

Additions to Bambusicolous Fungi of Savoryellaceae from Southwest China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Morphological Examination
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Phylogenetic Analyses
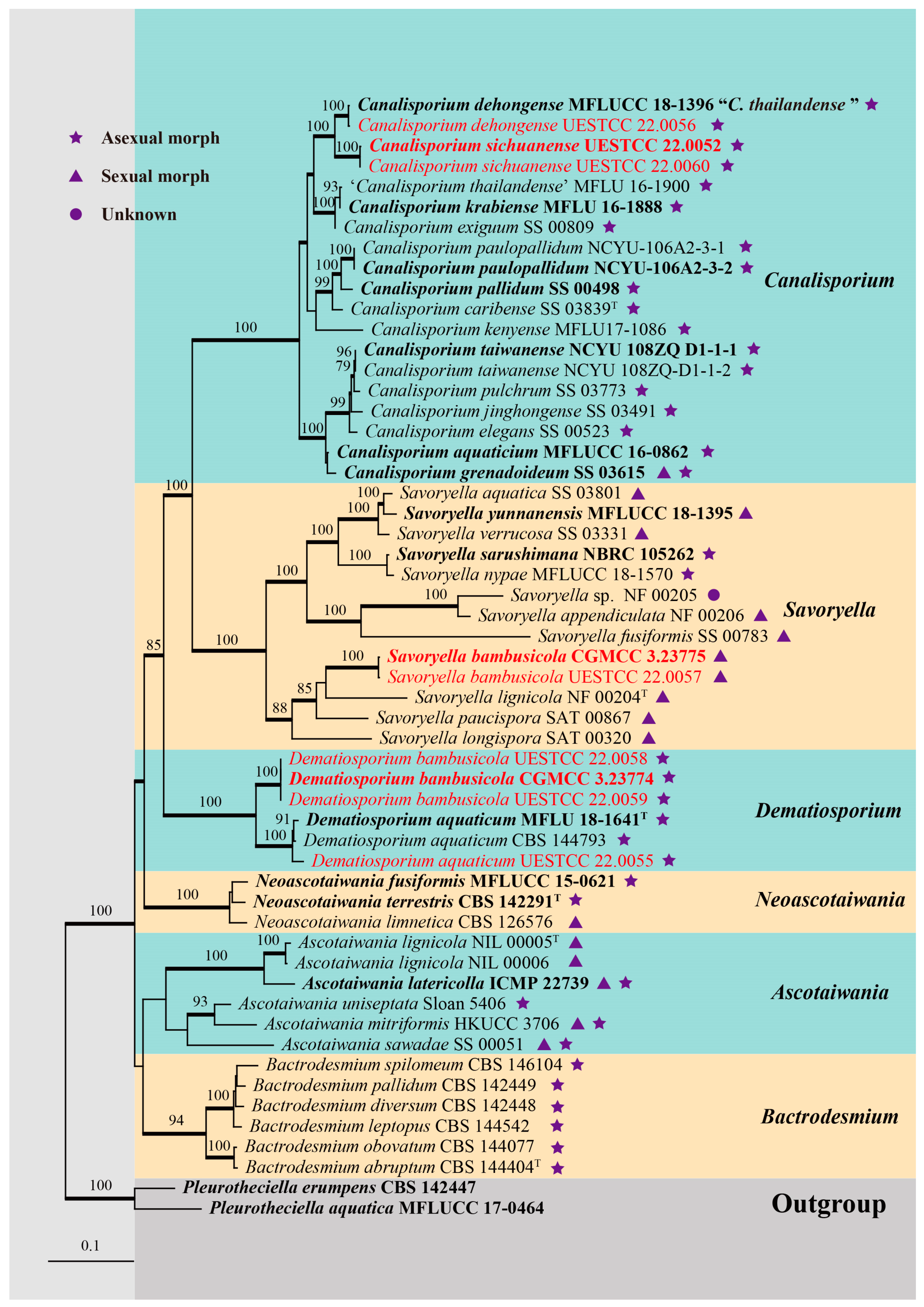
3. Results
3.1. Phylogenetic Analyses
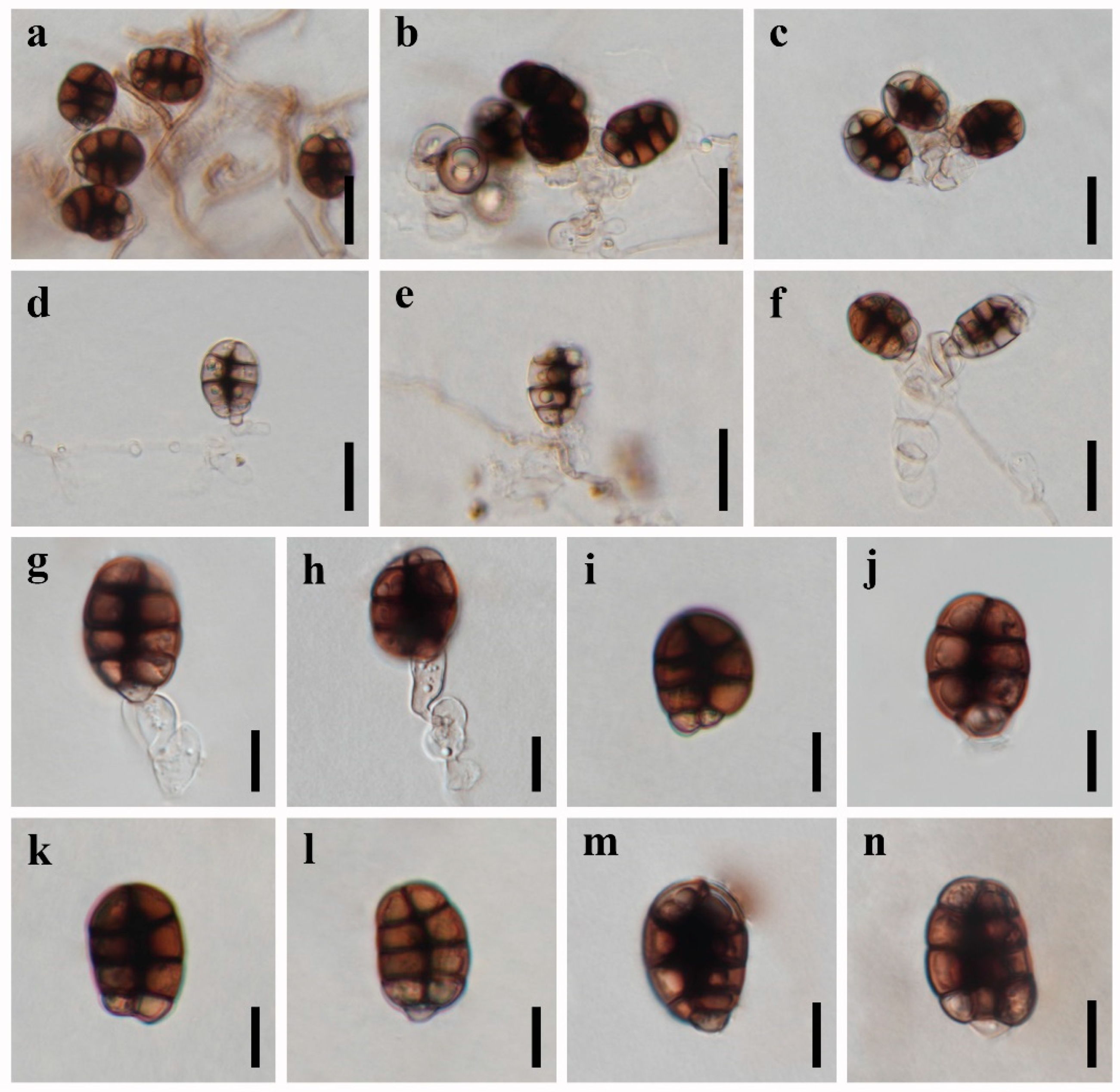
3.2. Taxonomy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Phookamsak, R.; Jiang, H.; Suwannarach, N.; Lumyong, S.; Xu, J.; Xu, S.; Liao, C.F.; Chomnunti, P. Bambusicolous Fungi in Pleosporales: Introducing Four Novel Taxa and a New Habitat Record for Anastomitrabeculia didymospora. J. Fungi 2022, 8, 630. [Google Scholar] [CrossRef] [PubMed]
- Dai, D.Q.; Phookamsak, R.; Wijayawardene, N.N.; Li, W.J.; Bhat, D.J.; Xu, J.C.; Taylor, J.E.; Hyde, K.D.; Chukeatirote, E. Bambusicolous fungi. Fungal Divers. 2017, 82, 1–105. [Google Scholar] [CrossRef]
- Shukla, A.; Singh, A.; Tiwari, D.; Ahirwar, B.K. Bambusicolous fungi: A reviewed documentation. Int. J. Pure Appl. Biosci. 2016, 4, 304–310. [Google Scholar] [CrossRef]
- Hyde, K.D.; Zhou, D.Q.; Dalisay, T. Bambusicolous fungi: A review. Fungal Divers. 2002, 9, 1–14. [Google Scholar]
- Tanaka, K.; Hirayama, K.; Yonezawa, H.; Hatakeyama, S.; Harada, Y.; Sano, T.; Shirouzu, T.; Hosoya, T. Molecular taxonomy of bambusicolous fungi: Tetraplosphaeriaceae, a new pleosporalean family with Tetraploa-like anamorphs. Stud. Mycol. 2009, 64, 175–209. [Google Scholar] [CrossRef]
- Dai, D.Q.; Tang, L.Z.; Wang, H.B. A Review of Bambusicolous Ascomycetes. Bamboo Curr. Future Prospect. 2018, 165, 10. [Google Scholar] [CrossRef]
- Shi, J.Y.; Zhou, D.Q.; Ma, L.S.; Yao, J.; Zhang, D.M. Diversity of Bamboo Species in China. World Bamboo Ratt. 2020, 18, 55–65. [Google Scholar] [CrossRef]
- Jiang, H.; Phookamsak, R.; Hongsanan, S.; Bhat, D.J.; Mortimer, P.E.; Suwannarach, N.; Kakumyan, P.; Xu, J. A review of bambusicolous Ascomycota in China with an emphasis on species richness in southwest China. Stud. Fungi 2022, 7, 1–33. [Google Scholar] [CrossRef]
- Yu, X.D.; Zhang, S.N.; Liu, J.K. Morpho-Phylogenetic Evidence Reveals Novel Pleosporalean Taxa from Sichuan Province, China. J. Fungi 2022, 8, 720. [Google Scholar] [CrossRef]
- Tai, F.L. Sylloge Fungorum Sinicorum. Sylloge Fungorum Sinicorum; Science Press: Beijing, China, 1979. [Google Scholar]
- Yang, C.L.; Xu, X.L.; Liu, Y.G. Two new species of Bambusicola (Bambusicolaceae, Pleosporales) on Phyllostachys heteroclada from Sichuan, China. Nova Hedwig. 2019, 108, 527–545. [Google Scholar] [CrossRef]
- Zeng, Q.; Lv, Y.C.; Xu, X.L.; Deng, Y.; Wang, F.H.; Liu, S.Y.; Liu, L.J.; Yang, C.L.; Liu, Y.G. Morpho-Molecular Characterization of Microfungi Associated with Phyllostachys (Poaceae) in Sichuan, China. J. Fungi 2022, 8, 702. [Google Scholar] [CrossRef]
- Yang, C.L.; Xu, X.L.; Dong, W.; Wanasinghe, D.N.; Liu, Y.G.; Hyde, K.D. Introducing Arthrinium phyllostachium sp. nov.(Apiosporaceae, Xylariales) on Phyllostachys heteroclada from Sichuan province, China. Phytotaxa 2019, 406, 91–110. [Google Scholar] [CrossRef]
- Li, L.; Guan, M.; Liu, H.; Zhu, T.; Li, S.; Han, S.; Lin, T.; Qiao, T.; Liu, Y.; Yang, C. Fusarium proliferatum Associated with Basal Rot Disease of Bambusa pervariabilis × Dendrocalamopsis grandis in China. Plant Dis. 2022, 6, 2531. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Xu, X.; Liu, Y.; Hyde, K.D.; Mckenzie, E.H. A new species of Phyllachora (Phyllachoraceae, Phyllachorales) on Phyllostachys heteroclada from Sichuan, China. Phytotaxa 2019, 392, 186–196. [Google Scholar] [CrossRef]
- Dayarathne, M.C.; Maharachchikumbura, S.S.N.; Jones, E.B.G.; Dong, W.; Devadatha, B.; Yang, J.; Ekanayaka, A.H.; De Silva, W.; Sarma, V.V.; Al-Sadi, A.M.; et al. Phylogenetic Revision of Savoryellaceae and Evidence for Its Ranking as a Subclass. Front. Microbiol. 2019, 10, 840. [Google Scholar] [CrossRef] [PubMed]
- Réblová, M.; Hernández-Restrepo, M.; Fournier, J.; Nekvindová, J. New insights into the systematics of Bactrodesmium and its allies and introducing new genera, species and morphological patterns in the Pleurotheciales and Savoryellales (Sordariomycetes). Stud. Mycol. 2020, 95, 415–466. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.G.; Eaton, R. Savoryella lignicola gen. et sp. nov. from water-cooling towers. Trans. Br. Mycol. Soc. 1969, 52, 161–165. [Google Scholar] [CrossRef]
- Zhang, S.N.; Abdel-Wahab, M.A.; Jones, E.B.G.; Hyde, K.D.; Liu, J.K. Additions to the genus Savoryella (Savoryellaceae), with the asexual morphs Savoryella nypae comb. nov. and S. sarushimana sp. nov. Phytotaxa 2019, 408, 195–207. [Google Scholar] [CrossRef]
- Tibpromma, S.; Hyde, K.D.; McKenzie, E.H.C.; Bhat, D.J.; Phillips, A.J.L.; Wanasinghe, D.N.; Samarakoon, M.C.; Jayawardena, R.S.; Dissanayake, A.J.; Tennakoon, D.S.; et al. Fungal diversity notes 840–928: Micro-fungi associated with Pandanaceae. Fungal Divers. 2018, 93, 1–160. [Google Scholar] [CrossRef]
- Hyde, K.D. Tropical Australian freshwater fungi. V.* Bombardia sp., Jahnula australiensis sp. nov., Savoryella aquatica sp. nov. and S. lignicola sp. nov. Aust. Syst. Bot. 1993, 6, 161–167. [Google Scholar] [CrossRef]
- Manimohan, P.; Amritha, M.; Sairabanu, N.K. A comparison of diversity of marine fungi on three co-habiting mangrove plants. Mycosphere 2011, 2, 533–538. [Google Scholar] [CrossRef]
- Yang, L.; Bao, D.F.; Luo, Z.L.; Su, X.J.; Su, H.Y. Neoascotaiwania aquatica sp. nov. from a freshwater habitat in Yunnan Province, China. Phytotaxa 2022, 531, 120–128. [Google Scholar] [CrossRef]
- Jaklitsch, W. Savoryellaceae Jaklitsch & Réblová. Index Fungorum 2015, 209, 1. [Google Scholar]
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D.; Chen, Y.J.; Bao, D.F.; Boonmee, S.; Calabon, M.S.; et al. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Luo, Z.L.; Hyde, K.D.; Liu, J.K.; Maharachchikumbura, S.S.N.; Jeewon, R.; Bao, D.F.; Bhat, D.J.; Lin, C.G.; Li, W.L.; Yang, J.; et al. Freshwater Sordariomycetes. Fungal Divers. 2019, 99, 451–660. [Google Scholar] [CrossRef]
- Nawawi, A.; Kuthubutheen, A.J. Canalisporium, a new genus of lignicolous hyphomycetes from Malaysia. Mycotaxon 1989, 34, 475–487. [Google Scholar]
- Sri-Indrasutdhi, V.; Boonyuen, N.; Suetrong, S.; Chuaseeharonnachai, C.; Sivichai, S.; Jones, E.B.G. Wood-inhabiting freshwater fungi from Thailand: Ascothailandia grenadoidia gen. et sp nov., Canalisporium grenadoidia sp nov with a key to Canalisporium species (Sordariomycetes, Ascomycota). Mycoscience 2010, 51, 411–420. [Google Scholar] [CrossRef]
- Ho, W.H.; Hyde, K.D.; Hodgkiss, I.J. Ascomycetes from tropical freshwater habitats: The genus Savoryella, with two new species. Mycol. Res. 1997, 101, 803–809. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.C.; Bhat, J.D.; Dayarathne, M.C.; Huang, S.K.; Norphanphoun, C.; Senanayake, I.C.; Perera, R.H.; et al. Families of Sordariomycetes. Fungal Divers. 2016, 79, 1–317. [Google Scholar] [CrossRef]
- Senanayake, I.; Rathnayake, A.; Marasinghe, D.; Calabon, M.; Gentekaki, E.; Lee, H. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Liu, J.K.; Chomnunti, P.; Cai, L.; Phookamsak, R.; Chukeatirote, E.; Jones, E.B.G.; Moslem, M.; Hyde, K.D. Phylogeny and morphology of Neodeightonia palmicola sp. nov. from palms. Sydowia 2010, 62, 261–276. [Google Scholar]
- Crous, P.W.; Gams, W.; Stalpers, J.A.; Robert, V.; Stegehuis, G. MycoBank: An online initiative to launch mycology into the 21st century. Stud. Mycol. 2004, 50, 19–22. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols. A Guide to Methods and Applications; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Kazutaka, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In Proceedings of the Nucleic Acids Symposium Series; Oxford University Press: Oxford, UK, 1999; pp. 95–98. [Google Scholar]
- Dissanayake, A.J.; Bhunjun, C.S.; Maharachchikumbura, S.S.M.; Liu, J.K. Applied aspects of methods to infer phylogenetic relationships amongst fungi. Mycosphere 2020, 11, 2652–2676. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods); Sinauer Associates: Sunderland, MA, USA, 2003. [Google Scholar]
- Rannala, B.; Yang, Z. Probability distribution of molecular evolutionary trees: A new method of phylogenetic inference. J. Mol. Evol. 1996, 43, 304–311. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef]
- Nylander, J.; Zoology, S.; Posada, D.; Mrmodeltest, R.; Os, F. MrModeltest2 v. 2.3 (Program for Selecting DNA Substitution Models Using PAUP*); Evolutionary Biology Centre: Uppsala, Sweden, 2008. [Google Scholar]
- Page, R.D.M. Tree View: An application to display phylogenetic trees on personal computers. Comput. Appl. Biosci. 1996, 12, 357–358. [Google Scholar] [CrossRef]
- Hyde, K.D.; Tennakoon, D.S.; Jeewon, R.; Bhat, D.J.; Maharachchikumbura, S.S.N.; Rossi, W.; Leonardi, M.; Lee, H.B.; Mun, H.Y.; Houbraken, J.; et al. Fungal diversity notes 1036–1150: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2019, 96, 1–242. [Google Scholar] [CrossRef]
- Koukol, O.; Delgado, G. Why morphology matters: The negative consequences of hasty descriptions of putative novelties in asexual ascomycetes. Ima Fungus 2021, 12, 1–8. [Google Scholar] [CrossRef]
- Goh, T.K.; Kuo, C.H. Reflections on Canalisporium, with descriptions of new species and records from Taiwan. Mycol. Prog. 2021, 20, 647–680. [Google Scholar] [CrossRef]
- Phookamsak, R.; Hyde, K.D.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Raspé, O.; Karunarathna, S.C.; Wanasinghe, D.N.; Hongsanan, S.; et al. Fungal diversity notes 929–1035: Taxonomic and phylogenetic contributions on genera and species of fungi. Fungal Divers. 2019, 95, 1–273. [Google Scholar] [CrossRef]
- Dulymamode, R.; Cannon, P.F.; Hyde, K.D.; Peerally, A. Four new ascomycete species from endemic Pandanus of Mauritius. Fungal Divers. 2001, 8, 87–96. [Google Scholar]
- Hyde, K. Fungi on palms XXV. A new species of Ascotaiwania. Sydowia 1995, 47, 31–37. [Google Scholar]
- Chang, H.S.; Hsieh, S.Y.; Jones, E.B.G.; Read, S.J.; Moss, S.T. New freshwater species of Ascotaiwania and Savoryella from Taiwan. Mycol. Res. 1998, 102, 709–718. [Google Scholar] [CrossRef]
- Ranghoo, V.M.; Hyde, K.D. Ascomycetes from freshwater habitats: Ascolacicola aquatica gen. et sp. nov. and a new species of Ascotaiwania from wood submerged in a reservoir in Hong Kong. Mycologia 1998, 90, 1055–1062. [Google Scholar] [CrossRef]
- Hyde, K.D.; Goh, T.K. Fungi on submerged wood from the River Coln, England. Mycol. Res. 1999, 103, 1561–1574. [Google Scholar] [CrossRef]
- Borse, B.; Pawar, N.; Patil, S. Bactrodesmium aquatica sp. nov.(mitosporic fungi) on submerged wood from freshwater, Maharashtra, India. Int. J. Adv. Res. 2019, 7, 1365–1368. [Google Scholar] [CrossRef]
- Hyde, K.D.; Dong, Y.; Phookamsak, R.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Liu, N.G.; Abeywickrama, P.D.; Mapook, A.; Wei, D.; et al. Fungal diversity notes 1151–1276: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2020, 100, 5–277. [Google Scholar] [CrossRef]
- Goh, T.K.; Ho, W.H.; Hyde, K.D.; Whitton, S.R.; Umali, T.E. New records and species of Canalisporium (Hyphomycetes), with a revision of the genus. Can. J. Bot. 1998, 76, 142–152. [Google Scholar] [CrossRef]
- Cai, L.; Kq, Z.; McKenzie, E.; Lumyong, S.; Hyde, K. New species of Canalisporium and Dictyosporium from China and a note on the differences between these genera. Cryptogam. Mycol. 2003, 24, 3–11. [Google Scholar]
- Ferrer, A. New records and a new species of Canalisporium from aquatic habitats in Panama. Mycotaxon 2005, 93, 179–188. [Google Scholar]
- Goh, T.K.; Hyde, K.D. A new species of Canalisporium from Australia. Mycologia 2000, 92, 589–592. [Google Scholar] [CrossRef]
- Yang, J.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Hyde, K.D.; Mckenzie, E.H.C.; Jones, E.B.G.; Alsadi, A.M.; Lumyong, S. Fuscosporellales, a New Order of Aquatic and Terrestrial Hypocreomycetidae (Sordariomycetes). Cryptogam. Mycologie 2016, 4, 449–475. [Google Scholar] [CrossRef]
- Du, H.Z.; Yang, J.; Liu, N.G.; Cheewangkoon, R.; Liu, J.K. Morpho-Phylogenetic Evidence Reveals New Species of Fuscosporellaceae and Savoryellaceae from Freshwater Habitats in Guizhou Province, China. J. Fungi 2022, 8, 1138. [Google Scholar] [CrossRef] [PubMed]
- Tsui, C.K.M.; Hyde, K.D. Biodiversity of fungi on submerged wood in a stream and its estuary in the Tai Ho Bay, Hong Kong. Fungal Divers. 2004, 15, 205–220. [Google Scholar]
- Hernández-Restrepo, M.; Gené, J.; Castañeda-Ruiz, R.F.; Mena-Portales, J.; Crous, P.W.; Guarro, J. Phylogeny of saprobic microfungi from Southern Europe. Stud. Mycol. 2017, 86, 53–97. [Google Scholar] [CrossRef]
- Chang, H. Trichocladium anamorph of Ascotaiwania hsilio and monodictys-like anamorphic states of Ascotaiwania lignicola. Fungal. Sci. 2001, 16, 35–38. [Google Scholar]
- Sivichai, S.; Hywel-Jones, N.; Jones, E. Lignicolous freshwater ascomycota from Thailand: 1. Ascotaiwania sawada and its anamorph state Monotosporella. Mycoscience 1998, 39, 307–311. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Vouchers/Strains/Isolates | GenBank Accession Numbers | ||||
|---|---|---|---|---|---|---|
| SSU | ITS | LSU | rpb2 | tef1α | ||
| Ascotaiwania latericolla | ICMP 22739 | N/A | MN699390 | MN699407 | MN704312 | N/A |
| Ascotaiwania lignicola | NIL 00005 | HQ446284 | HQ446341 | HQ446364 | HQ446419 | HQ446307 |
| Ascotaiwania lignicola | NIL 00006 | HQ446285 | HQ446342 | HQ446365 | N/A | HQ446308 |
| Ascotaiwania mitriformis | HKUCC 3706 | N/A | N/A | AF132324 | N/A | N/A |
| Ascotaiwania sawadae | SS 00051 | HQ446283 | HQ446340 | HQ446363 | HQ446418 | HQ446306 |
| Ascotaiwania uniseptata | Sloan 5406 | N/A | N/A | KT278718 | N/A | N/A |
| Bactrodesmium abruptum | CBS 144404 | MN699365 | MN699391 | MN699408 | MN704288 | MN704313 |
| Bactrodesmium diversum | CBS 142448 | MN699369 | MN699352 | MN699412 | MN704292 | MN704317 |
| Bactrodesmium leptopus | CBS 144542 | MN699374 | MN699388 | MN699423 | MN704297 | MN704321 |
| Bactrodesmium obovatum | CBS 144077 | MN699375 | MN699395 | MN699424 | MN704298 | MN704322 |
| Bactrodesmium pallidum | CBS 142449 | MN699379 | MN699363 | MN699428 | MN704301 | MN704326 |
| Bactrodesmium spilomeum | CBS 146104 | MN699381 | N/A | N/A | MN704303 | MN704328 |
| Canalisporium aquaticium | MFLUCC 16-0862 | MN061353 | MN061351 | MN061365 | N/A | N/A |
| Canalisporium caribense | SS 03839 | GQ390253 | GQ390283 | GQ390268 | HQ446421 | N/A |
| Canalisporium dehongense * | UESTCC 22.0056 | OQ428250 | OQ428266 | OQ428258 | OQ437183 | OQ437176 |
| Canalisporium dehongense | MFLUCC 18-1396 | MK051035 | MK051033 | MK051034 | N/A | N/A |
| Canalisporium elegans | SS 00523 | GQ390255 | GQ390285 | GQ390270 | HQ446423 | HQ446310 |
| Canalisporium exiguum | SS 00809 | GQ390266 | GQ390296 | GQ390281 | HQ446436 | N/A |
| Canalisporium grenadoideum | SS 03615 | GQ390252 | GQ390282 | GQ390267 | HQ446420 | HQ446309 |
| Canalisporium jinghongense | SS 03491 | GQ390257 | GQ390287 | GQ390272 | HQ446426 | HQ446313 |
| Canalisporium kenyense | MFLU17-1086 | N/A | MH701998 | MH701999 | N/A | MH708885 |
| Canalisporium krabiense | MFLU 16-1888 | N/A | MH275051 | MH260283 | N/A | N/A |
| Canalisporium pallidum | SS 00498 | GQ390265 | GQ390295 | GQ390280 | HQ446435 | HQ446322 |
| Canalisporium paulopallidum | NCYU-106A2-3-1 | N/A | MT946658 | N/A | N/A | N/A |
| Canalisporium paulopallidum | NCYU-106A2-3-2 | N/A | MT946659 | N/A | N/A | N/A |
| Canalisporium pulchrum | SS 03773 | GQ390263 | GQ390293 | GQ390278 | HQ446432 | HQ446319 |
| Canalisporium sichuanense * | CGMCC 3.23926 | OQ428254 | OQ428270 | OQ428262 | OQ437186 | OQ437180 |
| Canalisporium sichuanense * | UESTCC 22.0060 | OQ428255 | OQ428271 | OQ428263 | N/A | N/A |
| Canalisporium taiwanense | NCYU-108ZQ-D1-1-1 | N/A | MT946663 | N/A | N/A | N/A |
| Canalisporium taiwanense | NCYU-108ZQ-D1-1-2 | N/A | MT946664 | N/A | N/A | N/A |
| ‘Canalisporium thailandense’ | MFLU 16-1900 | N/A | MH275052 | MH260284 | N/A | N/A |
| Dematiosporium aquaticum | CBS 144793 | MN699385 | MN699402 | MN699433 | MN704307 | MN704330 |
| Dematiosporium aquaticum | MFLU 18-1641 | N/A | N/A | MK835855 | MN194029 | MN200286 |
| Dematiosporium aquaticum * | UESTCC 22.0055 | N/A | N/A | OQ428257 | N/A | N/A |
| Dematiosporium bambusicola* | CGMCC 3.23774 | OQ428252 | OQ428268 | OQ428260 | N/A | OQ437178 |
| Dematiosporium bambusicola * | UESTCC 22.0058 | N/A | OQ428272 | OQ428264 | N/A | OQ437181 |
| Dematiosporium bambusicola * | UESTCC 22.0059 | OQ428256 | OQ428273 | OQ428265 | N/A | OQ437182 |
| Neoascotaiwania fusiformis | MFLUCC 15-0621 | N/A | MG388215 | KX550893 | KX576871 | N/A |
| Neoascotaiwania limnetica | CBS 126576 | KT278689 | KY853452 | KY853513 | MN704308 | MN704331 |
| Neoascotaiwania terrestris | CBS 142291 | KY853547 | KY853454 | KY853515 | N/A | N/A |
| Pleurotheciella aquatica | MFLUCC 17-0464 | MF399220 | MF399236 | MF399253 | MF401405 | N/A |
| Pleurotheciella erumpens | CBS 142447 | MN699387 | MN699406 | MN699435 | MN704311 | MN704334 |
| Savoryella appendiculata | NF 00206 | HQ446293 | HQ446350 | N/A | HQ446442 | HQ446327 |
| Savoryella aquatica | SS 03801 | HQ446292 | HQ446349 | HQ446372 | HQ446441 | HQ446326 |
| Savoryella bambusicola* | CGMCC 3.23775 | OQ428253 | OQ428269 | OQ428261 | OQ437185 | OQ437179 |
| Savoryella bambusicola * | UESTCC 22.0057 | OQ428251 | OQ428267 | OQ428259 | OQ437184 | OQ437177 |
| Savoryella fusiformis | SS 00783 | HQ446294 | HQ446351 | N/A | HQ446443 | HQ446328 |
| Savoryella lignicola | NF 00204 | HQ446300 | HQ446357 | HQ446378 | N/A | HQ446334 |
| Savoryella longispora | SAT 00320 | HQ446301 | HQ446358 | HQ446379 | HQ446449 | HQ446335 |
| Savoryella nypae | MFLUCC 18-1570 | MK543237 | MK543219 | MK543210 | N/A | MK542516 |
| Savoryella paucispora | SAT 00867 | HQ446304 | HQ446361 | HQ446382 | HQ446452 | HQ446338 |
| Savoryella sarushimana | NBRC 105262 | MK411005 | N/A | MK411004 | N/A | N/A |
| Savoryella sp. | NF 00205 | HQ446305 | HQ446362 | N/A | N/A | HQ446339 |
| Savoryella verrucosa | SS 03331 | HQ446298 | HQ446355 | HQ446376 | HQ446447 | HQ446332 |
| Savoryella yunnanensis | MFLUCC 18-1395 | MK411423 | N/A | MK411422 | N/A | MK411424 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, X.-D.; Zhang, S.-N.; Liu, J.-K. Additions to Bambusicolous Fungi of Savoryellaceae from Southwest China. J. Fungi 2023, 9, 571. https://doi.org/10.3390/jof9050571
Yu X-D, Zhang S-N, Liu J-K. Additions to Bambusicolous Fungi of Savoryellaceae from Southwest China. Journal of Fungi. 2023; 9(5):571. https://doi.org/10.3390/jof9050571
Chicago/Turabian StyleYu, Xian-Dong, Sheng-Nan Zhang, and Jian-Kui Liu. 2023. "Additions to Bambusicolous Fungi of Savoryellaceae from Southwest China" Journal of Fungi 9, no. 5: 571. https://doi.org/10.3390/jof9050571
APA StyleYu, X. -D., Zhang, S. -N., & Liu, J. -K. (2023). Additions to Bambusicolous Fungi of Savoryellaceae from Southwest China. Journal of Fungi, 9(5), 571. https://doi.org/10.3390/jof9050571