
Novelties in Macrofungi of the Tropical Montane Cloud Forest in Mexico

 ,
,  ,
,  , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Morphological Studies
2.2. Extraction, Amplification, and Sequencing
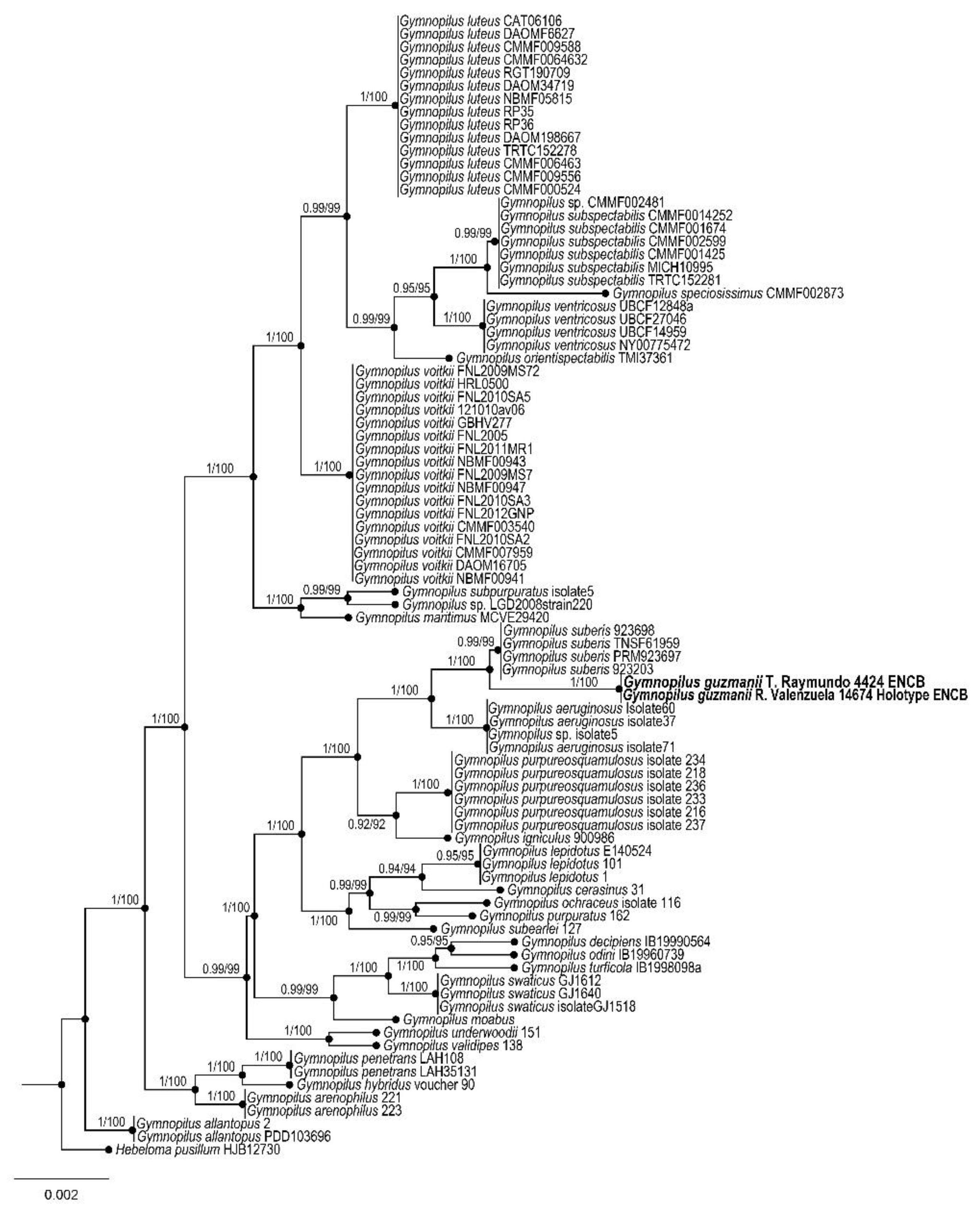
2.3. Phylogenetic Analysis
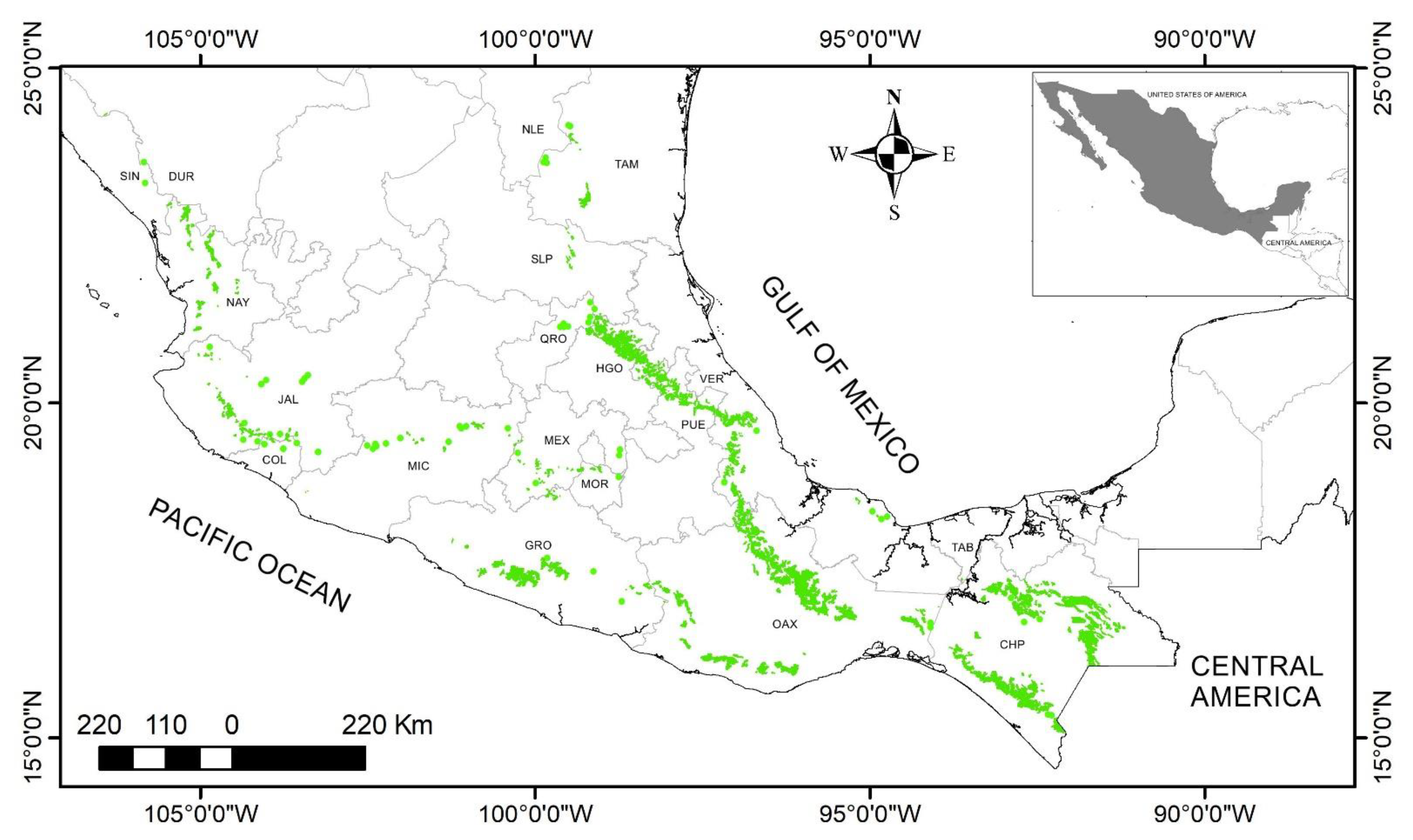
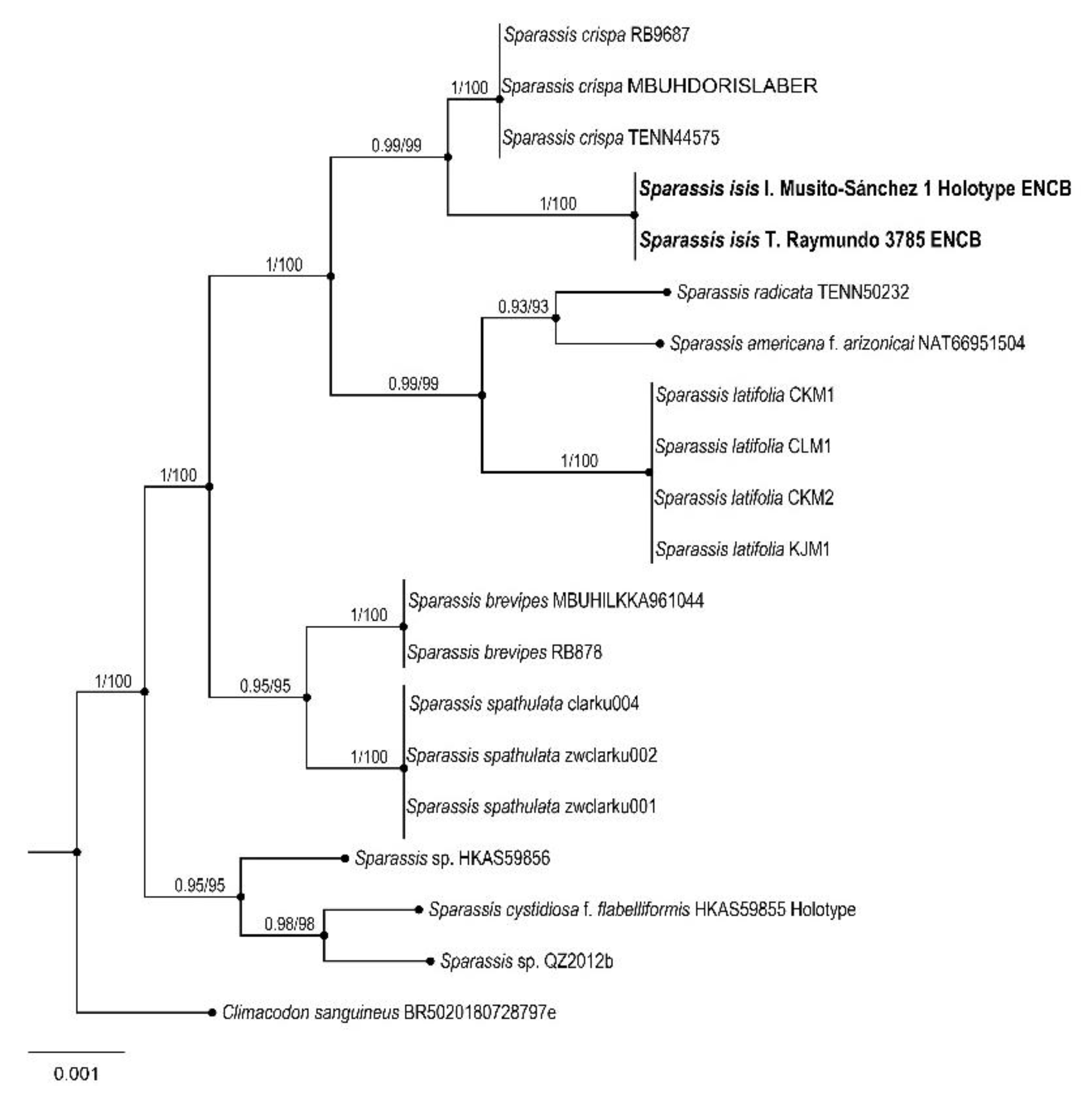
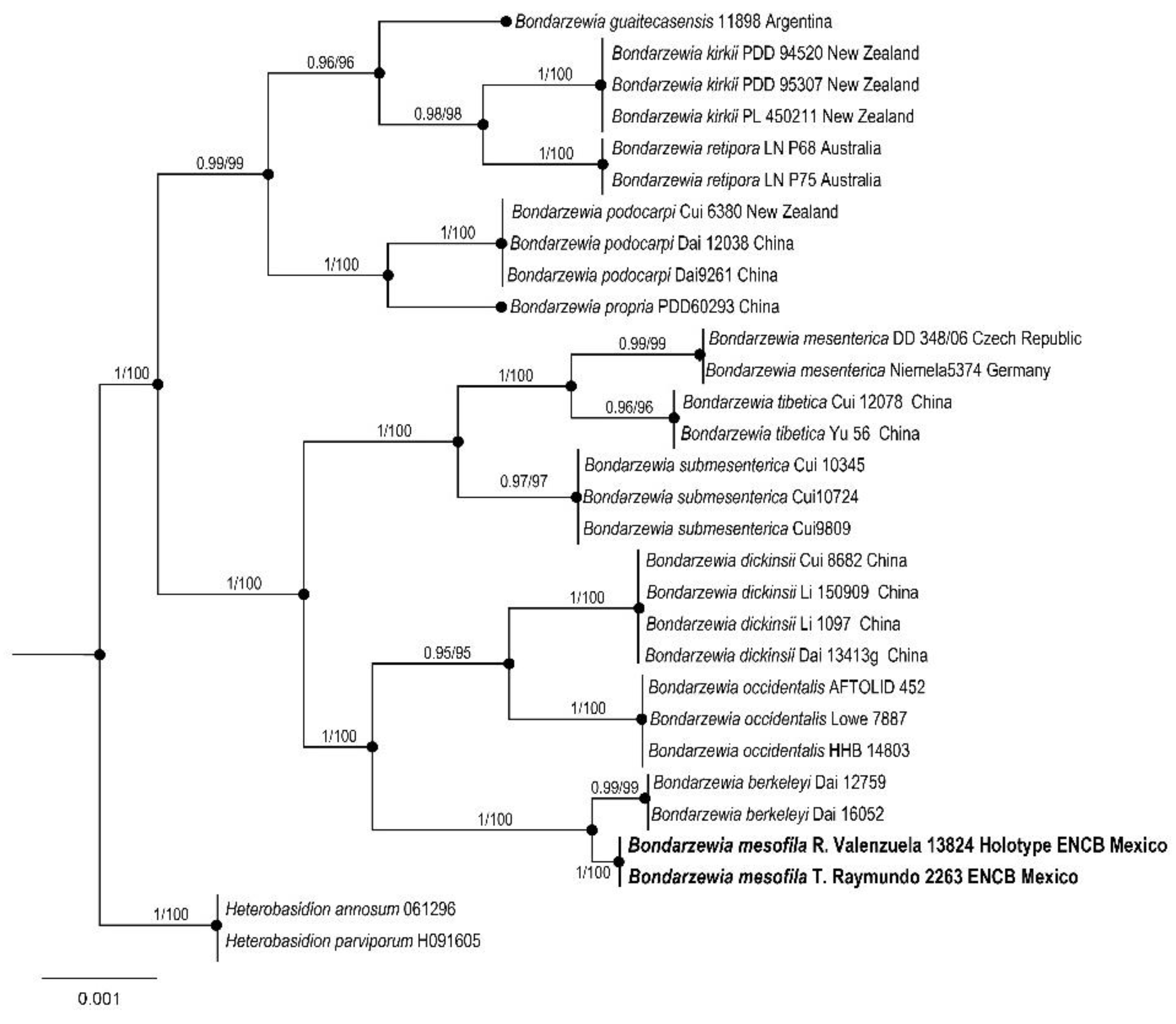
3. Results
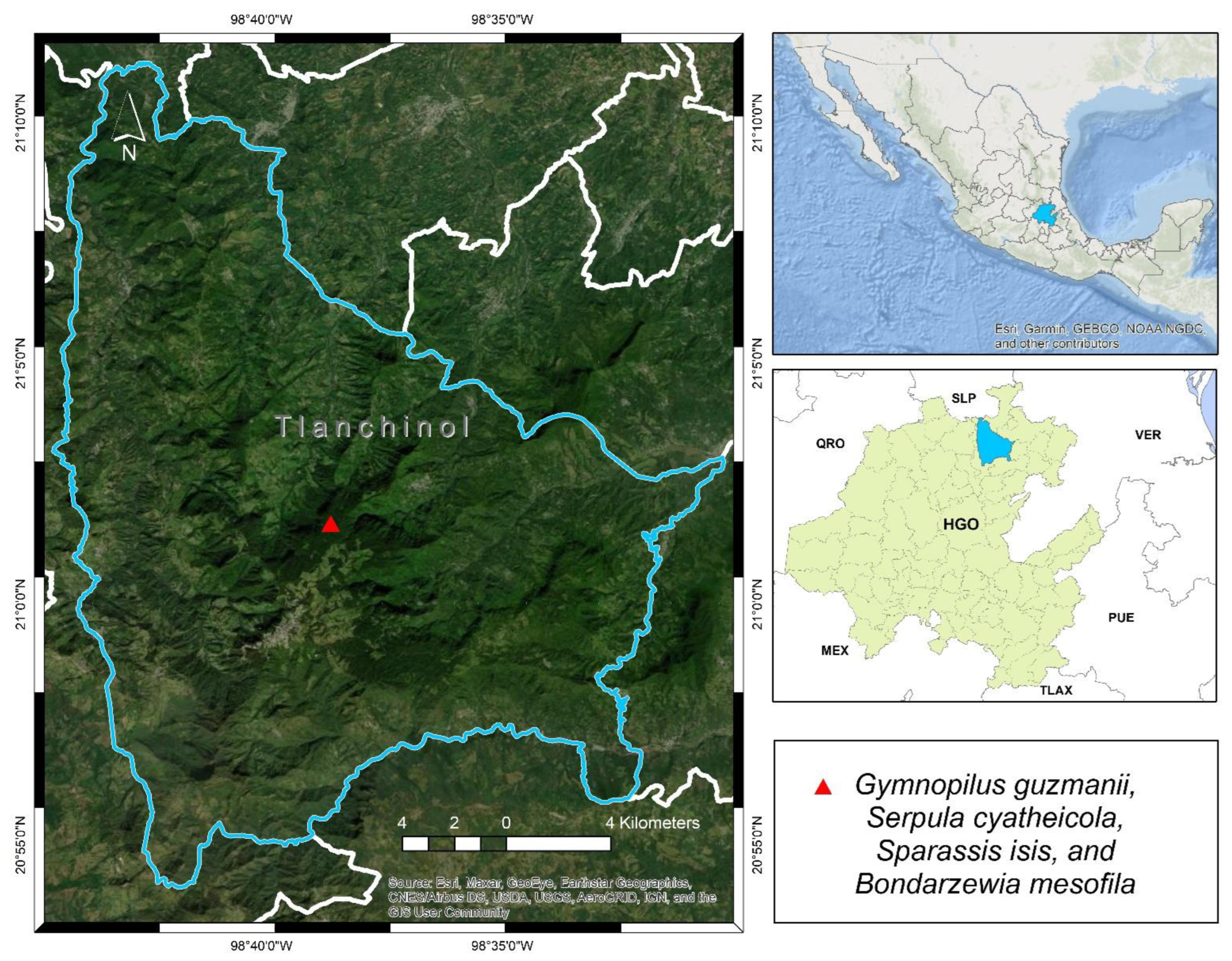
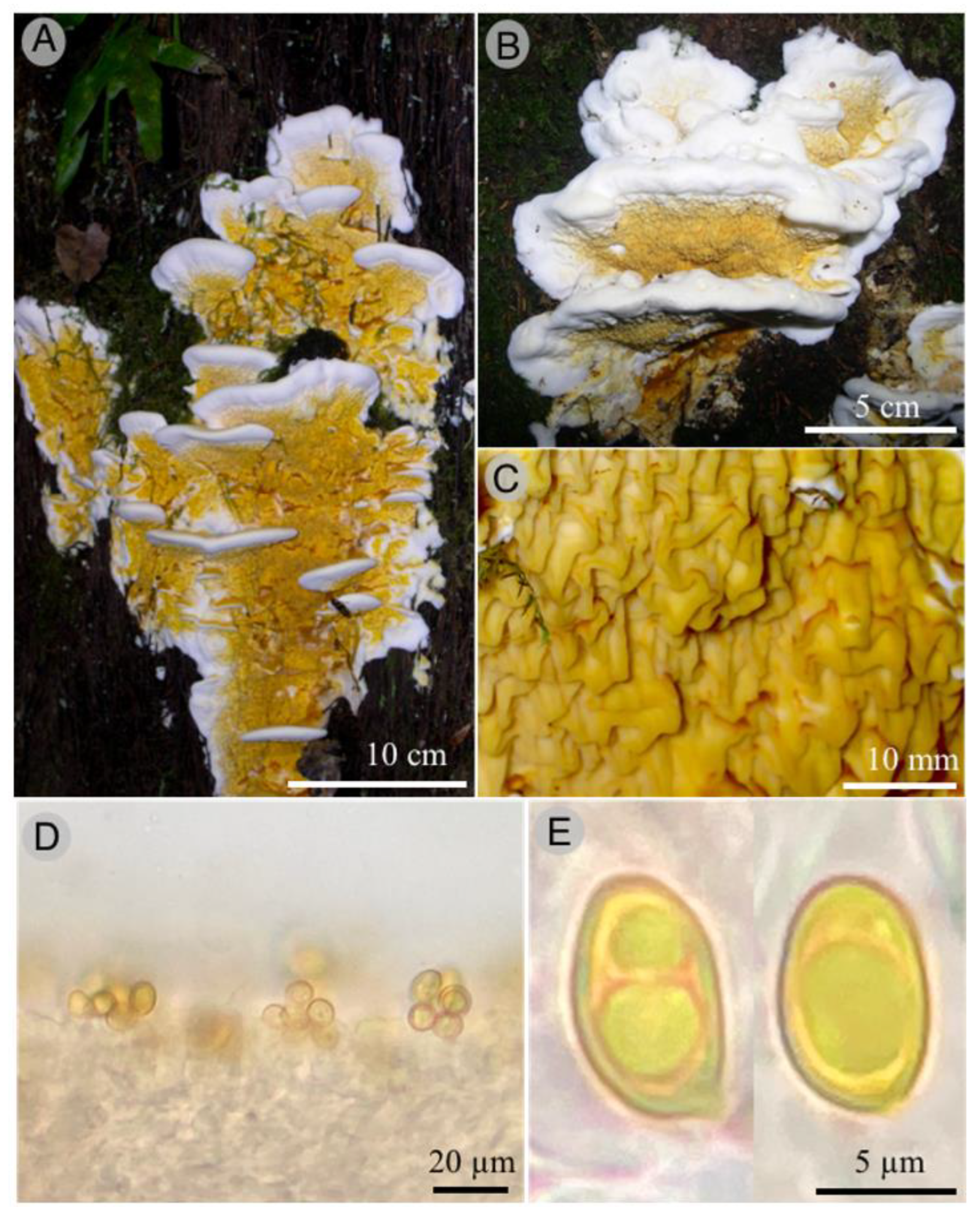
Taxonomy
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rzedowski, J. Análisis preliminar de la flora vascular de los bosques mesófilos de montaña de México. Acta Bot. Mex. 1996, 35, 25–40. [Google Scholar] [CrossRef] [Green Version]
- Blanco-Macías, A. Análisis Sucesional Del Bosque Mesófilo De Montaña En El Rincón, Sierra Norte De Oaxaca. Bachelor Dissertation, Facultad de Estudios Superiores Iztacala, Universidad Nacional Autónoma de México, Los Reyes, Iztacala, Mexico City, Mexico, 2001. [Google Scholar]
- Jardel, E.; Santiago, A.L.; Muñoz, M. El bosque mesófilo de montaña de la Sierra de Manantlán. Tiempos Cienc. 1993, 30, 20–28. [Google Scholar]
- Puig, H. Árboles y Arbustos del Bosque Mesófilo de Montaña de la Reserva: el Cielo de Tamaulipas; Instituto de Ecología & Centre National de la Recherche Scientifique: Xalapa, Mexico, 1993; pp. 1–85. [Google Scholar]
- Rzedowski, J. La Vegetación de México; Limusa: Mexico City, Mexico, 1978; pp. 1–432. [Google Scholar]
- Luna-Vega, I.; Almeida, L.; Villers, L.; Lorenzo, L. Reconocimiento florístico y consideraciones fitogeográficas del bosque mesófilo de montaña de Teocelo, Ver. Bol. Soc. Bot. Mex. 1988, 48, 35–63. [Google Scholar]
- Webster, G.L. The panorama of Neotropical cloud forests. In Biodiversity and Conservation of Neotropical Montane Forests, 2nd ed.; Churchill, S.P., Balslev, H., Forero, E., Luteyn, J.L., Eds.; The New York Botanical Garden: New York, NY, USA, 1995; pp. 53–77. [Google Scholar]
- Bruijnzeel, L.A.; Veneklaas, E.J. Climatic conditions and tropical montane forest productivity: The fog has not lifted yet. Ecology 1998, 79, 3–9. [Google Scholar] [CrossRef]
- Bruijnzeel, L.A.; Proctor, J. Hydrology and biogeochemistry of tropical montane cloud forest: What do we really know? In Tropical Montane Forests, 2nd ed.; Hamilton, L.S., Juvick, J.O., Scatena, F.N., Eds.; Springer: New York, NY, USA, 1995; pp. 38–78. [Google Scholar]
- Boyle, B.L. Changes on Altitudinal and Latitudinal Gradients in Neotropical Montane Forests; Washington University: St. Louis, MO, USA, 1996. [Google Scholar]
- INEGI Carta de uso Actual del Suelo y Vegetación Serie VII; Instituto Nacional de Geografía, Estadística e Informática: Aguascalientes City, Mexico, 2022.
- Williams-Linera, G. Ecología del paisaje y el bosque mesófilo en el centro de Veracruz. Cienc. Y Desarro. 1992, 105, 133–138. [Google Scholar]
- Ortega-Escalona, F.; Castillo-Campos, G. El bosque mesófilo de montaña y su importancia forestal. Ciencias 1996, 43, 32–39. [Google Scholar]
- Challenger, A. Utilización y Conservación de Los Ecosistemas Terrestres de México. Pasado, Presente y Futuro; Conabio-Instituto de Biología, Universidad Nacional Autónoma de México-Agrupación Sierra Madre: Mexico City, Mexico, 1998; pp. 443–518. [Google Scholar]
- Bautista, C.A.; del Castillo, R.F. Soils changes during secondary succession in a tropical montane cloud forest area. Soils Sci. Soc. Am. J. 2005, 69, 906–914. [Google Scholar] [CrossRef]
- Miranda, F. Estudios sobre la vegetación de México, V. Rasgos de la vegetación en la cuenca del Río de las Balsas. Rev. Soc. Mex. Hist. Nat. 1947, 8, 95–114. [Google Scholar]
- Miranda, F.; Sharp, A.J. Characteristics of the vegetation in certain temperate regions of eastern Mexico. Ecology 1950, 31, 313–333. [Google Scholar] [CrossRef]
- Rzedowski, J. Nota sobre el bosque mesófilo de montaña en el Valle de México. An. Inst Biol Ser Bot 1970, 18, 91–106. [Google Scholar]
- Rzedowski, J.; Palacios-Chávez, R. El bosque de Engelhardtia (Oreomunnea) mexicana en la región de la Chinantla (Oaxaca, México) una reliquia del Cenozoico. Bot. Sci. 1977, 36, 93–127. [Google Scholar] [CrossRef] [Green Version]
- Puig, H. La Végétation De La Huasteca (Mexique); Mission Archéologique et Ethnologique Française au Mexique: Mexico City, Mexico, 1976; pp. 1–531. [Google Scholar]
- Puig, H. Análisis fitogeográfico del bosque mesófilo de Gómez Farías. Biotam 1989, 1, 34–53. [Google Scholar]
- Puig, H.; Bracho, R.; Sosa, V.J. El bosque mesófilo de montaña: Composición florística y estructura. In El Bosque Mesófilo de Montaña en Tamaulipas; Puig, H., Bracho, R., Eds.; Instituto de Ecología: Mexico City, Mexico, 1987; pp. 55–79. ISBN 9687213078. [Google Scholar]
- Luna-Vega, I.; Almeida-Leñero, L.; Llorente-Busquets, J. Florística y aspectos fitogeográficos del bosque mesófilo de montaña de las cañadas de Ocuilan, estados de Morelos y México. An. Inst. Biol. Ser. Bot. 1989, 59, 63–87. [Google Scholar]
- Luna-Vega, I.; Ocegueda, S.; Alcántara, O. Florística y notas biogeográficas del bosque mesófilo de montaña del municipio de Tlanchinol, Hidalgo, México. An. Inst. Biol. Ser. Bot. 1994, 65, 31–62. [Google Scholar]
- Meave, J.; Soto, M.A.; Calvo, L.M.; Paz, H.; Valencia, S. Análisis sinecológico del bosque mesófilo de montaña de Omiltemi, Guerrero. Bol. Soc. Bot. Mex. 1992, 52, 31–77. [Google Scholar]
- Acosta, S. Afinidades fitogeográficas del bosque mesófilo de montaña de la zona de Pluma Hidalgo, Oaxaca, México. Polibotánica 1997, 6, 25–39. [Google Scholar]
- Acosta, S. Afinidades de la flora genérica de algunos bosques mesófilos de montaña del nordeste, centro y sur de México: Un enfoque fenético. An. Inst. Biol. Ser. Bot. 2004, 75, 61–72. [Google Scholar]
- Ramírez-Marcial, N. Diversidad florística del bosque mesófilo en el norte de Chiapas y su relación con México y Centroamérica. Bol. Soc. Bot. Mex. 2001, 69, 63–76. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Lücking, R. Fungal diversity revisited: 2.2 to 3.8 million species. Microbiol. Spectr. 2017, 5, FUNK-0052-2016. [Google Scholar] [CrossRef]
- Mueller, G.; Schmit, J.; Leacock, P.; Buyck, B.; Cifuentes, J.; Desjardin, D.; Halling, R.; Hjortstam, K.; Iturriaga, T.; Larsson, K.H.; et al. Global diversity and distribution of macrofungi. Biodivers. Conserv. 2007, 16, 37–48. [Google Scholar] [CrossRef] [Green Version]
- Stadler, M.; Læssøe, T.; Fournier, J.; Decock, C.; Schmieschek, B.; Tichy, H.V.; Peršoh, D. A polyphasic taxonomy of Daldinia (Xylariaceae). Stud. Mycol. 2014, 77, 1–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzmán, G. Diversity and use of traditional Mexican medicinal fungi. A Review. Int. J. Med. Mushrooms 2008, 10, 209–217. [Google Scholar] [CrossRef]
- Aguirre-Acosta, E.; Ulloa, M.; Aguilar, S.; Cifuentes, J.; Valenzuela, R. Biodiversidad de hongos en México. Rev. Mex. Biod. 2014, 85, 76–81. [Google Scholar] [CrossRef] [Green Version]
- Cifuentes, J. Hongos. Catálogo taxonómico de especies de México. In Capital Natural de México, Conocimiento Actual de la Biodiversidad. Vol. 1; CONABIO: Mexico City, Mexico, 2008. [Google Scholar]
- Del Olmo-Ruiz, M.; García-Sandoval, R.; Alcántara-Ayala, O.; Véliz, M.; Luna-Vega, I. Current knowledge of fungi from Neotropical montane cloud forests: Distributional patterns and composition. Biodivers. Conserv. 2017, 26, 1919–1942. [Google Scholar] [CrossRef]
- Miller, S.L.; Larsson, E.; Larsson, K.H.; Verbeken, A.; Nuytinck, J. Perspectives in the new Russulales. Mycologia 2006, 98, 960–970. [Google Scholar] [CrossRef]
- He, M.-Q.; Zhao, R.-L.; Hyde, K.D.; Begerow, D.; Kemler, M.; Yurkov, A.; McKenzie, E.H.C.; Raspé, O.; Kakishima, M.; Sánchez-Ramírez, S.; et al. Notes, outline and divergence times of Basidiomycota. Fungal Divers. 2019, 99, 105–367. [Google Scholar] [CrossRef] [Green Version]
- Matheny, P.B.; Curtis, J.M.; Hofstetter, V.M.; Catherine Aime, M.C.; Moncalvo, J.M.; Ge, Z.W.; Yang, Z.L.; Slot, J.C.; Ammirati, J.F.; Baroni, T.B.; et al. Major clades of Agaricales: A multilocus phylogenetic overview. Mycologia 2006, 98, 982–995. [Google Scholar] [CrossRef]
- Binder, M.; Justo, A.; Riley, R.; Salamov, A.; Lopez-Giraldez, F.; Sj€okvist, E.; Copeland, A.; Foster, B.; Sun, H.; Larsson, E.; et al. Phylogenetic and phylogenomic overview of the Polyporales. Mycologia 2013, 105, 1350–1373. [Google Scholar] [CrossRef] [Green Version]
- Binder, M.; Hibbett, D.S. Molecular systematics and biological diversification of Boletales. Mycologia 2006, 98, 971–981. [Google Scholar] [CrossRef]
- Justo, A.; Miettinen, O.; Floudas, D.; Ortiz-Santana, B.; Sjökvist, E.; Lindner, D.; Nakasone, K.; Niemelä, T.; Larsson, K.H.; Ryvarden, L.; et al. A revised family-level classification of the Polyporales (Basidiomycota). Fungal Biol. 2017, 121, 798–824. [Google Scholar] [CrossRef]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Eyre: London, UK, 1978; pp. 1–252. [Google Scholar]
- Bessette, A.E.; Roody, W.C.; Bessette, A.R. North American Boletes: A Color Guide to the Fleshy Pored Mushrooms, 1st ed.; Syracuse University Press: Syracuse, NY, USA, 2000; pp. 1–396. [Google Scholar]
- Ulloa, M.; Hanlin, R.T. Illustrated Dictionary of Mycology, 2nd ed.; APS Press: St. Paul, MN, USA, 2012; pp. 1–762. [Google Scholar]
- Martínez-González, C.R.; Ramírez-Mendoza, R.; Jiménez-Ramírez, J.; Gallegos-Vázquez, C.; Luna-Vega, I. Improved method for genomic DNA extraction for Opuntia Mill. (Cactaceae). Plant Methods 2017, 13, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 135–322. [Google Scholar] [CrossRef]
- Matheny, P.B.; Wang, Z.; Binder, M. Contributions of rpb2 and tef1 to the phylogeny of mushrooms and allies (Basidiomycota, Fungi). Mol. Phylogenetics Evol. 2007, 43, 430–451. [Google Scholar] [CrossRef]
- Wu, G.; Feng, B.; Xu, J.; Zhu, X.T.; Li, Y.C.; Zeng, N.K.; Hosen, M.I.; Yang, Z.L. Molecular phylogenetic analyses redefine seven major clades and reveal 25 new generic lineages in the fungal family Boletaceae. Fungal Divers. 2014, 69, 93–115. [Google Scholar] [CrossRef]
- Giachini, A.J.; Hosaka, K.; Nouhra, E.; Spatafora, J.; Trappe, J.M. Phylogenetic relationships of the Gomphales based on nuc-25S-rDNA, mit-12S-rDNA, and mit-atp6-DNA combined sequences. Fungal Biol. 2010, 114, 224–234. [Google Scholar] [CrossRef]
- Khan, J.; Kiran, M.; Jabeen, S.; Sher, H.; Khalid, A.N. Gymnopilus penetrans and G. swaticus sp. nov. (Agaricomycota: Hymenogastraceae); a new record and a new species from northwest Pakistan. Phytotaxa 2017, 312, 60–70. [Google Scholar] [CrossRef] [Green Version]
- Acharya, K.; Paloi, S.; Dutta, A.K.; Sikder, R.; Saha, T. Gymnopilus purpureosquamulosus Høil. (Agaricales, Basidiomycota): A new distributional record from India. Check List 2017, 13, 2064. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.I.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignments based on fast Fourier transform. Nucleic. Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2017, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Müller, K. SeqState. Appl. Bioinforma. 2005, 4, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.H.; Das, K.; Bera, I.; Chen, Y.H.; Bhatt, R.P.; Ghosh, A.; Hembrom, M.E.; Hofstetter, V.; Parihar, A.; Vizzini, A.; et al. Fungal Biodiversity Profiles 81-90. Cryptogam. Mycol. 2019, 40, 57–95. [Google Scholar] [CrossRef]
- Hughes, K.H.; Reboredo-Segovia, A.; Petersen, R.H. Transatlantic disjunction in fleshy fungi. I. The Sparassis crispa complex. Mycol. Prog. 2014, 13, 407–427. [Google Scholar] [CrossRef]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis, Version 3.70. 2021. Available online: http://mesquiteproject.org (accessed on 19 October 2022).
- Song, J.; Chen, J.-J.; Wang, M.; Chen, Y.-Y.; Cui, B.-K. Phylogeny and biogeography of the remarkable genus Bondarzewia (Basidiomycota, Russulales). Sci. Rep. 2016, 6, 34568. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frandsen, P.B.; Calcott, B.; Mayer, C.; Lanfear, R. Automatic selection of partitioning schemes for phylogenetic analyses using iterative k-means clustering of site rates. BMC Evol. Biol. 2015, 15, 13. [Google Scholar] [CrossRef] [Green Version]
- Lanfear, R.; Calcott, B.; Kainer, D.; Mayer, C.; Stamatakis, A. Selecting optimal partitioning schemes for phylogenomic datasets. BMC Evol. Biol. 2014, 14, 82. [Google Scholar] [CrossRef] [Green Version]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. Partition Finder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Suchard, M.A.; Xie, D.; Drummond, A.J. Tracer v1.6, 2014. Available online: http://beast.bio.ed.ac.uk/Tracer(accessed on 31 July 2022).
- Thorn, R.G.; Malloch, D.W.; Saar, I.; Lamoureux, Y.; Nagasawa, E.; Redhead, S.A.; Margaritescu, S.; Moncalvo, J.-M. New species in the Gymnopilus junonius group (Basidiomycota: Agaricales). Botany 2020, 98, 293–315. [Google Scholar] [CrossRef]
- Singer, R. The Agaricales in Modern Taxonomy, 4th ed.; Koeltz Scientific Books: Koenigstein, Germany, 1986; pp. 1–981. [Google Scholar]
- Binder, M.; Larsson, K.H.; Matheny, P.B.; Hibbett, D.S. Amylocorticiales ord. nov. and Jaapiales ord. nov.: Early diverging clades of Agaricomycetidae dominated by corticioid forms. Mycologia 2010, 102, 865–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsson, K.H. Re-thinking the classification of corticioid fungi. Mycol. Res. 2007, 111, 1040–1063. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.L.; Li, G.J.; Sánchez-Ramírez, S.; Stata, M.; Yang, Z.L.; Wu, G.; Dai, Y.C.; He, S.H.; Cui, B.K.; Zhou, J.L.; et al. A six-gene phylogenetic overview of Basidiomycota and allied phyla with estimated divergence times of higher taxa and a phyloproteomics perspective. Fungal Divers. 2017, 84, 43–74. [Google Scholar] [CrossRef]
- Coker, B. The genera Serpula and Meruliporia. Mycologia 1957, 49, 197–225. [Google Scholar] [CrossRef]
- Skrede, I.; Engh, I.B.; Binder, M.; Carlsen, T.; Kauserud, H.; Bendiksby, M. Evolutionary history of Serpulaceae (Basidiomycota): Molecular phylogeny, historical biogeography and evidence for a single transition of nutritional mode. J. Evol. Biol. 2011, 11, 230. [Google Scholar] [CrossRef] [Green Version]
- Carslen, T.; Engh, I.B.; Decock, C.; Rajchenberg, M.; Kauserud, H. Multiple cryptic species with divergent substrate affinities in the Serpula himantioides species complex. Fungal Biol. 2010, 115, 54–61. [Google Scholar] [CrossRef]
- Kauserud, H.; Svegården, I.B.; Sætre, G.P.; Knudsen, H.; Stensrud, Ø.; Schmidt, S.; Doi, S.; Sugiyama, T.; Högberg, N. Asian origin and rapid global spread of the destructive dry rot fungus Serpula lacrymans. Mol. Ecol. 2007, 16, 3350–3360. [Google Scholar] [CrossRef]
- Tapia, F.; Chacón, S.; Pérez-Gorjón, S. Hongos corticioides (Agaricomycetes) citados de México. Acta Bot. Mex. 2016, 114, 15–50. [Google Scholar] [CrossRef] [Green Version]
- Ginns, J.H. The genus Merulius IV. Species proposed by Berkeley, by Berkeley and Curtis, and by Berkeley and Broome. Mycologia 1971, 63, 219–236. [Google Scholar] [CrossRef]
- Desjardin, D.E.; Wang, Z.; Binder, M.; Hibbett, D.S. Sparassis cystidiosa sp. nov. from Thailand is described using morphological and molecular data. Mycologia 2004, 96, 1010–1014. [Google Scholar] [CrossRef]
- Blanco-Dios, J.B.; Wang, Z.; Binder, M.; Hibbett, D. A new Sparassis species from Spain described using morphological and molecular data. Mycol. Res. 2006, 110, 1227–1231. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Feng, B.; Yang, Z.L.; Dai, Y.C.; Wang, Z.; Tolgor, B. New species and distinctive geographical divergences of the genus Sparassis (Basidiomycota): Evidence from morphological and molecular data. Mycol. Prog. 2013, 12, 445–454. [Google Scholar] [CrossRef]
- Chen, J.J.; Cui, B.K.; He, S.H.; Cooper, J.A.; Barrett, M.D.; Chen, J.L.; Song, J.; Dai, J.C. Molecular phylogeny and global diversity of remarkable genus Bondarzewia (Basidiomycota, Russulales). Mycologia 2016, 108, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Das, K.; Parihar, A.; Hembrom, M.E. A new species of Bondarzewia from India. Turk. J. Bot. 2015, 39, 128–133. [Google Scholar] [CrossRef]
- Luna-Vega, I.; Velázquez, A.; Velázquez, E. México. In Bosques Nublados del Neotrópico; Kappelle, M., Brown, A.D., Eds.; IMBio: Santo Domingo de Heredia, Costa Rica, 2001; pp. 183–229. [Google Scholar]
- Sánchez-González, L.A.; Morrone, J.J.; Navarro-Sigüenza, A.G. Distributional patterns of the Neotropical humid montane forest avifaunas. Biol. J. Linn. Soc. 2008, 94, 175–194. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.C.; Yang, Z.L.; Cui, B.K.; Wu, G.; Yuan, H.S.; Zhou, L.W.; He, S.H.; Ge, Z.W.; Wu, F.; Wei, Y.L.; et al. Diversity and systematics of the important macrofungi in Chinese forests. Mycosystema 2021, 40, 770–805. [Google Scholar] [CrossRef]
- Wang, Y.R.; Dai, Y.C.; Liu, H.G.; Vlasák, J.; Buchanan, P.; Yuan, Y.; Wu, Y.D. A new contribution to Megasporoporia sensu lato: Six new species and three new combinations. Front. Microbiol. 2022, 13, 1046777. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- García-Guzmán, O.M.; Garibay-Orijel, R.; Hernández, E.; Arellano-Torres, E.; Oyama, K. Word-wide meta-analysis of Quercus forests ectomycorrhizal fungal diversity reveals southwestern Mexico as a hotspot. Mycorrhiza 2017, 27, 811–822. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Loci/Segment | Primer | Sequence 5′-3′ | T (°C) | Reference |
|---|---|---|---|---|
| ITS | ITS5 | GGAAGTAAAAGTCGTAACAAGG | 58 | [48] |
| ITS4 | TCCTCCGCTTATTGATATGC | 58 | [48] | |
| nLSU | LROR | ACCCGCTGAACTTAAGC | 48 | [48] |
| LR3 | GGTCCGTGTTTCAAGAC | 48 | [48] | |
| rpb2 | bRPB2-6R2 | GARTGYCCDGGDCAYTTYGG | 52 | [49] |
| bRPB2-7R2 | CCNGCDATNTCRTTRTCCATRTA | 52 | [49] | |
| tef1 | EF1-B-F1 | ATYGCTTTAGAAAGTTYMTTTGC | 53 | [50] |
| EF1-B-R | GGDATRAARWAWGARAARAARTG | 53 | [50] | |
| atp6 | atp6-5 | ATYGCTTTAGAAAGTTYMTTTGC | 56 | [51] |
| atp6-6 | GGDATRAARWAWGARAARAARTG | 55 | [51] | |
| SSU | MS1 | CAGCAGTCAAGAATATTAGTCAATG | 63 | [48] |
| MS2 | GCGGATTATCGAATTAAATAAC | 53 | [48] |
| Species Name | Isolate/Voucher/Strain | GenBank Accessions |
|---|---|---|
| ITS | ||
| Gymnopilus aeruginosus (Peck) Singer | Isolate 60 | AY280975 |
| Isolate 71 | AT254102 | |
| Isolate 37 | AY280974 | |
| Gymnopilus cerasinus (Sacc.) Guzm.-Dáv., G.M. Muell., J. Cifuentes, A.N. Mill., and Santerre | 31 | GH412552 |
| Gymnopilus igniculus Deneyer, P.-A. Moreau and Wuilb. | 900986 | HY142500 |
| Gymnopilus guzmanii | T. Raymundo 4424 | OQ749940 |
| R. Valenzuela 14674 | OQ749941 | |
| Gymnopilus lepidotus Hesler | E140524 | YU415875 |
| 101 | UI475222 | |
| 1 | HJ410256 | |
| Gymnopilus luteus (Peck.) Hesler | CAT06106 | OP985111 |
| DAOMF6627 | OM672062 | |
| CMMF009588 | MN477932 | |
| CMMF0064632 | MN477919 | |
| RGT190709 | MN477917 | |
| DAOM34719 | MN477293 | |
| NBMF05815 | MN453485 | |
| RP35 | MN453484 | |
| RP36 | MN453483 | |
| DAOOM198668 | MN718842 | |
| TRTC152278 | MN206894 | |
| CMMF006463 | MN206893 | |
| CMMF009556 | MN206892 | |
| CMMF000524 | MN206891 | |
| Gymnopilus maritimus Contu, Guzm.-Dáv., A. Ortega, and Vizzini | MCVE29420 | JK210152 |
| Gymnopilus ochraceus Høil. | Isolate 116 | HJ125488 |
| Gymnopilus orientispectabilis Nagas, Malloch, and Thorn | TM137361 | YU125412 |
| Gymnopilus purpuratus (Cooke and Massee) Singer | 162 | HJ140258 |
| Gymnopilus purpureosquamulosus Høil. | Isolate 234 | HJ521410 |
| Isolate 218 | GH150236 | |
| Isolate 236 | GF852014 | |
| Isolate 233 | YG521400 | |
| Isolate 216 | GF251478 | |
| Isolate 237 | GB512014 | |
| Gymnopilus speciosissimus Y. Lamoureux, Malloch, and Thorn | CMMF002873 | TG512152 |
| Gymnopilus suberis (Maire) Singer | 923698 | GF250124 |
| TNSF61959 | FG214585 | |
| PRM923697 | FF510535 | |
| 9232063 | FV475210 | |
| Gymnopilus subpurpuratus Guzm.-Dáv. and Guzmán | Isolate 5 | FG152024 |
| Gymnopilus subspectabilis Hesler | CMMF001425 | HB415204 |
| CMMF001674 | BH125478 | |
| CMMF002599 | GH521402 | |
| CMMF001425 | HG251200 | |
| MICH10995 | BG520526 | |
| TRTC152281 | JH250145 | |
| Gymnopilus ventricosus (Earle) Hesler | UBCF12848a | GH250210 |
| UBCF27046 | TY820152 | |
| UBCF14959 | HG521520 | |
| NY00775472 | GB520145 | |
| Gymnopilus voitkii Malloch and Thorn | FNL200MS72 | HJ452145 |
| HRL0500 | HN452104 | |
| FNL2010SA5 | UY541203 | |
| 121010av06 | HN204789 | |
| GBHV277 | YH520384 | |
| FNL2005 | TG570134 | |
| FNL2011MR1 | NH521452 | |
| NBMF00943 | HJ251024 | |
| FNL2009MS7 | BN250145 | |
| NBMF00947 | GF204752 | |
| FNL2010SA3 | HJ520214 | |
| FNL2012GNP | FV520145 | |
| CMMF003540 | GH102547 | |
| FNL2010SA2 | TY520147 | |
| CMMF007959 | VF520147 | |
| DAOM16705 | VB254054 | |
| NBMF00941 | VT012585 |
| Species Name | Isolate/Voucher/Strain | GenBank Accessions |
|---|---|---|
| ITS | ||
| Serpula cyatheicola | T. Raymundo 3784 ENCB | OQ749942 |
| T. Raymundo 2891 ENCB | OQ749943 | |
| Serpula dendrocalami C.L. Zhao | 3113 | MK863408 |
| 3136 | MK863407 | |
| 3111 | MK863406 | |
| 3203 | MK863405 | |
| 3318 | MK863404 | |
| 3321 | MK863403 | |
| Serpula himantioides (Fr.) P. Karst. | SH164 | LC710851 |
| SH17 | LC710850 | |
| SH129 | LC710849 | |
| SH130 | LC710848 | |
| SH18 | LC710847 | |
| SH141 | LC710846 | |
| RLG 383 | LC710845 | |
| SH99 | LC710844 | |
| SH20 | LC710843 | |
| SH127 | LC710842 | |
| SH133 | LC710841 | |
| SH115 | LC637898 | |
| SH26 | LC637897 | |
| SH168 | LC637896 | |
| SH169 | LC637895 | |
| SH113 | LC637894 | |
| Serpula incrassata (Berk. and M.A. Curtis) Donk | SINC | HM135648 |
| Si14 | HM135647 | |
| DAOM 170590 | GU187541 | |
| Serpula lacrymans (Wulfen) J. Schröt. | CZ1 | GU066830 |
| CZ2 | MW491273 | |
| SL199 | HB158420 | |
| SL1 | BV120158 | |
| SL5 | HV157863 | |
| REG 383 | HG547896 | |
| SL 198 | FG541256 | |
| Serpula lacrymans var. shastensis (Harmsen) Ginns and M.N.L. Lefebvre | SHA30 | HD587452 |
| SHA8 | TG558412 | |
| Serpula similis (Berk. and Broome) Ginns | ZD 16061808 | MN523308 |
| ZD160827033 | MN523307 | |
| Serpula tignicola (Harmsen) M.P. Christ. | CBS 31154 | GU187543 |
| CBS 311542 | HJ452102 | |
| Serpula sp. | 1524 | HN458520 |
| Species Name | Isolate/Voucher/Strain | GenBank Accessions | |||
|---|---|---|---|---|---|
| ITS | nLSU | rpb2 | atp6 | ||
| Sparassis americana f. arizonica R.H. Petersen | NAT66951504 | MW403771 | MF693910 | ----- | ----- |
| Sparassis brevipes Krombh. | RB878 | MH861680 | MH873392 | AG452102 | ----- |
| MBUHILKKA961044 | KP100510 | KP100511 | AT412035 | ----- | |
| Sparassis crispa (Wulfen) Fr. | RB9687 | KJ754521 | KJ850214 | YD452110 | ----- |
| MBUHDORISLABER | HJ541225 | KL541252 | ----- | ----- | |
| TENN44575 | JK254102 | KL458520 | ----- | AY452102 | |
| Sparassis cystidiosa f. flabelliformis Q. Zhao, Zhu L. Yang, and Y.C. Dai | HKAS59855 | KY428924 | ----- | ----- | ----- |
| Sparassis isis | I Musito-Sánchez 1 | OQ749944 | OQ747755 | OX5210I7 | OX52102I |
| T. Raymundo 3785 | OQ749945 | OQ747756 | OX5210I8 | OX52I044 | |
| Sparassis latifolia Y.C. Dai and Zheng Wang | CKM1 | JK521452 | HJ874520 | AD478520 | ----- |
| CLM1 | LK521477 | KJ514268 | HG210548 | ----- | |
| CKM2 | JI525455 | IK870256 | ----- | JN452012 | |
| KJM1 | KL478601 | KL472500 | ----- | HF420156 | |
| Sparassis radicata Weir | TENN50232 | LK452102 | KI210045 | ----- | JH021585 |
| Sparassis spathulata (Schwein.) Fr. | Clarku004 | LO478521 | IO541256 | HF478520 | NB023575 |
| Zwclarku002 | KJ457424 | PO582114 | GH021452 | JV521452 | |
| Zwclarku001 | IK547888 | OL452101 | ----- | ----- | |
| Sparassis sp. | QZ2012B | JI471250 | OP852014 | ----- | ----- |
| Sparassis sp. | HKAS59856 | OK541200 | KL457520 | ----- | ----- |
| Species Name | Isolate/Voucher/Strain | GenBank Accessions | |||
|---|---|---|---|---|---|
| ITS | nLSU | tef1 | SSU | ||
| Bondarzewia berkeleyi (Fr.) Bondartsev and Singer | Dai 12759 | KJ583202 | KJ583216 | KX066138 | KX066169 |
| Dai 16052 | KX263720 | KX263 | ----- | ----- | |
| Bondarzewia dickinsii (Berk.) Jia J. Chen, B.K. Cui, and Y.C. Dai | Cui 8682 | KJ583209 | KJ583223 | KX066136 | KX066167 |
| Dai 13413 | KJ583210 | KJ583224 | KX066137 | KX066168 | |
| Li 150909/19 | KX263721 | KX263723 | ----- | ----- | |
| Li 1097 | FJ644288 | ----- | ----- | ----- | |
| Bondarzewia guaitecasensis (Henn.) J.E. Wright | Rajchenberg 11898 | FJ644287 | ----- | ----- | KX066175 |
| Bondarzewia kirkii J.A. Cooper, Jia J. Chen, and B.K. Cui | PDD 94520 | KJ583215 | KJ583229 | KX252748 | KX066180 |
| JAC 10839 | KJ734674 | KM067469 | KX252747 | KX066179 | |
| PL 450211 | KJ583214 | KJ583228 | KX252746 | KX066178 | |
| Bondarzewia mesenterica (Schaeff.) Kreisel | DD 348/06 | KM243328 | KM243331 | KX066147 | KX066182 |
| Niemelä | KM067468 | KM067470 | KX066146 | KX066181 | |
| Bondarzewia mesofila | R. Valenzuela 13824 | OQ749938 | OQ747753 | OQ5210I4 | OQ749946 |
| T. Raymundo 2263 | OQ749939 | OQ747754 | OQ52I027 | OQ749947 | |
| Bondarzewia occidentalis Jia J. Chen, B.K. Cui, and Y.C. Dai | Lowe 7887 | KM243330 | KM243333 | KX066143 | KX066177 |
| HHB 14803 | KM243329 | KM243332 | KX066142 | KX066176 | |
| AFTOL-ID 452 | DQ200923 | DQ234539 | DQ059044 | ----- | |
| Bondarzewia podocarpi Y.C. Dai and B.K. Cui | Cui 6380 | KJ583206 | KJ583220 | KX252745 | KX066174 |
| Dai 9261 | KJ583207 | KJ583221 | KX252743 | KX066172 | |
| Dai 12038 | KJ583208 | KJ583222 | KX252744 | KX066173 | |
| Bondarzewia propria (Lloyd) J.A. Cooper | PDD 60293 | KJ583213 | KJ583227 | ----- | ----- |
| Bondarzewia retipora (Cooke) M.D. Barrett | LNP 68 | KJ747633 | KJ747630 | ----- | KX066144 |
| LNP 75 | KJ747632 | KJ747629 | ----- | KX066145 | |
| Bondarzewia submesenterica Jia J. Chen, B.K. Cui, and Y.C. Dai | Cui 9809 | KJ583203 | KJ583217 | KX066141 | KX066171 |
| Cui 10345 | KJ583204 | KJ583218 | KX066140 | KX066170 | |
| Cui 10724 | KJ583205 | KJ583219 | KX066139 | ----- | |
| Bondarzewia tibetica B.K. Cui, J. Song, and Jia J. Chen | Cui 12078 | KT693020 | KT693204 | KX066149 | KX066184 |
| Yu 56 | KT693203 | KT693205 | KX066148 | KX066183 | |
| Heterobasidion annosum (Fr.) Bref. | 06129/6 | KJ583211 | KJ583225 | KX252741 | KJ651577 |
| Heterobasidion parviporum Niemelä and Korhonen | H 091605 | KJ651503 | KJ651561 | KX252742 | KJ651622 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valenzuela, R.; Luna-Vega, I.; Martínez-Pineda, M.; Martínez-González, C.R.; García-Jiménez, J.; de la Fuente, J.; Bautista-Hernández, S.; Acosta-Castellanos, S.; Raymundo, T. Novelties in Macrofungi of the Tropical Montane Cloud Forest in Mexico. J. Fungi 2023, 9, 477. https://doi.org/10.3390/jof9040477
Valenzuela R, Luna-Vega I, Martínez-Pineda M, Martínez-González CR, García-Jiménez J, de la Fuente J, Bautista-Hernández S, Acosta-Castellanos S, Raymundo T. Novelties in Macrofungi of the Tropical Montane Cloud Forest in Mexico. Journal of Fungi. 2023; 9(4):477. https://doi.org/10.3390/jof9040477
Chicago/Turabian StyleValenzuela, Ricardo, Isolda Luna-Vega, Michelle Martínez-Pineda, César Ramiro Martínez-González, Jesús García-Jiménez, Javier de la Fuente, Silvia Bautista-Hernández, Salvador Acosta-Castellanos, and Tania Raymundo. 2023. "Novelties in Macrofungi of the Tropical Montane Cloud Forest in Mexico" Journal of Fungi 9, no. 4: 477. https://doi.org/10.3390/jof9040477
APA StyleValenzuela, R., Luna-Vega, I., Martínez-Pineda, M., Martínez-González, C. R., García-Jiménez, J., de la Fuente, J., Bautista-Hernández, S., Acosta-Castellanos, S., & Raymundo, T. (2023). Novelties in Macrofungi of the Tropical Montane Cloud Forest in Mexico. Journal of Fungi, 9(4), 477. https://doi.org/10.3390/jof9040477