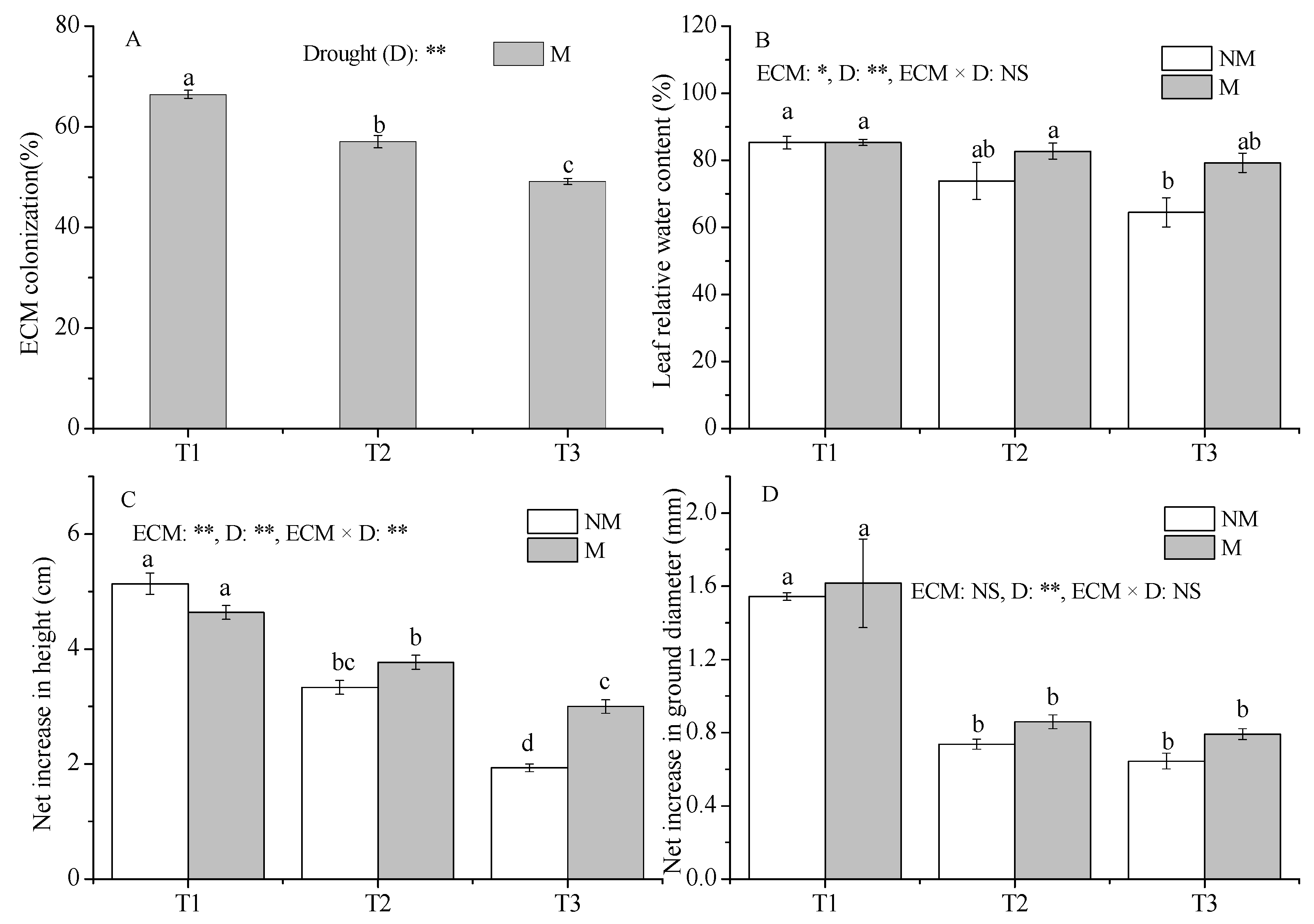
3.1. The Growth of P. massoniana
3.2. Biomass of P. massoniana
3.3. Gas Exchange Parameters of P. massoniana
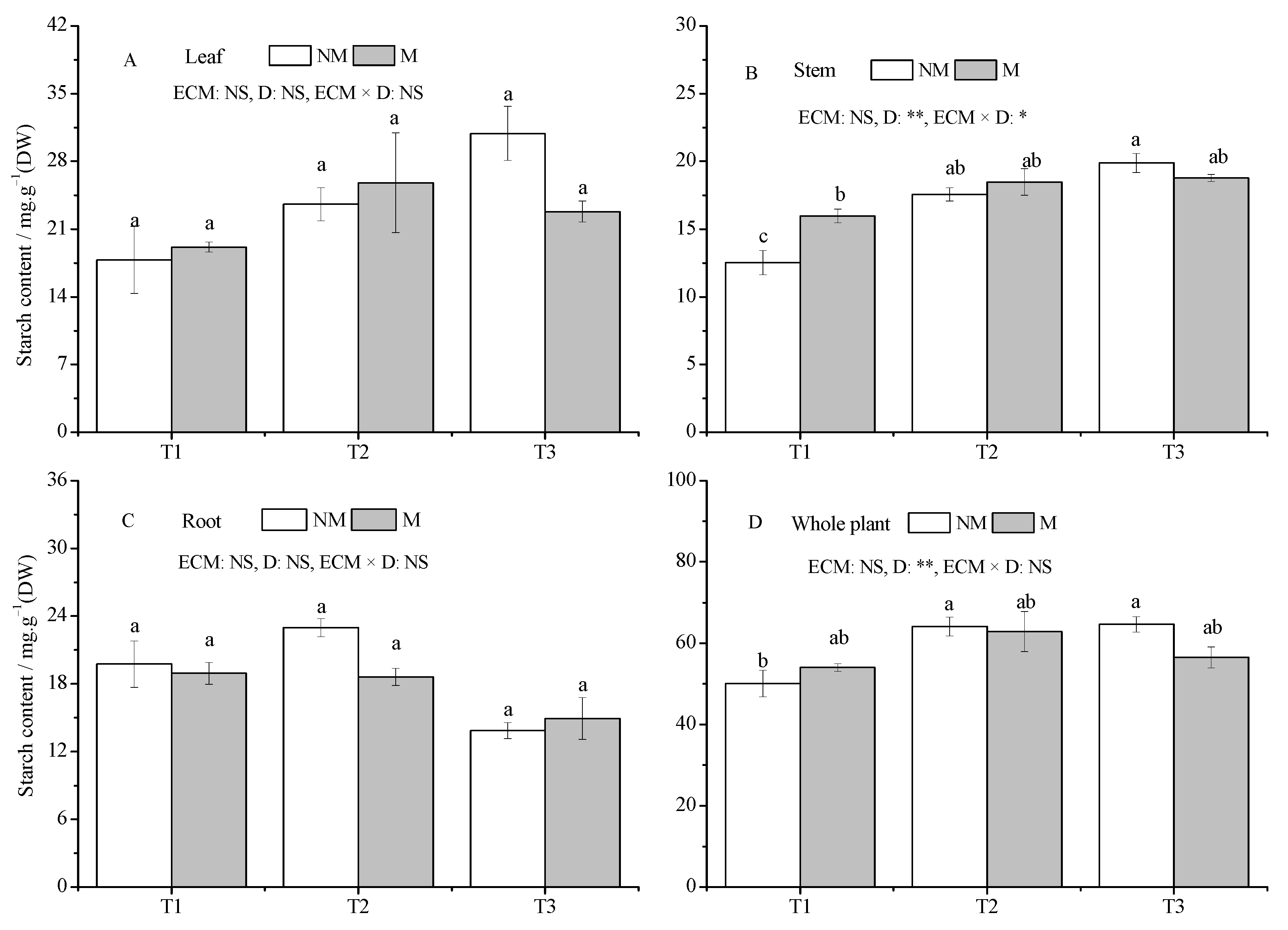
3.4. The Non-Structural Carbohydrates
3.5. Root Vitality
3.6. Correlation Analysis
4. Discussion
4.1. Inoculation Can Improve the Hydraulic Regulation of Seedlings and Maintain More Water
4.2. Inoculation Altered the Content and Distribution of NSCs in Seedlings under Drought Stress
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Choat, B. Predicting thresholds of drought-induced mortality in woody plant species. Tree Physiol. 2013, 33, 669–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plamboeck, A.H.; Dawson, T.E.; Egerton-Warburton, L.M.; North, M.; Bruns, T.D.; Querejeta, J.I. Water transfer via ectomycorrhizal fungal hyphae to conifer seedlings. Mycorrhiza 2007, 17, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, M.; Huygens, D.; Fernandez, C.; Gacitua, Y.; Olivares, E.; Saavedra, I.; Alberdi, M.; Valenzuela, E. Effect of ectomycorrhizal colonization and drought on reactive oxygen species metabolism of Nothofagus dombeyi roots. Tree Physiol. 2009, 29, 1047–1057. [Google Scholar] [CrossRef] [Green Version]
- Sun, P.F.; Shen, Y.F.; Wang, L.J.; Chen, T.; Zhang, M.; Xiao, W.F.; Cheng, R.M. Photosynthetic product allocations of Pinus massoniana seedlings inoculated with ectomycorrhizal fungi along a nitrogen addition gradient. Front. Plant Sci. 2022, 13, 948676. [Google Scholar]
- Vayssières, A.; Pěnčík, A.; Felten, J.; Kohler, A.; Ljung, K.; Martin, F.; Legué, V. Development of the poplar-Laccaria bicolor ectomycorrhiza modifies root auxin metabolism, signaling, and response. Plant Physiol. 2015, 169, 890–902. [Google Scholar] [CrossRef] [Green Version]
- Li, M.T.; Yuan, C.; Zhang, X.H.; Pang, W.B.; Zhang, P.P.; Xie, R.Z.; Lian, C.L.; Zhang, T.X. The Transcriptional Responses of Ectomycorrhizal Fungus, Cenococcum geophilum, to Drought Stress. J. Fungi 2022, 9, 15. [Google Scholar] [CrossRef] [PubMed]
- Colpaert, J.V.; Van Assche, J.A.; Luijtens, K. The growth of the extramatrical mycelium of ectomycorrhizal fungi and the growth response of Pinus sylvestris L. New Phytol. 1992, 120, 127–135. [Google Scholar] [CrossRef]
- Wang, Y.; Ding, G.J. Influence of ectomycorrhiza on nutrient absorption of Pinus massoniana seedlings under water stress. For. Res. Beijing 2013, 26, 227–233. [Google Scholar]
- Wang, Y.; Ding, G. Effects of drought stress on mycorrhizal seedlings growth of Pinus massoniana. J. For. Environ. 2016, 36, 173–179. [Google Scholar]
- Wang, Y.; Zhan, C.; Ding, G.J.; Li, Z.Q. Effect of water stress on physiology and photosynthesis properties of ectomycorrhizal Pinus massoniana seedlings. J. Zhejiang For. Sci. Technol. 2019, 39, 1–8. [Google Scholar]
- Schiestl-Aalto, P.; Ryhti, K.; Mäkelä, A.; Peltoniemi, M.; Bäck, J.; Kulmala, L. Analysis of the NSC storage dynamics in tree organs reveals the allocation to belowground symbionts in the framework of whole tree carbon balance. Front. For. Glob. Chang. 2019, 2, 17. [Google Scholar] [CrossRef] [Green Version]
- Pantin, F.; Fanciullino, A.-L.; Massonnet, C.; Dauzat, M.; Simonneau, T.; Muller, B. Buffering growth variations against water deficits through timely carbon usage. Front. Plant Sci. 2013, 4, 483. [Google Scholar] [CrossRef] [Green Version]
- Adams, H.D.; Germino, M.J.; Breshears, D.D.; Barron-Gafford, G.A.; Guardiola-Claramonte, M.; Zou, C.B.; Huxman, T.E. Nonstructural leaf carbohydrate dynamics of P inus edulis during drought-induced tree mortality reveal role for carbon metabolism in mortality mechanism. New Phytol. 2013, 197, 1142–1151. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Vilalta, J.; Anderegg, W.R.; Sapes, G.; Sala, A. Greater focus on water pools may improve our ability to understand and anticipate drought-induced mortality in plants. New Phytol. 2019, 223, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Sapes, G.; Demaree, P.; Lekberg, Y.; Sala, A. Plant carbohydrate depletion impairs water relations and spreads via ectomycorrhizal networks. New Phytol. 2021, 229, 3172–3183. [Google Scholar] [CrossRef]
- Wiley, E.; Huepenbecker, S.; Casper, B.B.; Helliker, B.R. The effects of defoliation on carbon allocation: Can carbon limitation reduce growth in favour of storage? Tree Physiol. 2013, 33, 1216–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camarero, J.J.; Sangüesa-Barreda, G.; Vergarechea, M. Prior height, growth, and wood anatomy differently predispose to drought-induced dieback in two Mediterranean oak speciesk. Ann. For. Sci. 2016, 73, 341–351. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.Q.; Li, J.; Liu, C.G.; Liu, Y.T.; Zhang, Y.P.; Sha, L.Q.; Wang, Z.; Song, Q.H.; Lin, Y.X.; Zhou, R.W. Carbohydrate dynamics of three dominant species in a Chinese savanna under precipitation exclusion. Tree Physiol. 2018, 38, 1371–1383. [Google Scholar] [CrossRef]
- Liu, H.Y.; Shangguan, H.L.; Zhou, M.; Airebule, P.; Zhao, P.W.; He, W.Q.; Xiang, C.L.; Wu, X.C. Differentiated responses of nonstructural carbohydrate allocation to climatic dryness and drought events in the Inner Asian arid timberline. Agric. For. Meteorol. 2019, 271, 355–361. [Google Scholar] [CrossRef]
- Nehls, U. Mastering ectomycorrhizal symbiosis: The impact of carbohydrates. J. Exp. Bot. 2008, 59, 1097–1108. [Google Scholar] [CrossRef]
- Wang, J.X.; Zhang, H.Q.; Gao, J.; Zhang, Y.; Liu, Y.Q.; Tang, M. Effects of ectomycorrhizal fungi (Suillus variegatus) on the growth, hydraulic function, and non-structural carbohydrates of Pinus tabulaeformis under drought stress. BMC Plant Biol. 2021, 21, 171. [Google Scholar] [CrossRef] [PubMed]
- Selosse, M.A.; Richard, F.; He, X.; Simard, S.W. Mycorrhizal networks: Des liaisons dangereuses? Trends Ecol. Evol. 2006, 21, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Kleczewski, N.M.; Herms, D.A.; Bonello, P. Effects of soil type, fertilization and drought on carbon allocation to root growth and partitioning between secondary metabolism and ectomycorrhizae of Betula papyrifera. Tree Physiol. 2010, 30, 807–817. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Yan, L.J.; Ye, L.; Zhou, J.; Zhang, B.; Peng, W.H.; Zhang, X.P.; Li, X.L. Chinese black truffle (Tuber indicum) alters the ectomycorrhizosphere and endoectomycosphere microbiome and metabolic profiles of the host tree Quercus aliena. Front. Microbiol. 2018, 9, 2202. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Wang, H.Y.; Zhao, X.Z.; Lu, Z.K.; Sun, X.G.; Ding, G.J. Role of Suillus placidus in improving the drought tolerance of Masson Pine (Pinus massoniana Lamb.) seedlings. Forests 2021, 12, 332. [Google Scholar] [CrossRef]
- Hansen, J.; Møller, I. Percolation of starch and soluble carbohydrates from plant tissue for quantitative determination with anthrone. Anal. Biochem. 1975, 68, 87–94. [Google Scholar] [CrossRef]
- Li, H.S.; Sun, Q.; Zhao, S.J. Principles and Techniques of Plant Physiological Biochemical Experiment; Higher Education Press: Beijing, China, 2000; pp. 119–121. [Google Scholar]
- Sebastiana, M.; Duarte, B.; Monteiro, F.; Malhó, R.; Caçador, I.; Matos, A.R. The leaf lipid composition of ectomycorrhizal oak plants shows a drought-tolerance signature. Plant Physiol. Biochem. 2019, 144, 157–165. [Google Scholar] [CrossRef]
- Guehl, J.-M.; Mousain, D.; Falconnet, G.; Gruez, J. Growth, carbon dioxide assimilation capacity and water-use efficiency of Pinus pinea L seedlings inoculated with different ectomycorrhizal fungi. Ann. Des Sci. For. 1990, 47, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Laliberté, E. Below-ground frontiers in trait-based plant ecology. New Phytol. 2017, 213, 1597–1603. [Google Scholar] [CrossRef]
- Lawlor, D.W. Limitation to photosynthesis in water-stressed leaves: Stomata vs. metabolism and the role of ATP. Ann. Bot. 2002, 89, 871–885. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M. Mycorrhizal types differ in ecophysiology and alter plant nutrition and soil processes. Biol. Rev. 2019, 94, 1857–1880. [Google Scholar] [CrossRef]
- Guerin-Laguette, A. Successes and challenges in the sustainable cultivation of edible mycorrhizal fungi–furthering the dream. Mycoscience 2021, 62, 10–28. [Google Scholar] [CrossRef]
- Hartmann, H.; Trumbore, S. Understanding the roles of nonstructural carbohydrates in forest trees–from what we can measure to what we want to know. New Phytol. 2016, 211, 386–403. [Google Scholar] [CrossRef] [Green Version]
- Piper, F.I.; Fajardo, A.; Hoch, G. Single-provenance mature conifers show higher non-structural carbohydrate storage and reduced growth in a drier location. Tree Physiol. 2017, 37, 1001–1010. [Google Scholar] [CrossRef]
- He, W.Q.; Liu, H.Y.; Qi, Y.; Liu, F.; Zhu, X.R. Patterns in nonstructural carbohydrate contents at the tree organ level in response to drought duration. Glob. Chang. Biol. 2020, 26, 3627–3638. [Google Scholar] [CrossRef]
- Körner, C. Carbon limitation in trees. J. Ecol. 2003, 91, 4–17. [Google Scholar] [CrossRef] [Green Version]
- Kozlowski, T. Carbohydrate sources and sinks in woody plants. Bot. Rev. 1992, 58, 107–222. [Google Scholar] [CrossRef]
- Tsuji, C.; Dannoura, M.; Desalme, D.; Angeli, N.; Takanashi, S.; Kominami, Y.; Epron, D. Drought affects the fate of non-structural carbohydrates in hinoki cypress. Tree Physiol. 2022, 42, 784–796. [Google Scholar] [CrossRef]
- Wang, L.; Dai, Y.X.; Guo, J.P.; Gao, R.M.; Wan, X.C. Interaction of hydraulic failure and carbon starvation on Robinia pseudoacacia seedlings during drought. Sci. Silvae Sin. 2016, 52, 1–9. [Google Scholar]
- Landhäusser, S.M.; Lieffers, V.J. Defoliation increases risk of carbon starvation in root systems of mature aspen. Trees 2012, 26, 653–661. [Google Scholar] [CrossRef]
- Sala, A.; Piper, F.; Hoch, G. Physiological mechanisms of drought-induced tree mortality are far from being resolved. New Phytol. 2010, 186, 274–281. [Google Scholar] [CrossRef]
- Trugman, A.; Detto, M.; Bartlett, M.; Medvigy, D.; Anderegg, W.; Schwalm, C.; Schaffer, B.; Pacala, S.W. Tree carbon allocation explains forest drought-kill and recovery patterns. Ecol. Lett. 2018, 21, 1552–1560. [Google Scholar] [CrossRef] [PubMed]
- Sevanto, S. Phloem transport and drought. J. Exp. Bot. 2014, 65, 1751–1759. [Google Scholar] [CrossRef] [Green Version]
- Salmon, Y.; Dietrich, L.; Sevanto, S.; Hölttä, T.; Dannoura, M.; Epron, D. Drought impacts on tree phloem: From cell-level responses to ecological significance. Tree Physiol. 2019, 39, 173–191. [Google Scholar] [CrossRef] [Green Version]
- Silva, E.N.; Ferreira-Silva, S.L.; Viégas, R.A.; Silveira, J.A.G. The role of organic and inorganic solutes in the osmotic adjustment of drought-stressed Jatropha curcas plants. Environ. Exp. Bot. 2010, 69, 279–285. [Google Scholar] [CrossRef]
- Fang, J.; Lutz, J.A.; Shugart, H.H.; Yan, X.D. A physiological model for predicting dynamics of tree stem-wood non-structural carbohydrates. J. Ecol. 2020, 108, 702–718. [Google Scholar] [CrossRef]
- Woodruff, D.R.; Meinzer, F.C.; Marias, D.E.; Sevanto, S.; Jenkins, M.W.; McDowell, N.G. Linking nonstructural carbohydrate dynamics to gas exchange and leaf hydraulic behavior in Pinus edulis and Juniperus monosperma. New Phytol. 2015, 206, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Nehls, U.; Grunze, N.; Willmann, M.; Reich, M.; Küster, H. Sugar for my honey: Carbohydrate partitioning in ectomycorrhizal symbiosis. Phytochemistry 2007, 68, 82–91. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drought Intensity | ECM Treatment | Pn (umol CO2 m−2 s−1) | Ci (umol CO2 mol−1) | Gs (mol H2O m−2 s−1) | Tr (mmol H2O m−2 s−1) | WUE |
|---|---|---|---|---|---|---|
| T1 | NM | 14.09 ± 0.08 a | 290.99 ± 5.24 a | 0.42 ± 0.04 a | 3.93 ± 0.12 a | 36.62 ± 3.13 d |
| M | 14.38 ± 0.34 a | 295.52 ± 2.91 a | 0.44 ± 0.01 a | 4.12 ± 0.04 a | 32.98 ± 1.51 d | |
| T2 | NM | 4.63 ± 0.18 bc | 160.08 ± 3.41 d | 0.04 ± 0.00 b | 0.99 ± 0.05 c | 125.08 ± 2.27 a |
| M | 5.59 ± 0.40 b | 206.31 ± 2.63 b | 0.05 ± 0.00 b | 1.41 ± 0.10 b | 107.69 ± 1.73 b | |
| T3 | NM | 3.79 ± 0.02 d | 152.75 ± 1.19 d | 0.03 ± 0.00 b | 0.94 ± 0.00 c | 129.35 ± 0.77 a |
| M | 4.32 ± 0.08 c | 184.84 ± 2.28 c | 0.05 ± 0.00 b | 1.40 ± 0.01 b | 91.87 ± 2.32 c | |
| ECM | * | ** | NS | ** | ** | |
| D | ** | ** | ** | ** | ** | |
| ECM × D | NS | ** | NS | NS | ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Wang, H.; Zhao, X.; Feng, W.; Ding, G.; Quan, W. Effect of Ectomycorrhizal Fungi on the Drought Resistance of Pinus massoniana Seedlings. J. Fungi 2023, 9, 471. https://doi.org/10.3390/jof9040471
Li M, Wang H, Zhao X, Feng W, Ding G, Quan W. Effect of Ectomycorrhizal Fungi on the Drought Resistance of Pinus massoniana Seedlings. Journal of Fungi. 2023; 9(4):471. https://doi.org/10.3390/jof9040471
Chicago/Turabian StyleLi, Min, Haoyun Wang, Xizhou Zhao, Wanyan Feng, Guijie Ding, and Wenxuan Quan. 2023. "Effect of Ectomycorrhizal Fungi on the Drought Resistance of Pinus massoniana Seedlings" Journal of Fungi 9, no. 4: 471. https://doi.org/10.3390/jof9040471
APA StyleLi, M., Wang, H., Zhao, X., Feng, W., Ding, G., & Quan, W. (2023). Effect of Ectomycorrhizal Fungi on the Drought Resistance of Pinus massoniana Seedlings. Journal of Fungi, 9(4), 471. https://doi.org/10.3390/jof9040471