
Molecular Variability of the Fusarium solani Species Complex Associated with Fusarium Wilt of Melon in Iran
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Survey, Sampling, and Fungal Isolation
2.2. Morphological Characterization
2.3. Pathogenicity Tests
2.4. DNA Extraction, PCR Amplification and Phylogenetic Analysis
2.5. Genetic Diversity and Haplotype Network Analysis
3. Results
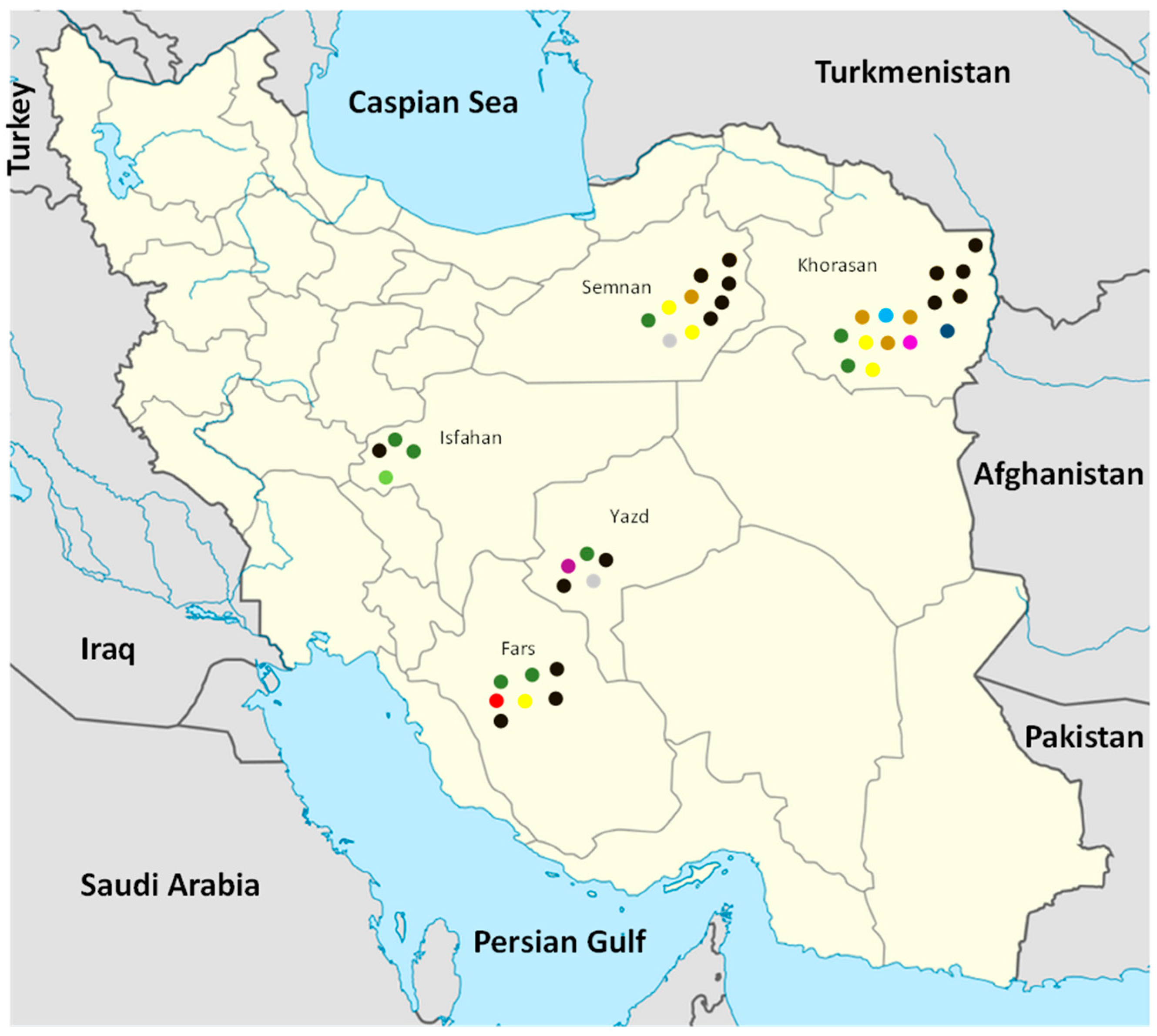
3.1. FSSC Isolates Obtained from Symptomatic Melon Plants in Iran
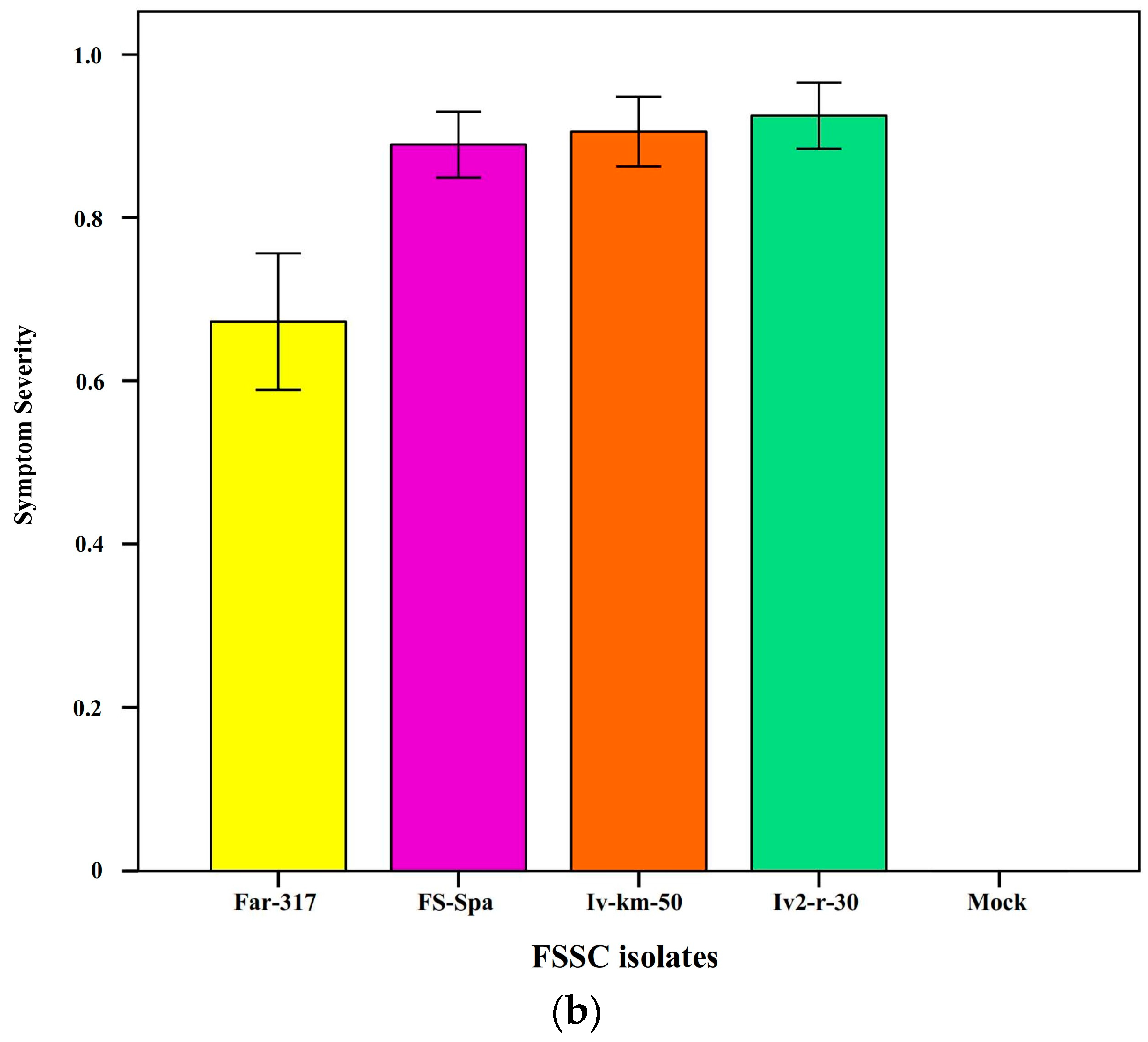
3.2. Pathogenicity and Host Range of FSSC Strains
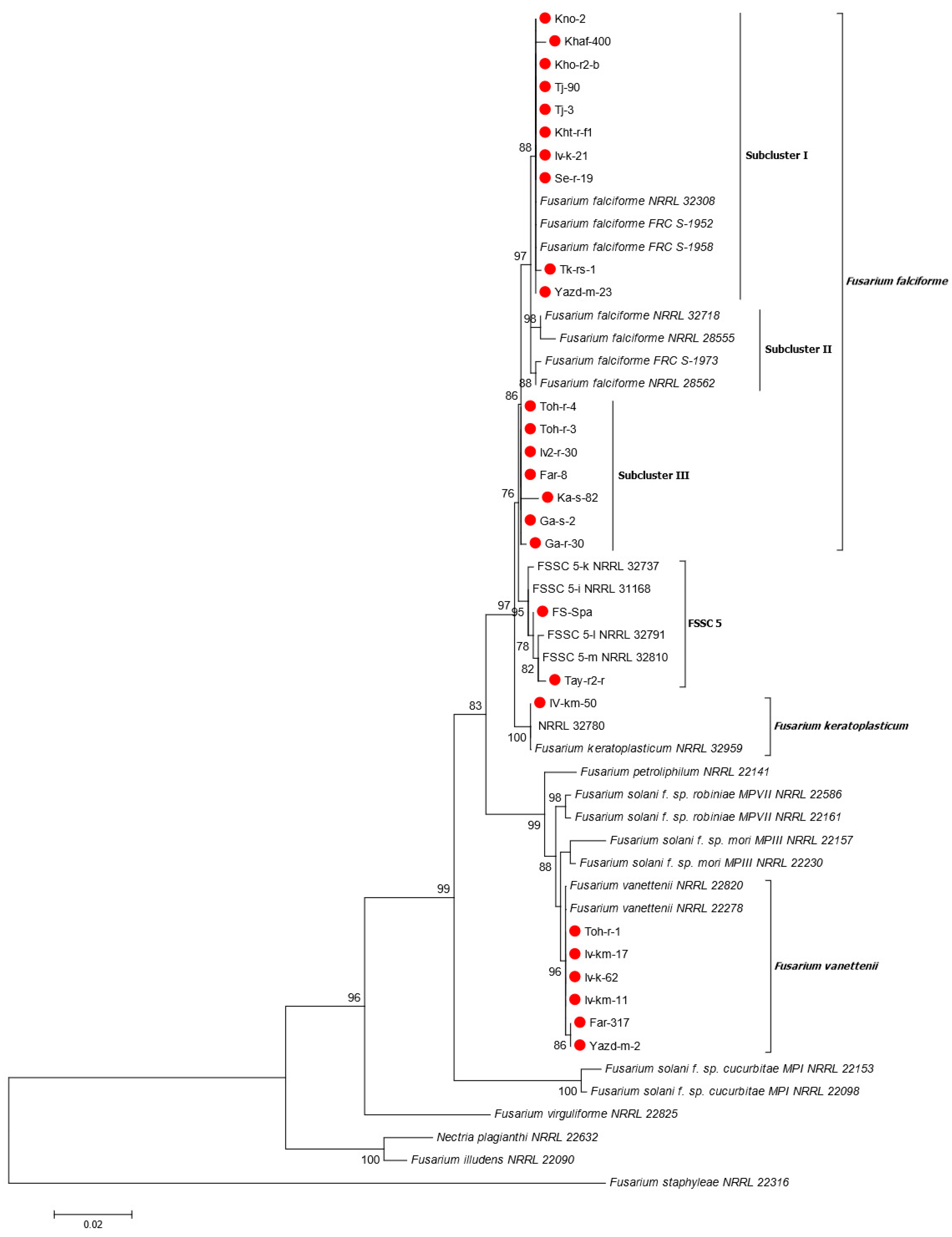
3.3. Phylogenetic Analyses
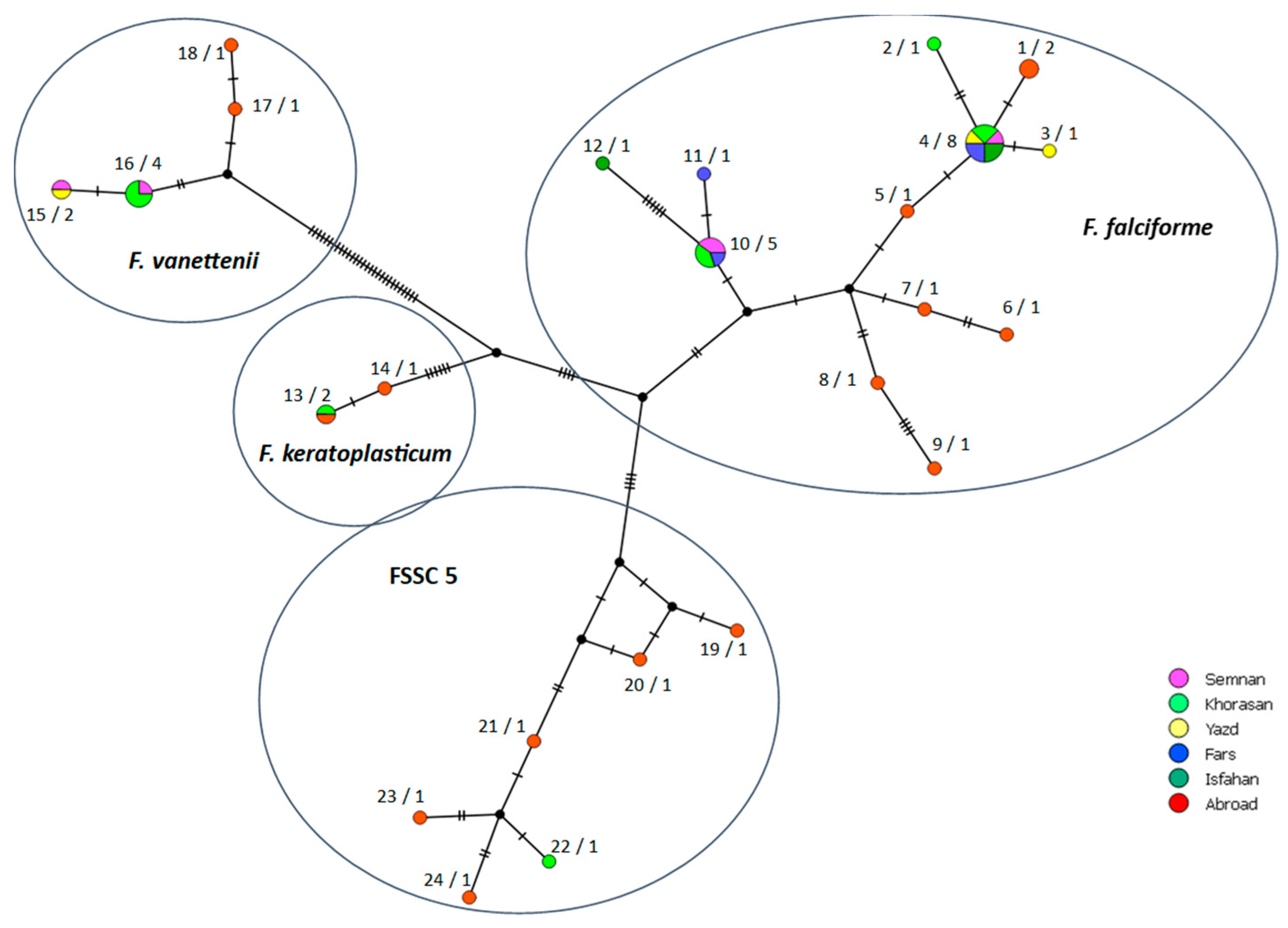
3.4. Genetic Diversity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Flores-León, A.; García-Martínez, S.; González, V.; Garcés-Claver, A.; Martí, R.; Julián, C.; Sifres, A.; Pérez de Castro, A.; Díez, M.J.; Lòpez, C.; et al. Grafting snake melon [Cucumis melo L. subsp. melo Var. flexuosus (L.) Naudin] in organic farming: Effects on agronomic performance; resistance to pathogens; sugar, acid, and VOC profiles; and consumer acceptance. Front. Plant Sci. 2021, 12, 613845. [Google Scholar] [CrossRef] [PubMed]
- Zohary, D.; Hopf, M. Domestication of Plants in the Old World, 3rd ed.; Oxford University Press: New York, NY, USA, 2000; p. 316. [Google Scholar]
- Watson, W. Early cereal cultivation in China. In The Domestication and Exploitation of Plants and Animals; Ucko, P.J., Dimbleby, G.W., Eds.; Gerald Duckworth and Co.: London, UK, 1969; pp. 397–402. [Google Scholar]
- Mallick, M.F.R.; Masui, M. Origin, distribution and taxonomy of melons. Sci. Hortic.-Amsterdam 1986, 28, 251–261. [Google Scholar] [CrossRef]
- Kerje, T.; Grum, M. The origin of melon, Cucumis melo: A review of the literature. Acta Hortic. 2000, 510, 37–44. [Google Scholar] [CrossRef]
- Oumouloud, A.; El-Otmani, M.; Chikh-Rouhou, H.; Claver, A.G.; Torres, R.G.; Perl-Treves, R.; Alvarez, J.M. Breeding melon for resistance to Fusarium wilt: Recent developments. Euphytica 2013, 192, 155–169. [Google Scholar] [CrossRef]
- Banihashemi, Z. The existence of Fusarium wilt of melon in Iran. Proc. First Nat. Cong. Plant Med. Iran 1968, 47–48. [Google Scholar]
- Banihashemi, Z. A new physiological race of Fusarium oxysporum f. sp. melonis in Iran. Iran. J. Plant Pathol. 1982, 18, 1–6. [Google Scholar]
- Sabahi, F.; Banihashemi, Z. Isolation, race determination and molecular characterization of Fusarium oxysporum f. sp. melonis in Bushehr province. Iran. J. Plant Pathol. 2020, 56, 359–369. [Google Scholar]
- Sabahi, F.; de Sain, M.; Banihashemi, Z.; Rep, M. Comparative genomics of Fusarium oxysporum f. sp. melonis strains reveals nine lineages and a new sequence type of AvrFom2. Environ. Microbiol. 2021, 23, 2035–2053. [Google Scholar] [CrossRef]
- Banihashemi, Z. Distribution of mating types of Phytophthora drechsleri the incitant of cucurbit root rot in Fars province. In Proceedings of the 9th Iranian Plant Protection Congress, Karaj, Iran; 1983; p. 102. [Google Scholar]
- Sarpeleh, A. The role of Monosporascus cannonballus in melon collapse in Iran. Australas. Plant Dis. Notes 2008, 3, 162–164. [Google Scholar]
- Sabahi, F.; Mirtalebi, M.; Banihashemi, Z. Paramyrothecium foliicola causes crown dry rot of melon (Cucumis melo) in Iran. J. Phytopathol. 2022, 170, 655–660. [Google Scholar] [CrossRef]
- Mirtalebi, M.; Sabahi, F.; Banihashemi, Z. First report of melon decline caused by Plectosphaerella cucumerina in Iran. Mycol. Iran. 2022, 9, 97–103. [Google Scholar]
- Mirtalebi, M.; Sabahi, F.; Banihashemi, Z. Fruit rot caused by Neoscytalidium hyalinum on melon in Iran. Australas. Plant Dis. Notes 2019, 14, 1–4. [Google Scholar] [CrossRef]
- Homa, M.; Galgóczy, L.; Manikandan, P.; Narendran, V.; Sinka, R.; Csernetics, Á.; Csaba, V.; László, K.; Papp, T. South Indian isolates of the Fusarium solani species complex from clinical and environmental samples: Identification, antifungal susceptibilities, and virulence. Front. Microbiol. 2018, 9, 1052. [Google Scholar] [CrossRef]
- Coleman, J.J. The Fusarium solani species complex: Ubiquitous pathogens of agricultural importance. Mol. Plant Pathol. 2016, 17, 146–158. [Google Scholar] [CrossRef] [PubMed]
- Kolattukudy, P.E.; Gamble, D.L. Nectria haematococca: Pathogenesis and host specificity in plant diseases. In Pathogenesis and Host Specificity in Plant Pathogenic Fungi and Nematodes; Kohmoto, K., Singh, U.S., Singh, R.P., Eds.; Elsevier Science Limited: Amsterdam, The Netherlands, 1995; pp. 83–102. [Google Scholar]
- Gordon, T.R. Fusarium oxysoporum and the Fusarium wilt syndrome. Annu. Rev. Phytopathol. 2017, 55, 23–39. [Google Scholar] [CrossRef] [PubMed]
- Dignani, M.C.; Anaissie, E.J. Human fusariosis. Clin. Microbiol. Infec. 2004, 10, 67–75. [Google Scholar] [CrossRef]
- Zhang, N.; O’Donnell, K.; Sutton, D.A.; Nalim, F.A.; Summerbell, R.C.; Padhye, A.A.; Geiser, D.M. Members of the Fusarium solani species complex that cause infections in both humans and plants are common in the environment. J. Clin. Microbiol. 2006, 44, 2186–2190. [Google Scholar] [CrossRef]
- O’Donnell, K.; Sutton, D.A.; Fothergill, A.; McCarthy, D.; Rinaldi, M.G.; Brandt, M.E.; Zhang, N.; Geiser, D.M. Molecular phylogenetic diversity, multilocus haplotype nomenclature, and in vitro antifungal resistance within the Fusarium solani species complex. J. Clin. Microbiol. 2008, 46, 2477–2490. [Google Scholar] [CrossRef]
- Muhammed, M.; Anagnostou, T.; Desalermos, A.; Kourkoumpetis, T.K.; Carneiro, H.A.; Glavis-Bloom, J.; Cleman, J.J.; Mylonakis, E. Fusarium infection: Report of 26 cases and review of 97 cases from the literature. Medicine 2013, 92, 305. [Google Scholar] [CrossRef]
- Mehl, H.L.; Epstein, L. Fusarium solani species complex isolates conspecific with Fusarium solani f. sp. cucurbitae race 2 from naturally infected human and plant tissue and environmental sources are equally virulent on plants, grow at 37 °C and are interfertile. Environ. Microbiol. 2007, 9, 2189–2199. [Google Scholar] [CrossRef]
- Guadet, J.; Julien, J.; Lafay, J.F.; Brygoo, Y. Phylogeny of some Fusarium species, as determined by large-subunit rRNA sequence comparison. Mol. Biol. Evol. 1989, 6, 227–242. [Google Scholar] [PubMed]
- O’Donnell, K. Progress towards a phylogenetic classification of Fusarium. Sydowia 1996, 48, 57–70. [Google Scholar]
- O’Donnell, K. Molecular phylogeny of the Nectria haematococca-Fusarium solani species complex. Mycologia 2000, 92, 919–938. [Google Scholar] [CrossRef]
- Nalim, F.A.; Samuels, G.J.; Wijesundera, R.L.; Geiser, D.M. New species from the Fusarium solani species complex derived from perithecia and soil in the old world tropics. Mycologia 2011, 103, 1302–1330. [Google Scholar] [CrossRef] [PubMed]
- Aoki, T.; O’Donnell, K.; Homma, Y.; Lattanzi, A.R. Sudden-death syndrome of soybean is caused by two morphologically and phylogenetically distinct species within the Fusarium solani species complex—F. virguliforme in North America and F. tucumaniae in South America. Mycologia 2003, 95, 660–684. [Google Scholar] [CrossRef] [PubMed]
- Al-Hatmi, A.M.; Ahmed, S.A.; Van Diepeningen, A.D.; Drogari-Apiranthitou, M.; Verweij, P.E.; Meis, J.F.; De Hoog, G.S. Fusarium metavorans sp. nov.: The frequent opportunist ‘FSSC6’. Med. Mycol. 2018, 56, S144–S152. [Google Scholar] [CrossRef]
- Schroers, H.J.; Samuels, G.J.; Zhang, N.; Short, D.P.; Juba, J.; Geiser, D.M. Epitypification of Fusisporium (Fusarium) solani and its assignment to a common phylogenetic species in the Fusarium solani species complex. Mycologia 2016, 108, 806–819. [Google Scholar] [CrossRef]
- Short, D.P.G.; O’Donnell, K.; Thrane, U.; Nielsen, K.F.; Zhang, N.; Juba, J.H.; Geiser, D.M. Phylogenetic relationships among members of the Fusarium solani species complex in human infections and the descriptions of F. keratoplasticum sp. nov. and F. petroliphilum stat. nov. Fungal Genet. Biol. 2013, 53, 59–70. [Google Scholar] [CrossRef]
- Summerbell, R.C.; Schroers, H.J. Analysis of phylogenetic relationship of Cylindrocarpon lichenicola and Acremonium falciforme to the Fusarium solani species complex and a review of similarities in the spectrum of opportunistic infections caused by these fungi. J. Clin. Microbiol. 2002, 40, 2866–2875. [Google Scholar] [CrossRef]
- Geiser, D.M.; Aoki, T.; Bacon, C.W.; Baker, S.E.; Bhattacharyya, M.K.; Brandt, M.E.; Brown, D.W.; Burgess, L.W.; Chulze, S.; Coleman, J.J.; et al. One fungus, one name: Defining the genus Fusarium in a scientifically robust way that preserves longstanding use. Phytopathology 2013, 103, 400–408. [Google Scholar] [CrossRef]
- O’Donnell, K.; Ward, T.J.; Robert, V.A.R.G.; Crous, P.W.; Geiser, D.M.; Kang, S. DNA sequence-based identification of Fusarium: Current status and future directions. Phytoparasitica 2015, 43, 583–595. [Google Scholar] [CrossRef]
- Sandoval-Denis, M.; Crous, P.W. Removing chaos from confusion assigning names to common human an animal pathogens in Neocosmospora. Persoonia 2018, 41, 109–129. [Google Scholar] [CrossRef] [PubMed]
- Sandoval-Denis, M.; Lombard, L.; Crous, P.W. Back to the roots: A reappraisal of Neocosmospora. Persoonia 2019, 43, 90–185. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Al-Hatmi, A.M.S.; Aoki, T.; Brankovics, B.; Cano-Lira, J.F.; Coleman, J.J.; de Hoog, S.G.; Di Pietro, A.; Frandsen, R.J.N.; Geiser, D.M.; et al. No to Neocosmospora: Phylogenomic and practical reasons for continued inclusion of the Fusarium solani species complex in the genus Fusarium. mSphere 2020, 5, e00810–e00820. [Google Scholar] [CrossRef]
- Geiser, D.M.; Al-Hatmi, A.M.S.; Aoki, T.; Arie, T.; Balmas, V.; Barnes, I.; Bergstrom, G.C.; Bhattacharyya, M.K.; Blomquist, C.L.; Bowden, R.L.; et al. Phylogenomic analysis of a 55.1 kb 19-gene dataset resolves a monophyletic Fusarium that includes the Fusarium solani species complex. Phytopathology 2021, 111, 1064–1079. [Google Scholar] [CrossRef]
- Guarnaccia, V.; van Niekerk, J.; Crous, P.W.; Sandoval-Denis, M. Neocosmospora spp. associated with dry root rot of citrus in South Africa. Phytopathol. Mediterr. 2021, 60, 79–100. [Google Scholar] [CrossRef]
- Kamali-Sarvestani, S.; Mostowfizadeh-Ghalamfarsa, R.; Salmaninezhad, F.; Cacciola, S.O. Fusarium and Neocosmospora species associated with rot of Cactaceae and other succulent plants. J. Fungi 2022, 8, 364. [Google Scholar] [CrossRef]
- Alymanesh, M.; Falahati, R.M.; Jafarpour, B.; Mahdikhanimoghadam, E. Identification of Fusarium solani f. sp. cucurbitae race 1 on cucurbit plants from Khorasan Razavi, northern Khorasan and some regions of Fars provinces, using classic and molecular methods. Hortic. Sci. 2007, 21, 49–56. [Google Scholar]
- Chehri, K.; Salleh, B.; Yli-Mattila, T.; Reddy, K.R.N.; Abbasi, S. Molecular characterization of pathogenic Fusarium species in cucurbit plants from Kermanshah province, Iran. Saudi J. Biol. Sci. 2011, 18, 341–351. [Google Scholar] [CrossRef]
- Zhian Jahangir, A.; Banihashemi, Z. Etiology of Fusarium Root Rot of Cucurbits with Emphasis on Fusarium solani f. sp. cucurbitae and its Vegetative Compatibility Groups and Host Range. Master’s Thesis in Agrometeorology, Shiraz University, Shiraz, Iran, 2011. [Google Scholar]
- Alymanesh, M.R.; Falahatirastegar, M.; Jafarpour, B.; Mahdikhanimoghadam, E. Genetic diversity in the fungus Fusarium solani f. sp. cucurbitae race 1, the causal agent of root and crown rot of cucurbits in Iran, using molecular markers. Pak. J. Biol. Sci. 2009, 12, 836–843. [Google Scholar] [CrossRef]
- Moradzadeh Eskandari, M.; Javan-Nikkhah, M.; Zare, R.A.S.O.U.L.; Okhowat, M.; Moretti, A.; Somma, S.; Stea, G. Study on genetic structure of Fusarium solani populations isolated from potato, cucurbit, bean and chickpea based on FAFLP markers. Iran. J. Plant Pathol. 2010, 45, 189–197. [Google Scholar]
- Porter, L.D.; Pasche, J.S.; Chen, W.; Harveson, R.M. Isolation, identification, storage, pathogenicity tests, hosts, and geographic range of Fusarium solani f. sp. pisi causing Fusarium root rot of pea. Plant Health Prog. 2015, 16, 136–145. [Google Scholar] [CrossRef]
- Snyder, W.C.; Hansen, H.N. Advantages of natural media and environments in the culture of fungi. Phtopathology 1974, 37, 420–421. [Google Scholar]
- Fisher, N.L.; Burgess, L.W.; Toussoun, T.A.; Nelson, P.E. Carnation leaves as a substrate and for preserving cultures of Fusarium species. Phytopathology 1982, 72, 151–153. [Google Scholar] [CrossRef]
- Pascoe, I.G. Fusarium morphology identification and characterization of a third conidial type, the mesoconidium. Mycotaxon 1990, 37, 121–160. [Google Scholar]
- Nirenberg, H.L. Untersuchungen Uber Die Morphologische Und Biologische Differenzierung in der FUSARIUM-Section Liseola; Kommissionsverlag Paul Parey: Berlin, Germany, 1976; pp. 1–117. [Google Scholar]
- Leslie, J.F.; Summerell, B.A.; Bullock, S. The Fusarium Laboratory Manual; Blackwell Publishing: Ames, IA, USA, 2006; pp. 1–388. [Google Scholar]
- Zhang, X.Y.; Yu, X.X.; Yu, Z.; Xue, Y.F.; Qi, L.P. A simple method based on laboratory inoculum and field inoculum for evaluating potato resistance to black scurf caused by Rhizoctonia solani. Breeding Sci. 2014, 64, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Nagao, H.; Sato, K.; Ogiwara, S. Susceptibility of Cucurbita spp. to the cucurbit root-rot fungus, Fusarium solani f. sp. cucurbitae race 1. Agronomie 1994, 14, 95–102. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, N., Gelfand, D., Sninsky, J., White, T., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- O’Donnell, K. Fusarium and its near relatives. In The Fungal Holomorph: Mitotic, Meiotic and Pleomorphic Speciation in Fungal Systematics; Reynolds, D.R., Taylor, J.W., Eds.; CAB International: Wallingford, UK, 1993; pp. 225–233. [Google Scholar]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef]
- National Library of Medicine. BLAST®. Available online: https://blast.ncbi.nlm.nih.gov/ (accessed on 10 February 2023).
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Iceni Technology. Available online: https://www.iceni.com/infix.htm (accessed on 10 February 2023).
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Clement, M.; Snell, Q.; Walker, P.; Posada, D.; Crandall, K. TCS: Estimating gene genealogies. Int. Parallel Distrib. Process. Symp. 2002, 2, 184. [Google Scholar]
- Leigh, J.W.; Bryant, D. POPART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- González, V.; García-Martínez, S.; Flores-León, A.; Ruiz, J.J.; Picó, B.; Garcés-Claver, A. Neocosmospora keratoplastica, a relevant human fusarial pathogen is found to be associated with wilt and root rot of muskmelon and watermelon crops in Spain: Epidemiological and molecular evidences. Eur. J. Plant Pathol. 2020, 156, 1189–1196. [Google Scholar] [CrossRef]
- González, V.; Ruiz, J.J.; Picó, B.; García-Martínez, S.; Garcés-Claver, A.; Flores-León, A. First report of Neocosmospora falciformis causing wilt and root rot of muskmelon in Spain. Plant Dis. 2020, 104, 1256. [Google Scholar] [CrossRef]
- Mohammadi, H.; Banihashemi, Z. Distribution, pathogenicity and survival of Fusarium spp. the causal agents of chickpea wilt and root rot in the Fars province of Iran. Iran. J. Plant Pathol. 2005, 41, 687–708. [Google Scholar]
- Šišić, A.; Baćanović-Šišić, J.; Al-Hatmi, A.M.S.; Karlovsky, P.; Ahmed, S.A.; Maier, W.; Sybren De Hoog, G.; Finckh, M.R. The ‘forma specialis’ issue in Fusarium: A case study in Fusarium solani f. sp. pisi. Sci. Rep.-UK 2018, 8, 1252. [Google Scholar] [CrossRef]
- Debbarma, R.; Kamil, D.; Maya Bashyal, B.; Choudhary, S.P.; Thokla, P. First report of root rot disease on Solanum lycopersicum L. caused by Fusarium vanettenii in India. J. Phytopathol. 2021, 169, 752–756. [Google Scholar] [CrossRef]
- Chehri, K.; Ghasempour, H.R.; Karimi, N. Molecular phylogenetic and pathogenetic characterization of Fusarium solani species complex (FSSC), the cause of dry rot on potato in Iran. Microb. Pathogenesis 2014, 67, 14–19. [Google Scholar] [CrossRef]
- Backhouse, D.; Burgess, L.W. Mycogeography of Fusarium: Climatic analysis of the distribution within Australia of Fusarium species in section Gibbosum. Mycol. Res. 1995, 99, 1218–1224. [Google Scholar] [CrossRef]
- Burgess, L.W.; Nelson, P.E.; Toussoun, T.A.; Forbes, G.A. Distribution of Fusarium species in sections Roseum, Arthrosporiella, Gibbosum and Discolor recovered from grassland, pasture and pine nursery soils of eastern Australia. Mycologia 1988, 80, 815–824. [Google Scholar] [CrossRef]
- Sangalang, A.E.; Backhouse, D.; Burgess, L.W. Survival and growth in culture of four Fusarium species in relation to occurrence in soils from hot climatic regions. Mycol. Res. 1995, 99, 529–533. [Google Scholar] [CrossRef]
- Saremi, H.; Burgess, L.W.; Backhouse, D. Temperature effects on the relative abundance of Fusarium species in a model plant–soil ecosystem. Soil Biol. Biochem. 1999, 31, 941–947. [Google Scholar] [CrossRef]
- Narouirad, M.; Bakhshi, B.; Kohpalekani, A.J.; Ghasemi, A.; Fazeli, R.M. Response of Iranian melon populations to deficit irrigation condition. Hort. Sci. 2020, 34, 161–169. [Google Scholar]
- Posada, D.; Crandall, K.A. Evaluation of methods for detecting recombination from DNA sequences: Computer simulations. Proc. Natl. Acad. Sci. USA 2001, 98, 13757–13762. [Google Scholar] [CrossRef] [PubMed]
- Pareek, V.; Varma, R. Fusarium solani a dominant seed borne pathogen in seeds of cluster bean grown in Rajasthan. Biosci. Biotech. Res. Commun. 2015, 8, 29–34. [Google Scholar]
- González, V.; Armengol, J.; Garcés-Claver, A. First report of Fusarium petroliphilum causing fruit root of butternut squash in Spain. Plant Dis. 2018, 102, 1662. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Species | Year a | Source | Collection b Locality | Sequence Type (ST) c Detected in: | Multilocus Haplotype (MH) | ||
|---|---|---|---|---|---|---|---|---|
| ITS | LSU | tef1 | ||||||
| Se-r-19 | F. falciforme | 2010 | long melon | Se, Is, Ir | 7 | 2 | 12 | 4 |
| Khaf-400 | F. falciforme | 2009 | long melon | Kha, Kho, Ir | 7 | 2 | 13 | 2 |
| Toh-r-3 | F. falciforme | 2009 | long melon | Toh, Kho, Ir | 7 | 2 | 6 | 10 |
| Toh-r-4 | F. falciforme | 2009 | long melon | Toh, Kho, Ir | 7 | 2 | 6 | 10 |
| Iv-k-21 | F. falciforme | 2011 | long melon | Ey, S, Ir | 7 | 2 | 12 | 4 |
| Iv2-r-30 | F. falciforme | 2009 | long melon | Ey, S, Ir | 7 | 2 | 6 | 10 |
| Yazd-m-23 | F. falciforme | 2010 | cantaloupe | Mey, Y, Ir | 7 | 2 | 12 | 4 |
| Tj-90 | F. falciforme | 2009 | long melon | Toj, Kho, Ir | 7 | 2 | 12 | 4 |
| Tk-rs-1 | F. falciforme | 2011 | cantaloupe | Ta, Y, Ir | 7 | 2 | 14 | 3 |
| Ga-r-30 | F. falciforme | 2009 | cantaloupe | Ga, Fa, Ir | 7 | 2 | 7 | 11 |
| Ga-s-2 | F. falciforme | 2011 | cantaloupe | Ga, Fa, Ir | 7 | 2 | 6 | 10 |
| Ka-s-82 | F. falciforme | 2010 | long melon | Ka, Is, Ir | 7 | 1 | 5 | 12 |
| Tj-3 | F. falciforme | 2010 | long melon | Toj, Kho, Ir | 7 | 2 | 12 | 4 |
| Far-8 | F. falciforme | 2011 | long melon | Fa, S, Ir | 7 | 2 | 6 | 10 |
| Kht-r-f1 | F. falciforme | 2009 | long melon | Khan, Is, Ir | 7 | 2 | 12 | 4 |
| Kno-2 | F. falciforme | 2009 | cantaloupe | Khaf, Fa, Ir | 7 | 2 | 12 | 4 |
| Kho-r2-b | F. falciforme | 2011 | cantaloupe | Khaf, Fa, Ir | 7 | 2 | 12 | 4 |
| Tay-r2-r | FSSC 5 | 2009 | long melon | Tay, Kho, Ir | 1 | 3 | 18 | 22 |
| Iv-k-62 | F. vanettenii | 2009 | long melon | Ey, S, Ir | 3 | 5 | 2 | 16 |
| Far-317 | F. vanettenii | 2009 | long melon | Far, S, Ir | 3 | 5 | 1 | 15 |
| Iv-km-11 | F. vanettenii | 2010 | long melon | Kash, Kho, Ir | 3 | 5 | 2 | 16 |
| Iv-km-17 | F. vanettenii | 2010 | long melon | Kash, Kho, Ir | 3 | 5 | 2 | 16 |
| Toh-r-1 | F. vanettenii | 2009 | long melon | Toh, Kho, Ir | 3 | 5 | 2 | 16 |
| Yazd-m-2 | F. vanettenii | 2010 | long melon | Mey, Y, Ir | 3 | 5 | 1 | 15 |
| Iv-km-50 | F. keratoplasticum | 2009 | long melon | Kash, Kho, Ir | 6 | 4 | 3 | 13 |
| Strains from different countries and sources | ||||||||
| NRRL 32718 | F. falciforme | 2006 | human eye | USA | 7 | 2 | 10 | 8 |
| FRC S-1973 | F. falciforme | 2011 | soil | Aust | 7 | 2 | 9 | 6 |
| FRC S-1958 | F. falciforme | 2011 | soil | Aust | 9 | 2 | 12 | 1 |
| FRC S-1952 | F. falciforme | 2011 | soil | Aust | 9 | 2 | 12 | 1 |
| NRRL 28555 | F. falciforme | 2006 | human wrist | USA | 8 | 2 | 11 | 9 |
| NRRL 28562 | F. falciforme | 2006 | human bone | USA | 7 | 2 | 8 | 7 |
| NRRL 32308 | F. falciforme | 2006 | human foot | SA | 7 | 2 | 12 | 5 |
| FS-Spa | FSSC 5 | ND | ND | Spain | 1 | 3 | 19 | 23 |
| NRRL 31168 | FSSC 5 | 2006 | human toe leukemia | USA | 2 | 3 | 15 | 20 |
| NRRL 32810 | FSSC 5 | 2006 | human eye | USA | 1 | 3 | 16 | 21 |
| NRRL 32737 | FSSC 5 | 2006 | human eye | USA | 2 | 3 | 20 | 19 |
| NRRL 32791 | FSSC 5 | 2006 | human | USA | 2 | 3 | 17 | 24 |
| NRRL 22820 | F. vanettenii | ND | ND | USA | 5 | 5 | 2 | 18 |
| NRRL 22278 | F. vanettenii | ND | ND | USA | 4 | 5 | 2 | 17 |
| NRRL 32780 | F. keratoplasticum | 2006 | sea turtle | USA | 6 | 4 | 3 | 13 |
| NRRL 32959 | F. keratoplasticum | 2006 | human skin | USA | 6 | 4 | 4 | 14 |
| Isolate | Species | Colony Growth Rate (mm/day) | Pigment on PDA a | Chlamydospores b | Shape/No. of Septa of Microconidia a | Shape of Basal and Apical Cell a | Length × Width of Macroconidia (µm) c | |
|---|---|---|---|---|---|---|---|---|
| 3 and 4 Septate | 5 Septate | |||||||
| Se-r19 | F. falciforme | 7.5 | WY or G | + | EO, OT/0-1 | BNP, Cu | 40.5 ± 1.5 × 5.0 ± 0.5 | - |
| Khaf-400 | F. falciforme | 7.5 | WY or G | + | EO, OT/0-1 | BNP, Cu | 41.5 ± 2.5 × 5.1 ± 0.5 | - |
| Toh-r3 | F. falciforme | 9.0 | WY or G | + | EO, OT/0-1 | BNP, Cu | 42.5 ± 2.5 × 5.2 ± 0.5 | - |
| Yazd-m23 | F. falciforme | 8.0 | WY or G | + | EO, OT/0-1 | BNP, Cu | 41.0 ± 1.5 × 5.2 ± 0.5 | - |
| Tj-90 | F. falciforme | 8.5 | WY or G | + | EO, OT/0-1 | BNP, Cu | 42.0 ± 2.5 × 5.1 ± 0.5 | - |
| Tk-rs-1 | F. falciforme | 9.0 | WY or G | + | EO, OT/0-1 | BNP, Cu | 41.5 ± 2.5 × 5.0 ± 0.5 | - |
| Ga-r30 | F. falciforme | 9.0 | WY or G | + | EO, OT/0-1 | BNP, Cu | 40.5 ± 1.5 × 5.0 ± 0.5 | - |
| Ga-s2 | F. falciforme | 8.5 | WY or G | + | EO, OT/0-1 | BNP, Cu | 42.5 ± 2.5 × 5.2 ± 0.5 | - |
| Ka-s-82 | F. falciforme | 9.0 | WY or G | + | EO, OT/0-1 | BNP, Cu | 40.5 ± 1.5 × 5.0 ± 0.5 | - |
| Tj-3 | F. falciforme | 7.0 | WY or G | + | EO, OT/0-1 | BNP, Cu | 41.5 ± 2.5 × 5.1 ± 0.5 | - |
| Far-8 | F. falciforme | 7.5 | WY or G | + | EO, OT/0-1 | BNP, Cu | 41.5 ± 2.5× 5.1 ± 0.5 | - |
| Yazd-m2 | F. falciforme | 8.0 | WY or G | + | EO, OT/0-1 | BNP, Cu | 42.0 ± 2.5 × 5.3 ± 0.5 | - |
| Kho-r2-b | F. falciforme | 8.0 | WY or G | + | EO, OT/0-1 | BNP, Cu | 42.5 ± 2.5 × 5.1 ± 0.5 | - |
| Kht-r-f1 | F. falciforme | 7.5 | WY or G | + | EO, OT/0-1 | BNP, Cu | 42.5 ± 2.5 × 5.2 ± 0.5 | - |
| Toh-r4 | F. falciforme | 7.0 | WY or G | + | EO, OT/0-1 | BNP, Cu | 40.5 ± 1.5 × 5.0 ± 0.5 | - |
| iv-k-21 | F. falciforme | 9.0 | WY or G | + | EO, OT/0-1 | BNP, Cu | 40.5 ± 1.5 × 5.0 ± 0.5 | - |
| iv2-r30 | F. falciforme | 8.5 | WY or G | + | EO, OT/0-1 | BNP, Cu | 41.5 ± 2.5 × 5.1 ± 0.5 | - |
| Kno2 | F. falciforme | 7.0 | WY or G | + | EO, OT/0-1 | BNP, Cu | 41.0 ± 2.5 × 5.2 ± 0.5 | - |
| iv-km-50 | F. keratoplasticum | 8.5 | WY | + | O, PC, S/0-1 | NB | 36.5 ± 2.3 × 4.8 ± 0.3 | - |
| Far-317 | F. venattenii | 5.5 | WB | + | OR and T/0-1 | PR | 46.2 ± 2.0 × 4.35 ± 0.4 | - |
| iv-km-11 | F. venattenii | 6.0 | WB | + | OR and T/0-1 | PR | 45.2 ± 1.5 × 4.35 ± 0.4 | - |
| iv-km-17 | F. venattenii | 6.0 | WB | + | OR and T/0-1 | PR | 46.2 ± 2.5 × 4.33 ± 0.1 | - |
| Toh-r1 | F. venattenii | 5.0 | WB | + | OR and T/0-1 | PR | 44.1 ± 2.3 × 4.34 ± 0.2 | - |
| iv-k-62 | F. venattenii | 5.5 | WB | + | OR and T/0-1 | PR | 44.2 ± 2.0 × 4.35 ± 0.3 | - |
| Tay-r2-r | FSSC 5 | 8.0 | WY or G | + | EO, OT/0-1 | BNP, Cu | 40.5 ± 1.5 × 5.1 ± 0.5 | - |
| FS-Spa | FSSC 5 | 8.5 | WC | + | O, EO, C, R/0-1 | BN, P Cu | 38.6 ± 1.5 × 5.5 ± 0.3 | 42.5 ± 2.5 × 5.8 ± 0.3 |
| No. of | Value for Indicated Neutrality Test b | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Region, FSSC Species | Gene | Strains | Nucleotides | Haplotypes | Haplotype Frequency a | Number of Segregating Sites | Polymorphic Sites (%) | Nucleotide Diversity (π) | Number of Mutation (ղ) | Haplotype (Gene) Diversity | Tajima’s D | Fu and Li’s D* | Fu and Li’s F* | Minimum No. of Recombination Events b |
| F. falciforme | Concatenated | 24 | 1556 | 12 | 0.50 | 24 | 1.542 | 0.00262 | 24 | 0.859 | −1.47289 NS | −2.27745 NS | −2.37548 NS | 0 |
| ITS | 24 | 439 | 4 | 0.166 | 3 | 0.683 | 0.00181 | 3 | 0.634 | −0.12171 NS | −0.18894 NS | −0.19630 NS | 0 | |
| tef1 | 24 | 668 | 10 | 0.416 | 18 | 2.694 | 0.00452 | 18 | 0.764 | −1.46801 NS | −2.18497 NS | −2.29803 NS | 0 | |
| LSU | 24 | 435 | 2 | 0.083 | 3 | 0.689 | 0.00059 | 3 | 0.083 | −1.73253 NS | −2.52572* (p < 0.05) | −2.65835* (p < 0.05) | 0 | |
| F. keratoplasticum | Concatenated | 3 | 1556 | 2 | 0.666 | 1 | 0.064 | 0.00045 | 1 | 0.667 | NA | NA | NA | 0 |
| ITS | 3 | 439 | 1 | 0.333 | 0 | 0.000 | 0.000 | 0 | 0.00 | NA | NA | NA | 0 | |
| tef1 | 3 | 668 | 2 | 0.666 | 1 | 0.149 | 0.00105 | 1 | 0.667 | NA | NA | NA | 0 | |
| LSU | 3 | 435 | 1 | 0.333 | 0 | 0.000 | 0.000 | 0 | 0.000 | NP | NP | NP | NP | |
| FSSC 5 | Concatenated | 6 | 1556 | 6 | 1.000 | 10 | 0.642 | 0.00297 | 10 | 1.000 | 0.11945 NS | 0.7918 NS | 0.9479 NS | 2 |
| ITS | 6 | 439 | 2 | 0.333 | 1 | 0.227 | 0.00143 | 1 | 0.600 | 1.44510 NS | 1.05247 NS | 1.15768 NS | 0 | |
| tef1 | 6 | 668 | 6 | 1.000 | 9 | 1.347 | 0.00595 | 9 | 1.000 | −0.11324 NS | −0.09221 NS | −0.10424 NS | 1 | |
| LSU | 6 | 435 | 1 | 0.166 | 0 | 0.000 | 0.000 | 0 | 0.000 | NP | NP | NP | NP | |
| F. vanettenii | Concatenated | 8 | 1556 | 4 | 0.50 | 5 | 0.321 | 0.00131 | 5 | 0.750 | 0.08445 NS | 0.74709 NS | 0.65390 NS | 0 |
| ITS | 8 | 439 | 3 | 0.375 | 4 | 0.911 | 0.00367 | 4 | 0.464 | −0.01957 NS | 0.56807 NS | 0.47502 NS | 0 | |
| Tef-1α | 8 | 668 | 2 | 0.25 | 1 | 0.149 | 0.00066 | 1 | 0.429 | 0.33350 NS | 0.88779 NS | 0.82528 NS | 0 | |
| LSU | 8 | 435 | 1 | 0.125 | 0 | 0.000 | 0.000 | 0 | 0.000 | NP | NP | NP | NP | |
| Iran | ||||||||||||||
| F. falciforme | Concatenated | 17 | 1556 | 6 | 0.352 | 14 | 0.899 | 0.00218 | 14 | 0.721 | −0.83607 NS | −1.90361 NS | −1.85065 NS | 0 |
| ITS | 17 | 439 | 2 | 0.117 | 1 | 0.227 | 0.00122 | 1 | 0.515 | 1.43020 NS | 0.67700 NS | 0.99442 NS | 0 | |
| tef1 | 17 | 668 | 6 | 0.352 | 10 | 1.497 | 0.00372 | 10 | 0.721 | −0.73074 NS | −1.30215 NS | −1.68143 NS | 0 | |
| LSU | 17 | 435 | 2 | 0.117 | 3 | 0.689 | 0.00083 | 3 | 0.118 | −1.70573 NS | −2.25481 NS | −2.41419 NS | 0 | |
| Non-Iran | ||||||||||||||
| F. falciforme | Concatenated | Concatenated | 7 | 1556 | 6 | 0.857 | 12 | 0.771 | 0.00313 | 12 | 0.952 | −0.25712 NS | −0.07864 NS | −0.13028 NS |
| ITS | ITS | 7 | 439 | 3 | 0.428 | 3 | 0.683 | 0.00291 | 3 | 0.667 | 0.05031 NS | 0.38925 NS | 0.33832 NS | |
| tef1 | 7 | 668 | 5 | 0.714 | 9 | 1.347 | 0.00542 | 9 | 0.857 | −0.35433 NS | −0.25556 NS | −0.30404 NS | 0 | |
| LSU | 7 | 435 | 1 | 0.142 | 0 | 0.000 | 0.000 | 0 | 0.000 | NP | NP | NP | NP | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sabahi, F.; Banihashemi, Z.; Mirtalebi, M.; Rep, M.; Cacciola, S.O. Molecular Variability of the Fusarium solani Species Complex Associated with Fusarium Wilt of Melon in Iran. J. Fungi 2023, 9, 486. https://doi.org/10.3390/jof9040486
Sabahi F, Banihashemi Z, Mirtalebi M, Rep M, Cacciola SO. Molecular Variability of the Fusarium solani Species Complex Associated with Fusarium Wilt of Melon in Iran. Journal of Fungi. 2023; 9(4):486. https://doi.org/10.3390/jof9040486
Chicago/Turabian StyleSabahi, Fatemeh, Zia Banihashemi, Maryam Mirtalebi, Martijn Rep, and Santa Olga Cacciola. 2023. "Molecular Variability of the Fusarium solani Species Complex Associated with Fusarium Wilt of Melon in Iran" Journal of Fungi 9, no. 4: 486. https://doi.org/10.3390/jof9040486
APA StyleSabahi, F., Banihashemi, Z., Mirtalebi, M., Rep, M., & Cacciola, S. O. (2023). Molecular Variability of the Fusarium solani Species Complex Associated with Fusarium Wilt of Melon in Iran. Journal of Fungi, 9(4), 486. https://doi.org/10.3390/jof9040486