
Beech Leaf Disease Severity Affects Ectomycorrhizal Colonization and Fungal Taxa Composition

 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Site Description
2.2. Sampling Methods
2.3. Root Processing
2.4. Molecular Methods
2.5. Pipeline
2.6. Statistical Analysis
2.6.1. ECM Root Colonization Analysis
2.6.2. High-Throughput Sequence Analysis
2.6.3. Wald Tests
3. Results
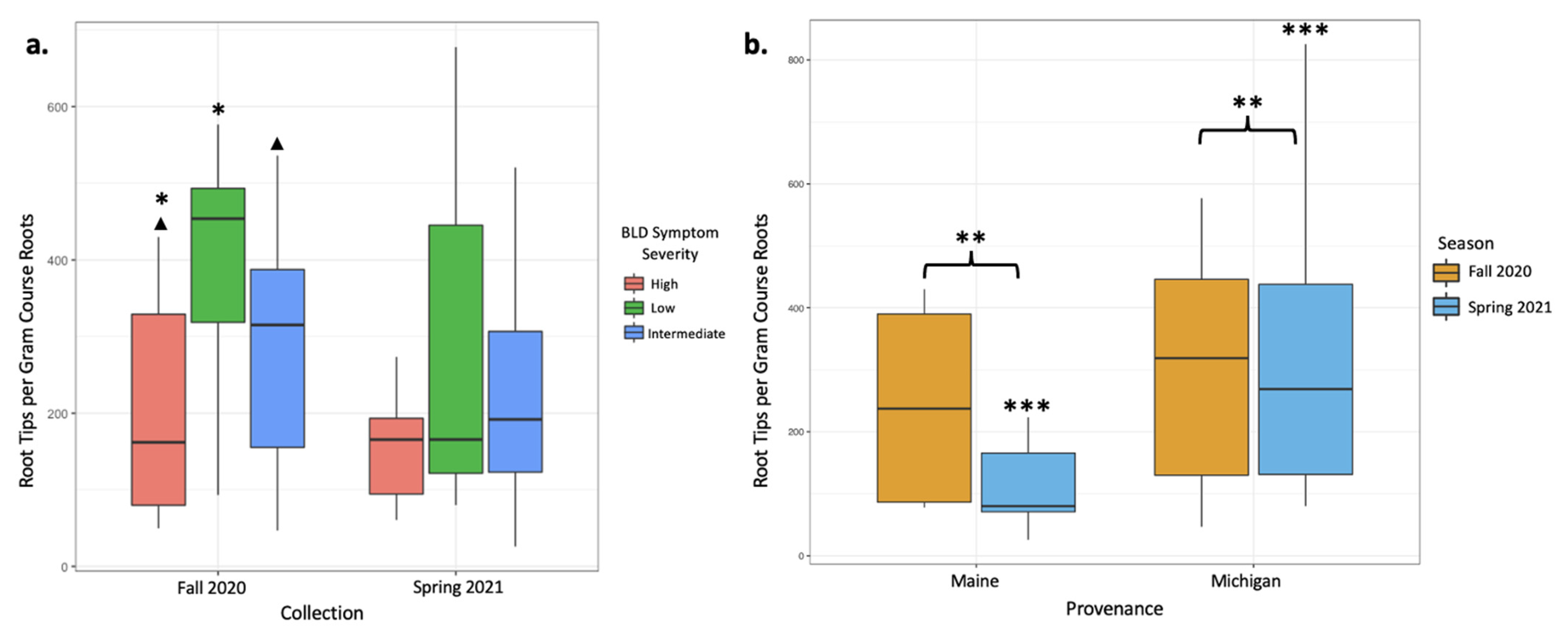
3.1. Root Colonization by ECM Fungi
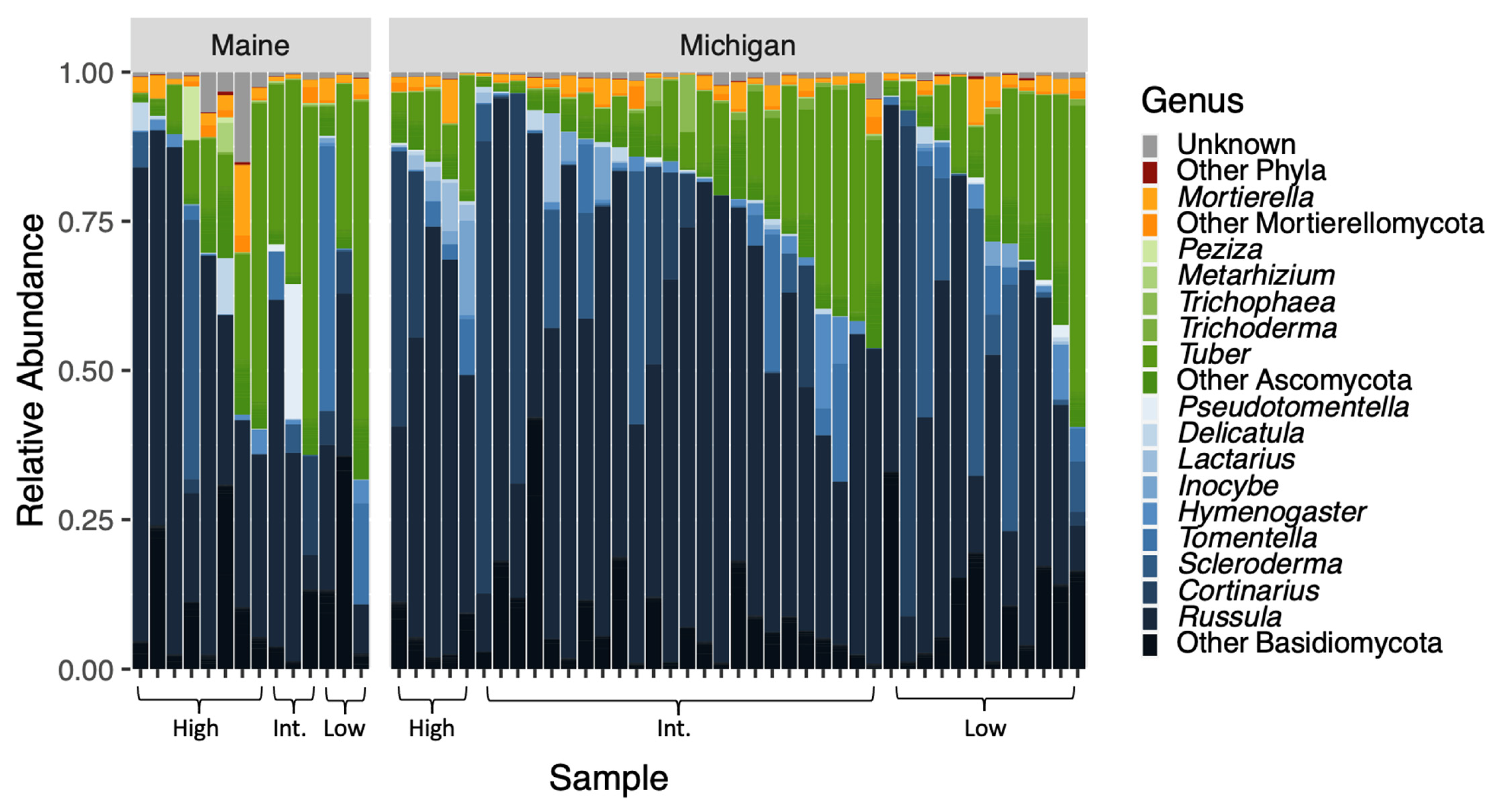
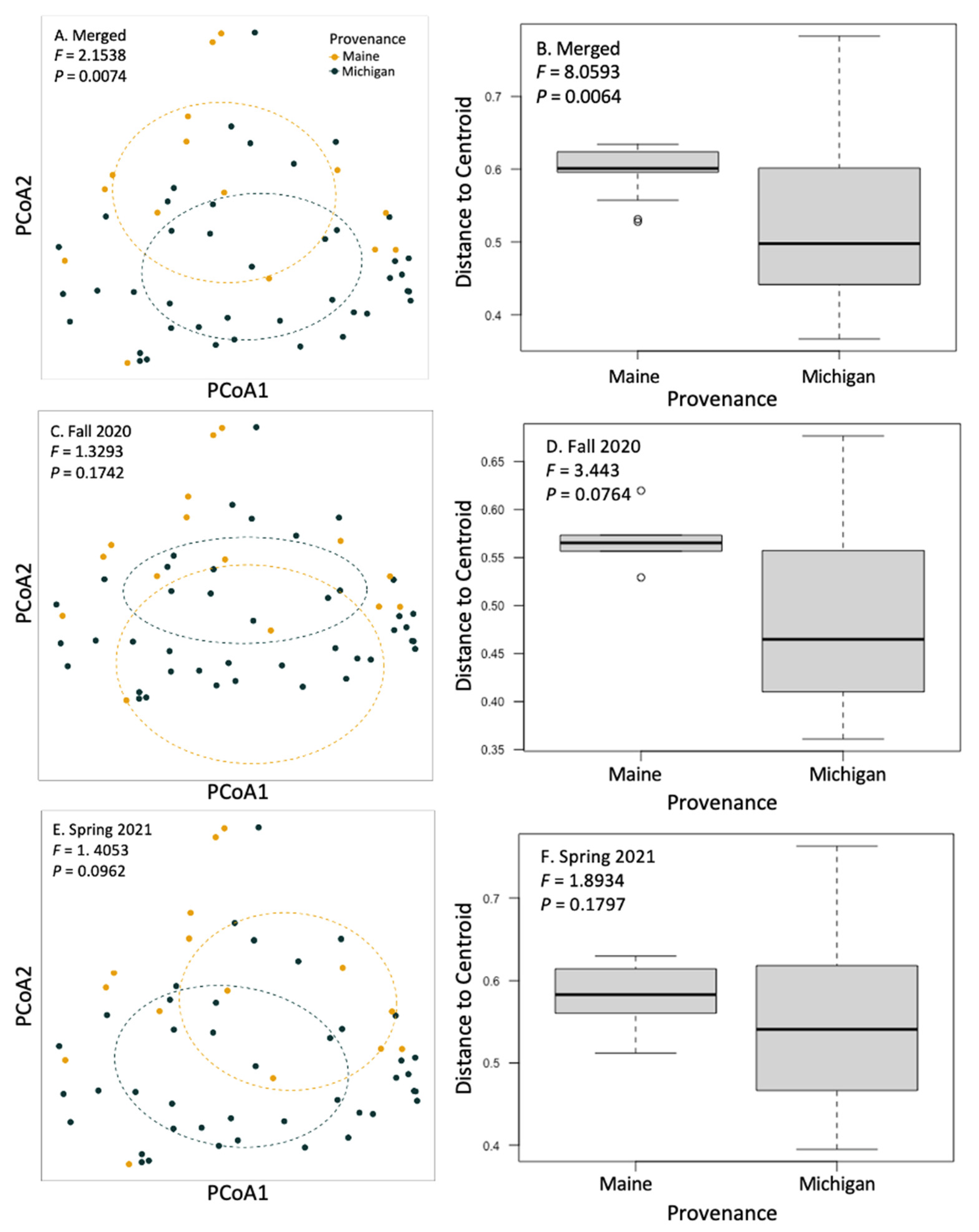
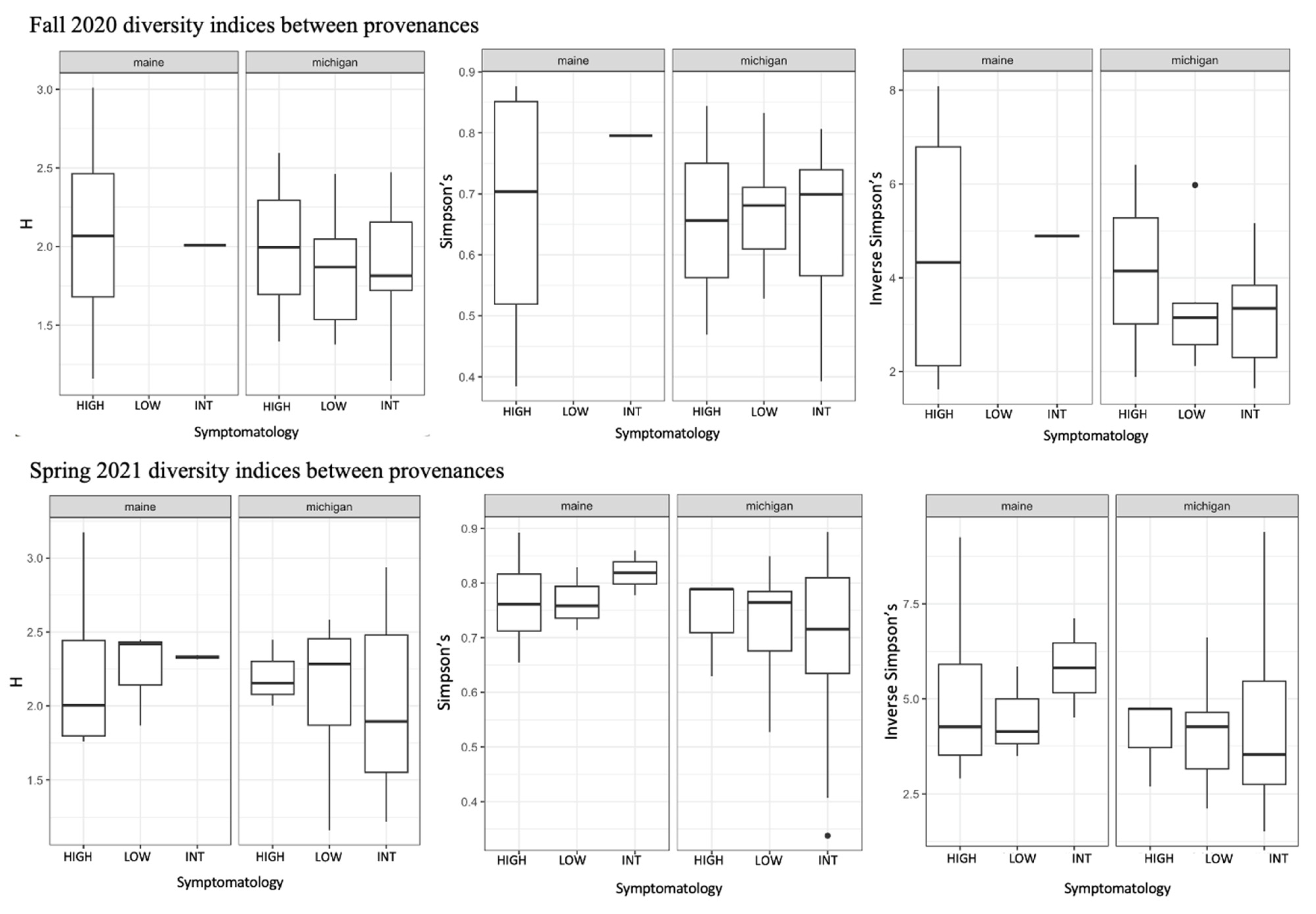
3.2. Community Composition and Diversity of Root Fungi
3.3. Wald Tests of zOTU Abundance by Provenance and BLD Symptomatology
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ewing, C.J.; Hausman, C.E.; Pogacnik, J.; Slot, J.; Bonello, P. Beech leaf disease: An emerging forest epidemic. For. Pathol. 2018, 49, e12488. [Google Scholar] [CrossRef]
- Carta, L.; Handoo, Z.; Li, S.; Burke, D. Beech leaf disease symptoms caused by newly recognized nematode subspecies Litylenchus crenatae mccannii (Anguinata) described from Fagus grandifolia in North America. For. Pathol. 2020, 50, e12580. [Google Scholar] [CrossRef]
- Burke, D.; Hoke, A.; Koch, J. The emergence of beech leaf disease in Ohio: Probing the plant microbiome in search of the cause. For. Pathol. 2020, 50, e12579. [Google Scholar] [CrossRef]
- Bose, A.K.; Weiskittel, A.; Wagner, R.G. A three-decade assessment of climate-associated changes in forest composition across the north-eastern USA. J. Appl. Ecol. 2017, 54, 1592–1604. [Google Scholar] [CrossRef]
- Reed, S.E.; Volk, D.; Martin, D.K.H.; Hausman, C.E.; Macy, T.; Tomon, T.; Cousins, S. The distribution of beech leaf disease and the causal agents of beech bark disease (Cryptoccocus fagisuga, Neonectria faginata, N. ditissima) in forests surrounding Lake Erie and future implications. For. Ecol. Manag. 2023, 503, 119753. [Google Scholar] [CrossRef]
- Finlay, R.D. Ecological aspects of mycorrhizal symbiosis: With special emphasis on the functional diversity of interactions involving the extraradical mycelium. J. Exp. Bot. 2008, 59, 1115–1126. [Google Scholar] [CrossRef]
- Martin, F.; Kohler, A.; Murat, C.; Veneault-Fourrey, C.; Hibbett, D.S. Unearthing the roots of ectomycorrhizal symbioses. Nat. Rev. Microbiol. 2016, 14, 760–773. [Google Scholar] [CrossRef]
- Tresender, K.; Torn, M.; Masiello, C. An ecosystem-scale radiocarbon tracer to test use of litter carbon by ectomycorrhizal fungi. Soil Biol. Biochem. 2006, 38, 1077–1082. [Google Scholar] [CrossRef]
- Burke, D.J.; Smemo, K.A.; Hewins, C.R. Ectomycorrhizal fungi isolated from old-growth northern hardwood forest display variability in extracellular enzyme activity in the presence of plant litter. Soil Biol. Biochem. 2014, 68, 219–222. [Google Scholar] [CrossRef]
- Vrålstad, T.; Schumacher, T.; Taylor, A.F.S. Mycorrhizal synthesis between fungal strains of the Hymenoscyphus ericae aggregate and potential ectomycorrhizal and ericoid hosts. New Phytol. 2002, 153, 143–152. [Google Scholar] [CrossRef]
- Peterson, R.L.; Massicotte, H.B. Exploring structural definitions of mycorrhizas, with emphasis on nutrient-exchange interfaces. Can. J. Bot. 2004, 82, 1074–1088. [Google Scholar] [CrossRef]
- Luo, Z.-B.; Janz, D.; Jiang, X.; Gobel, C.; Wildhagen, H.; Tan, Y.; Rennenberg, H.; Feussner, I.; Polle, A. Upgrading Root Physiology for Stress Tolerance by Ectomycorrhizas: Insights from Metabolite and Transcriptional Profiling into Reprogramming for Stress Anticipation. Plant Physiol. 2009, 151, 1902–1917. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, H.; Gao, J.; Zhang, Y.; Liu, Y.; Tang, M. Effects of ectomycorrhizal fungi (Suillus variegatus) on the growth, hydraulic function, and non-structural carbohydrates of Pinus tabulaeformis under drought stress. BMC Plant Biol. 2021, 21, 171. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.; Wang, C.; Li, Z.; Wang, H.; Xiao, Y.; Chen, J.; Tang, M. The dark septate endophytes and ectomycorrhizal fungi effect on Pinus tabulaeformis Carr. seedling growth and their potential effects to pine wilt disease resistance. Forests 2019, 10, 140. [Google Scholar] [CrossRef]
- Gehring, C.A.; Whitham, T.G. Comparisons of ectomycorrhizae on pinyon pines (Pinus edulis; Pinaceae) across extremes of soil type and herbivory. Am. J. Bot. 1994, 81, 1509–1516. [Google Scholar] [CrossRef]
- Gehring, C.A.; Whitham, T.G. Mycorrhizae-Herbivore Interactions: Population and Community Consequences. In Mycorrhizal Ecology; Springer: Berlin/Heidelberg, Germany, 2002; pp. 295–320. [Google Scholar]
- Vendettuoli, J.F.; Orwig, D.A.; Krumins, J.A.; Waterhouse, M.D.; Preisser, E.L. Hemlock woolly adelgid alters fine root bacterial abundance and mycorrhizal associations in eastern hemlock. For. Ecol. Manag. 2015, 339, 112–116. [Google Scholar] [CrossRef]
- Lewis, J.D.; Licitra, J.; Tuininga, A.R.; Sirulnik, A.; Turner, G.D.; Johnson, J. Oak seedling growth and ectomycorrhizal colonization are less in eastern hemlock stands infested with hemlock woolly adelgid than in adjacent oak stands. Tree Physiol. 2008, 28, 629–636. [Google Scholar] [CrossRef]
- Chu, H.; Wang, C.; Wang, H.; Chen, H.; Tang, M. Pine wilt disease alters soil properties and root-associated fungal communities in Pinus tabulaeformis forest. Plant Soil 2016, 404, 237–249. [Google Scholar] [CrossRef]
- Chu, H.; Tang, M.; Wang, H.; Wang, C. Pinewood nematode infection alters root mycoflora of Pinus tabulaeformis Carr. J. Appl. Microbiol. 2018, 125, 554–563. [Google Scholar] [CrossRef]
- Cale, J.A.; Garrison-Johnston, M.T.; Teale, S.A.; Castello, J.D. Beech bark disease in North America: Over a century of research revisited. For. Ecol. Manag. 2017, 394, 86–103. [Google Scholar] [CrossRef]
- Pena, R.; Offermann, C.; Simon, J.; Naumann, P.S.; Geßler, A.; Holst, J.; Dannenmann, M.; Mayer, H.; Kögel-Knabner, I.; Rennenberg, H.; et al. Girdling Affects Ectomycorrhizal Fungal (EMF) Diversity and Reveals Functional Differences in EMF Community Composition in a Beech Forest. Appl. Environ. Microbiol. 2010, 76, 1831–1841. [Google Scholar] [CrossRef] [PubMed]
- Courty, P.-E.; Buèe, M.; Diedhiou, A.G.; Frey-Klett, P.; Le Tacon, F.; Rineau, F.; Turpault, M.-P.; Uroz, S.; Garbaye, J. The role of ectomycorrhizal communities in forest ecosystem processes: New perspectives and emerging concepts. Soil Biol. Biochem. 2010, 42, 679–698. [Google Scholar] [CrossRef]
- Agerer, R. Exploration types of ectomycorrhizae: A proposal to classify ectomycorrhizal mycelial systems according to their patterns of differentiation and putative ecological importance. Mycorrhiza 2001, 11, 107–114. [Google Scholar] [CrossRef]
- Lilleskov, E.A.; Hobbie, E.A.; Horton, T.R. Conservation of ectomycorrhizal fungi: Exploring the linkages between functional and taxonomic responses to anthropogenic N deposition. Fungal Ecol. 2011, 4, 174–183. [Google Scholar] [CrossRef]
- Johnson, N.C.; Wilson, G.W.T.; Bowker, M.A.; Wilson, J.A.; Miller, R.M. Resource limitation is a driver of local adaptation in mycorrhizal symbioses. Proc. Natl. Acad. Sci. USA 2010, 107, 2093–2098. [Google Scholar] [CrossRef] [PubMed]
- Rúa, M.A.; Antoninka, A.; Antunes, P.M.; Chaudhary, V.B.; Gehring, C.; Lamit, L.J.; Piculell, B.J.; Bever, J.D.; Zabinski, C.; Meadow, J.F.; et al. Home-field advantage? Evidence of local adaptation among plants, soil, and arbuscular mycorrhizal fungi through meta-analysis. BMC Evol. Biol. 2016, 16, 122. [Google Scholar] [CrossRef] [PubMed]
- Koch, J.L. Beech bark disease: The oldest “new” threat to American beech in the United States. Outlooks Pest Manag. 2010, 21, 64–68. [Google Scholar] [CrossRef]
- Koch, J.; Allmaras, M.; Barnes, S.; Berrang, P.; Hall, T.; Iskra, A.; Kochenderfer, J.; Macdonald, W.; Rogers, S.; Rose, J. Beech seed orchard development: Identification and propagation of beech bark resistant American beech trees. In Forest Health Monitoring: National Status, Trends and Analysis; General Technical Report SRS-209; Potter, K.M., Conkling, B.L., Eds.; U.S. Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2015; Chapter 8; pp. 103–108. [Google Scholar]
- Burke, D.J. Effects of Alliaria petiolata (garlic mustard: Brassicaceae) on mycorrhizal colonization and community structure in three herbaceous plants in a mixed deciduous forest. Am. J. Bot. 2008, 95, 1416–1425. [Google Scholar] [CrossRef]
- Martin, K.J.; Rygiewicz, P.T. Fungal-specific PCR primers developed for analysis of the ITS region of environmental DNA extracts. BMC Microbiol. 2005, 5, 28. [Google Scholar] [CrossRef]
- Yang, R.-H.; Su, J.-H.; Shang, J.-J.; Wu, Y.-Y.; Li, Y.; Bao, D.-P.; Yao, Y.-J. Evaluation of the ribosomal DNA internal transcribed spacer (ITS), specifically ITS1 and ITS2, for the analysis of fungal diversity by deep sequencing. PLoS ONE 2018, 13, e0206428. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UNOISE2: Improved error-correction for Illumina 16S and ITS amplicon sequencing. bioRxiv 2016. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet.J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Edgar, R.C. SINTAX: A simple non-Bayesian taxonomy classifier for 16S and ITS sequences. bioRxiv 2016. [Google Scholar] [CrossRef]
- UNITE Community. UNITE USEARCH/UTAX Release for Fungi; Version 18.11.2018; UNITE Community: Tartu, Estonia, 2019. [Google Scholar]
- R Development Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: http://www.r-project.org/ (accessed on 1 February 2023).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. _nlme: Linear and Nonlinear Mixed Effects Models_. R Package Version 3.1-150. 2020. Available online: https://CRAN.R-project.org/package=nlme (accessed on 1 February 2023).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. R Package Version 2.5-6. 2019. Available online: https://CRAN.R-project.org/package=vegan (accessed on 1 February 2023).
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Zhu, A.; Ibrahim, J.G.; Love, M.I. Heavy-tailed prior distributions for sequence count data: Removing the noise and preserving large differences. Bioinformatics 2018, 35, 2084–2092. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Elsevier: New York, NY, USA, 2008; pp. 1–800. [Google Scholar]
- Nabity, P.D.; Zavala, J.A.; DeLucia, E.A. Indirect suppression of photosynthesis on individual leaves by arthropod herbivory. Ann. Bot. 2009, 103, 655–663. [Google Scholar] [CrossRef]
- Welsh, A.; Burke, D.; Hamerlynck, E.; Hahn, D. Seasonal analyses of arbuscular mycorrhizae, nitrogen-fixing bacteria and growth performance of the salt marsh grass Spartina patens. Plant Soil. 2009, 330, 251–266. [Google Scholar] [CrossRef]
- Hewins, C.; Carrino-Kyker, S.; Burke, D. Seasonal variation in mycorrhizal fungi colonizing roots of Allium tricoccum (wild leek) in a mature mixed hardwood forest. Mycorrhiza 2015, 25, 469–483. [Google Scholar] [CrossRef] [PubMed]
- Tierney, G.L.; Fahey, T.J.; Groffman, P.M.; Hardy, J.P.; Fitzhugh, R.D.; Driscoll, C.T.; Yavitt, J.B. Environmental control of fine root dynamics in a northern hardwood forest. Global Chang. Biol. 2003, 9, 670–679. [Google Scholar] [CrossRef]
- Swaty, R.L.; Deckert, R.J.; Whitham, T.G.; Gehring, C.A. Ectomycorrhizal abundance and community composition shifts with drought: Predictions from tree rings. Ecology 2004, 85, 1072–1084. [Google Scholar] [CrossRef]
- Burke, D.; Carrino-Kyker, S.; Burns, J. Is it climate or chemistry? Soil fungal communities respond to soil nutrients in a multi-year high-resolution analysis. Ecosphere 2019, 10, e02896. [Google Scholar] [CrossRef]
- Carrino-Kyker, S.; Coyle, K.; Kluber, L.; Burke, D. Fungal and Bacterial Communities Exhibit Consistent Responses to Reversal of Soil Acidification and Phosphorus Limitation over Time. Microorganisms 2019, 8, 1. [Google Scholar] [CrossRef]
- Rubini, A.; Riccioni, C.; Belfiori, B.; Paolocci, F. Impact of the competition between mating types on the cultivation of Tuber melanosporum: Romeo and Juliet and the matter of space and time. Mycorrhiza 2014, 24, 19–27. [Google Scholar] [CrossRef]
- Pánková, H.; Münzbergová, Z.; Rydlová, J.; Vosátka, M. Co-Adaptation of Plants and Communities of Arbuscular Mycorrhizal Fungi to Their Soil Conditions. Folia Geobot. 2014, 49, 521–540. [Google Scholar] [CrossRef]
- Hoeksema, J.D.; Hernandez, J.V.; Rogers, D.L.; Mendoza, L.L.; Thompson, J.N. Geographic divergence in a species-rich symbiosis: Interactions between Monterey pines and ectomycorrhizal fungi. Ecology 2012, 93, 2274–2285. [Google Scholar] [CrossRef]
- Rúa, M.A.; Lamit, L.J.; Gehring, C.; Antunes, P.M.; Hoeksema, J.D.; Zabinski, C.; Karst, J.; Burns, C.; Woods, M. Accounting for local adaptation in ectomycorrhizas: A call to track geographical origin of plants, fungi, and soils in experiments. Mycorrhiza 2018, 28, 187–195. [Google Scholar] [CrossRef]
- Schelkle, M.; Peterson, R.L. Suppression of common root pathogens by helper bacteria and ectomycorrhizal fungi in vitro. Mycorrhiza 1996, 6, 481–485. [Google Scholar] [CrossRef]
- Kanekar, S.S.; Cale, J.A.; Erbilgin, N. Ectomycorrhizal fungal species differentially affect the induced defensive chemistry of lodgepole pine. Oecologia 2018, 188, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Vishwanathan, K.; Zienkiewicz, K.; Liu, Y.; Janz, D.; Feussner, I.; Polle, A.; Haney, C.H. Ectomycorrhizal fungi induce systemic resistance against insects on a nonmycorrhizal plant in a CERK1-dependent manner. New Phytol. 2020, 228, 728–740. [Google Scholar] [CrossRef] [PubMed]
- Dreischhoff, S.; Das, I.S.; Jakobi, M.; Kasper, K.; Polle, A. Local responses and systemic induced resistance mediated by ectomycorrhizal fungi. Front. Plant Sci. 2020, 11, 590063. [Google Scholar] [CrossRef]
- Chu, H.; Wang, H.; Zhang, Y.; Li, Z.; Wang, C.; Dai, D.; Tang, M. Inoculation with ectomycorrhizal fungi and dark septate endophytes contributes to the resistance of Pinus spp. to pine wilt disease. Front. Microbiol. 2021, 12, 687304. [Google Scholar] [CrossRef] [PubMed]
- Costanza, R.; d’Arge, R.; de Groot, R.; Farber, S.; Grasso, M.; Hannon, B.; Limburg, K.; Naeem, S.; O’Niell, R.V.; Paruelo, J.; et al. The value of the world’s ecosystem services and natural capital. Nature 1997, 387, 253–260. [Google Scholar] [CrossRef]
- Ferlian, O.; Goldmann, K.; Eisenhauer, N.; Tarkka, M.T.; Buscot, F.; Heintz-Buschart, A. Distinct effects of host and neighbour tree identity on arbuscular and ectomycorrhizal fungi along a tree diversity gradient. ISME Commun. 2021, 1, 40. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fall Collection Date | Spring Collection Date | Tree ID | Provenance | Symptomatology |
|---|---|---|---|---|
| 16 October 2020 | 15 June 2021 | 57-C | Michigan | Intermediate |
| 16 October 2020 | 28 June 2021 | 57-G | Michigan | Intermediate |
| 1 October 2020 | 29 June 2021 | 57-L | Michigan | Intermediate |
| 1 October 2020 | 14 June 2021 | 57-M | Michigan | High |
| 16 October 2020 | 29 June 2021 | 57-N | Michigan | Low |
| 16 October 2020 | 29 June 2021 | 57-Q | Michigan | Low |
| 1 October 2020 | 15 June 2021 | 57-S1 | Michigan | Intermediate |
| 16 October 2020 | 14 June 2021 | 58-A | Michigan | Intermediate |
| 1 October 2020 | 29 June 2021 | 58-D | Michigan | Low |
| 16 October 2020 | 15 June 2021 | 58-E | Michigan | Low |
| 16 October 2020 | 15 June 2021 | 59-A | Michigan | Intermediate |
| NA | 14 June 2021 | 61-C | Maine | Low |
| NA | 29 June 2021 | 61-D | Maine | Low |
| 16 October 2020 | 29 June 2021 | 61-G | Maine | Intermediate |
| NA | 17 June 2021 | 61-I | Maine | Low |
| 1 October 2020 | 14 June 2021 | 61-K | Maine | High |
| 1 October 2020 | 14 June 2021 | 61-N | Maine | High |
| 1 October 2020 | 14 June 2021 | 61-P | Maine | High |
| 1 October 2020 | 14 June 2021 | 61-Q | Maine | High |
| NA | 28 June 2021 | 61-U | Maine | Intermediate |
| 16 October 2020 | 15 June 2021 | 94-A | Michigan | Intermediate |
| 16 October 2020 | 28 June 2021 | 94-B | Michigan | Intermediate |
| 1 October 2020 | 15 June 2021 | 94-E | Michigan | Intermediate |
| 16 October 2020 | 15 June 2021 | 94-F | Michigan | Low |
| 16 October 2020 | 14 June 2021 | 94-H | Michigan | Low |
| 1 October 2020 | 29 June 2021 | 94-I | Michigan | Intermediate |
| 1 October 2020 | 15 June 2021 | 94-J | Michigan | High |
| 1 October 2020 | 14 June 2021 | 94-K | Michigan | High |
| 1 October 2020 | 15 June 2021 | 94-M | Michigan | Intermediate |
| 1 October 2020 | 29 June 2021 | 94-N | Michigan | Intermediate |
| Fall 2020 Collection (N = 26) | |||
| Tree Condition | High Symptom (n = 7) | Intermediate Symptom (n = 13) | Low Symptom (n = 6) |
| ECM tips/g | 188.6 | 388.7 | 549.8 |
| Spring 2021 Collection (N = 30) | |||
| Tree Condition | High Symptom (n = 7) | Intermediate Symptom (n = 14) | Low Symptom (n = 9) |
| ECM tips/g | 254.2 | 271.8 | 267.8 |
| Combined Fall 2020 and Spring 2021 Collections (N = 56) | |||
| Tree Condition | High Symptom (n = 14) | Intermediate Symptom (n = 27) | Low Symptom (n = 15) |
| ECM tips/g | 221.4 | 328.1 | 380.6 |
| Coefficient | Value | Std. Error | p Value |
|---|---|---|---|
| Fall 2020 Collection | |||
| Low Symptom | 1.10 | 0.48 | p = 0.03 * |
| Intermediate Symptom | 0.74 | 0.40 | p = 0.07 |
| Spring 2021 Collection | |||
| Low Symptom | 0.16 | 0.43 | p = 0.71 |
| Intermediate Symptom | 0.11 | 0.39 | p = 0.78 |
| Combined Fall 2020 and Spring 2021 Collection | |||
| Low Symptom | 0.56 | 0.32 | p = 0.08 |
| Intermediate Symptom | 0.42 | 0.28 | p = 0.15 |
| Fall 2020 Collection (N = 26) | ||
| Provenance | Maine (n = 5) | Michigan (n = 21) |
| ECM tips/g | 244.32 | 402.4 |
| Spring 2021 Collection (N = 30) | ||
| Provenance | Maine (n = 9) | Michigan (n = 21) |
| ECM tips/g | 114.7 | 331.5 |
| Combined Fall 2020 and Spring 2021 Collection (N = 56) | ||
| Provenance | Maine (n = 14) | Michigan (n = 42) |
| ECM tips/g | 161.01 | 366.96 |
| Coefficient | Value | Std. Error | p Value |
|---|---|---|---|
| Fall 2020 Collection | |||
| Michigan | 0.36 | 0.46 | p = 0.44 |
| Spring 2021 Collection | |||
| Michigan | 1.02 | 0.27 | p < 0.001 * |
| Combined Fall 2020 and Spring 2021 Collection | |||
| Michigan | 0.78 | 0.25 | p = 0.003 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bashian-Victoroff, C.; Brown, A.; Loyd, A.L.; Carrino-Kyker, S.R.; Burke, D.J. Beech Leaf Disease Severity Affects Ectomycorrhizal Colonization and Fungal Taxa Composition. J. Fungi 2023, 9, 497. https://doi.org/10.3390/jof9040497
Bashian-Victoroff C, Brown A, Loyd AL, Carrino-Kyker SR, Burke DJ. Beech Leaf Disease Severity Affects Ectomycorrhizal Colonization and Fungal Taxa Composition. Journal of Fungi. 2023; 9(4):497. https://doi.org/10.3390/jof9040497
Chicago/Turabian StyleBashian-Victoroff, Claudia, Alexis Brown, Andrew L. Loyd, Sarah R. Carrino-Kyker, and David J. Burke. 2023. "Beech Leaf Disease Severity Affects Ectomycorrhizal Colonization and Fungal Taxa Composition" Journal of Fungi 9, no. 4: 497. https://doi.org/10.3390/jof9040497
APA StyleBashian-Victoroff, C., Brown, A., Loyd, A. L., Carrino-Kyker, S. R., & Burke, D. J. (2023). Beech Leaf Disease Severity Affects Ectomycorrhizal Colonization and Fungal Taxa Composition. Journal of Fungi, 9(4), 497. https://doi.org/10.3390/jof9040497