

Recent Advancements in Pathogenic Mechanisms, Applications and Strategies for Entomopathogenic Fungi in Mosquito Biocontrol


Abstract
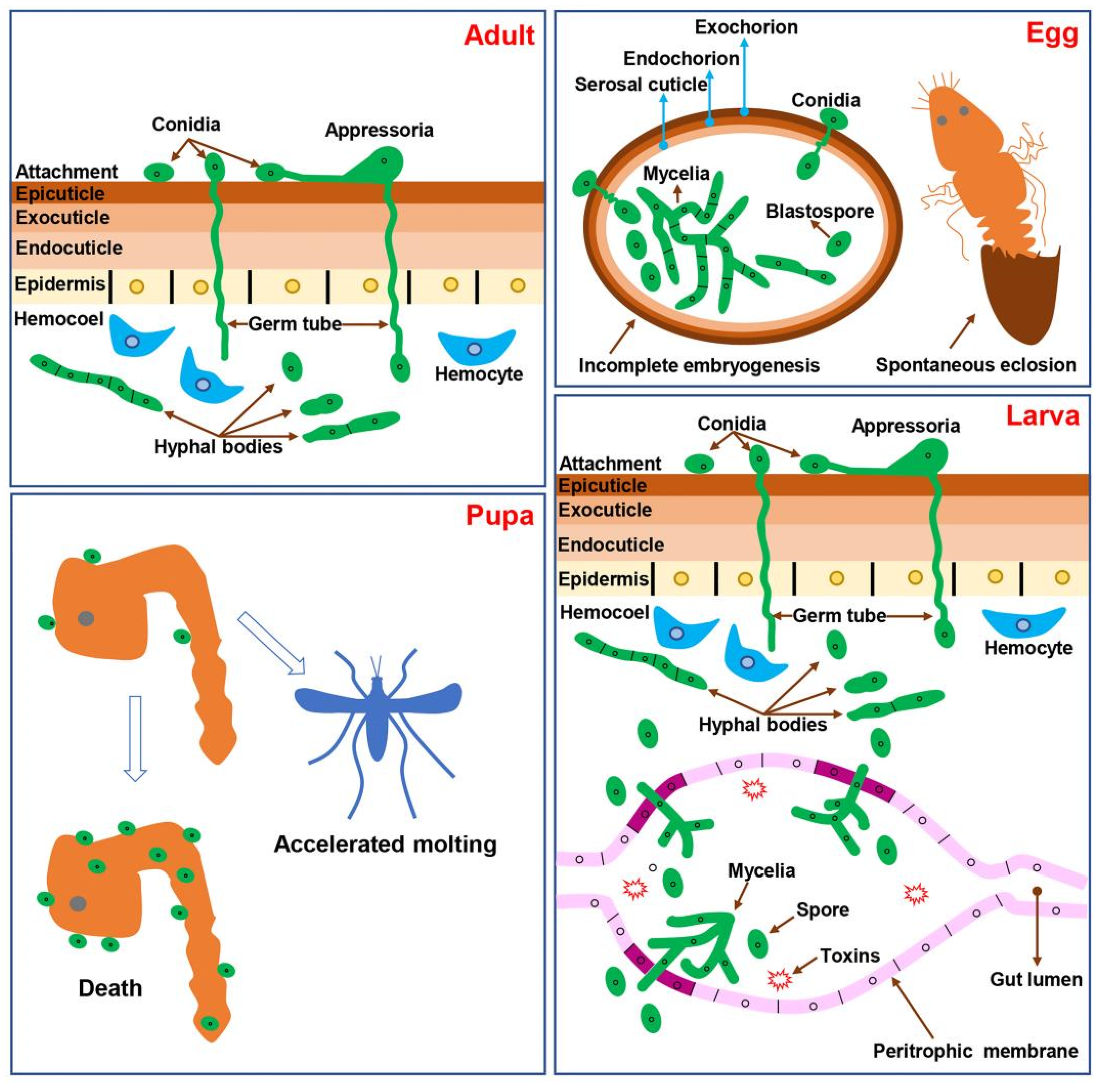
:1. Introduction
2. Pathogenic Mechanisms of EPF and Immune Response of Host Mosquito
2.1. Invasion through Mosquito Cuticle
2.2. Invasion through Digestive Tract
2.3. Immune Response of Mosquito
3. Effectiveness of EPF in Mosquito Control
3.1. Effectiveness of EPF on Different Development Stage of Mosquito
3.2. Factors That Influence Spore Quality
4. Combination of EPF with Other Strategies in Mosquito Control
4.1. Combined with Chemical Insecticides
4.2. Combined with Microbial Metabolites or Microbial Organisms
4.3. Combined with Mosquito Attractants
4.4. Combined with Predators
5. Engineering Manipulation of EPF to Improve Their Mosquitocidal Efficacy
5.1. Introducing Insecticidal Molecules into Mosquito
5.2. Introducing Antipathogen Effector to Block Vector Disease Transmission
5.3. Increasing the Fungal Tolerance to Adverse Environmental Conditions
6. Conclusions and Future Prospect
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Global Vector Control Response 2017–2030; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- World Health Organization. Vector-Borne Diseases. 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/vector-borne-diseases (accessed on 2 March 2020).
- World Health Organization. World Malaria Report 2016; World Health Organization: Geneva, Switzerland, 2016; ISBN 978-92-4-151171-1. [Google Scholar]
- Lovett, B.; Bilgo, E.; Diabate, A.; St. Leger, R. A review of progress toward field application of transgenic mosquitocidal entomopathogenic fungi. Pest. Manag. Sci. 2019, 75, 2316–2324. [Google Scholar] [CrossRef] [PubMed]
- Shen, D.; Nyawira, K.T.; Xia, A. New discoveries and applications of mosquito fungal pathogens. Curr. Opin. Insect. Sci. 2020, 40, 111–116. [Google Scholar] [CrossRef]
- Benelli, G. Managing mosquitoes and ticks in a rapidly changing world-facts and trends. Saudi J. Biol. Sci. 2019, 26, 921–929. [Google Scholar] [CrossRef]
- Gao, H.; Cui, C.; Wang, L.; Jacobs-Lorena, M.; Wang, S. Mosquito microbiota and implications for disease control. Trends Parasitol. 2020, 36, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Cafarchia, C.; Pellegrino, R.; Romano, V.; Friuli, M.; Demitri, C.; Pombi, M.; Benelli, G.; Otranto, D. Delivery and effectiveness of entomopathogenic fungi for mosquito and tick control: Current knowledge and research challenges. Acta Trop. 2022, 234, 106627. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, J.; Campos, V.C.; Humber, R.A.; Luz, C. Efficacy of Culicinomyces spp. against Aedes aegypti eggs, larvae and adults. J. Invertebr. Pathol. 2018, 157, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Shen, D.; Tang, Z.; Wang, C.; Wang, J.; Dong, Y.; Chen, Y.; Wei, Y.; Cheng, B.; Zhang, M.; Grenville-Briggs, L.J.; et al. Infection mechanisms and putative effector repertoire of the mosquito pathogenic oomycete Pythium guiyangense uncovered by genomic analysis. PLoS Genet. 2019, 15, e1008116. [Google Scholar] [CrossRef] [PubMed]
- De Paula, A.R.; Silva, L.E.I.; Ribeiro, A.; Da Silva, G.A.; Silva, C.P.; Butt, T.M.; Samuels, R.I. Metarhizium anisopliae blastospores are highly virulent to adult Aedes aegypti, an important arbovirus vector. Parasit Vectors 2021, 14, 555. [Google Scholar] [CrossRef]
- Renuka, S.; Vani, H.C.; Alex, E. Entomopathogenic fungi as a potential management tool for the control of urban malaria vector, Anopheles stephensi (Diptera: Culicidae). J. Fungi 2023, 9, 223. [Google Scholar] [CrossRef]
- Sousa, N.A.; Rodrigues, J.; Luz, C.; Humber, R.A. Exposure of newly deposited Aedes aegypti eggs to Metarhizium humberi and fungal development on the eggs. J. Invertebr. Pathol. 2023, 197, 107898. [Google Scholar] [CrossRef]
- Deng, S.; Huang, Q.; Wei, H.; Zhou, L.; Yao, L.; Li, D.; Wu, S.; Chen, J.; Peng, H. Beauveria bassiana infection reduces the vectorial capacity of Aedes albopictus for the Zika virus. J. Pest. Sci. 2019, 92, 781–789. [Google Scholar] [CrossRef]
- Shoukat, R.F.; Hassan, B.; Shakeel, M.; Zafar, J.; Li, S.; Freed, S.; Xu, X.; Jin, F. Pathogenicity and transgenerational effects of Metarhizium anisopliae on the demographic parameters of Aedes albopictus (Culicidae: Diptera). J. Med. Entomol. 2020, 57, 677–685. [Google Scholar] [CrossRef]
- Martinez, J.M.; Rodrigues, J.; Marreto, R.N.; Mascarin, G.M.; Fernandes, É.K.K.; Humber, R.A.; Luz, C. Efficacy of focal applications of a mycoinsecticide to control Aedes aegypti in Central Brazil. Appl. Microbiol. Biotechnol. 2021, 105, 8703–8714. [Google Scholar] [CrossRef] [PubMed]
- Lovett, B.; Bilgo, E.; Millogo, S.A.; Ouattarra, A.K.; Sare, I.; Gnambani, E.J.; Dabire, R.K.; Diabate, A.; St. Leger, R.J. Transgenic Metarhizium rapidly kills mosquitoes in a malaria-endemic region of Burkina Faso. Science 2019, 364, 894–897. [Google Scholar] [CrossRef]
- Abd El Rahman, S.; Hoffmann, B.; Karam, R.; El-Beskawy, M.; Hamed, M.F.; Forth, L.F.; Höper, D.; Eschbaumer, M. Sequence analysis of egyptian foot-and-mouth disease virus field and vaccine strains: Intertypic recombination and evidence for accidental release of virulent virus. Viruses 2020, 12, 990. [Google Scholar] [CrossRef]
- Niu, G.; Hao, Y.; Wang, X.; Gao, J.-M.; Li, J. Fungal metabolite asperaculane B inhibits malaria infection and transmission. Molecules 2020, 25, 3018. [Google Scholar] [CrossRef]
- Gutierrez, A.C.; Rueda Páramo, M.E.; Falvo, M.L.; López Lastra, C.C.; García, J.J. Leptolegnia Chapmanii (Straminipila: Peronosporomycetes) as a future biorational tool for the control of Aedes aegypti (L.). Acta Trop. 2017, 169, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Noskov, Y.A.; Polenogova, O.V.; Yaroslavtseva, O.N.; Belevich, O.E.; Yurchenko, Y.A.; Chertkova, E.A.; Kryukova, N.A.; Kryukov, V.Y.; Glupov, V.V. Combined effect of the entomopathogenic fungus Metarhizium robertsii and avermectins on the survival and immune response of Aedes aegypti larvae. PeerJ 2019, 7, e7931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wooding, M.; Naudé, Y.; Rohwer, E.; Bouwer, M. Controlling mosquitoes with semiochemicals: A review. Parasit Vectors 2020, 13, 80. [Google Scholar] [CrossRef]
- Malla, R.K.; Mandal, K.K.; Burman, S.; Das, S.; Ghosh, A.; Chandra, G. Numerical analysis of predatory potentiality of Toxorhynchites splendens against larval Aedes albopictus in laboratory and semi-field conditions. Sci. Rep. 2023, 13, 7403. [Google Scholar] [CrossRef]
- Buckner, E.A.; Williams, K.F.; Ramirez, S.; Darrisaw, C.; Carrillo, J.M.; Latham, M.D.; Lesser, C. A field efficacy evaluation of In2Care mosquito traps in comparison with routine integrated vector management at reducing Aedes aegypti. J. Am. Mosq. Control Assoc. 2021, 37, 242–249. [Google Scholar] [CrossRef]
- Wang, L.; Lai, Y.; Chen, J.; Cao, X.; Zheng, W.; Dong, L.; Zheng, Y.; Li, F.; Wei, G.; Wang, S. The ASH1-PEX16 regulatory pathway controls peroxisome biogenesis for appressorium-mediated insect infection by a fungal pathogen. Proc. Natl. Acad. Sci. USA 2023, 120, e2217145120. [Google Scholar] [CrossRef] [PubMed]
- Aw, K.M.S.; Hue, S.M. Mode of Infection of Metarhizium spp. fungus and their potential as biological control agents. J. Fungi 2017, 3, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Xia, Y. Host-pathogen interactions between Metarhizium spp. and locusts. J. Fungi 2022, 8, 602. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Chen, H.; Wei, G.; Wang, G.; Li, F.; Wang, S. In vivo gene expression profiling of the entomopathogenic fungus Beauveria bassiana elucidates its infection stratagems in Anopheles mosquito. Sci. China Life Sci. 2017, 60, 839–851. [Google Scholar] [CrossRef] [PubMed]
- De Sousa, N.A.; Rodrigues, J.; Arruda, W.; Humber, R.A.; Luz, C. Development of Metarhizium humberi in Aedes aegypti eggs. J. Invertebr. Pathol. 2021, 184, 107648. [Google Scholar] [CrossRef] [PubMed]
- Scholte, E.J.; Knols, B.G.J.; Takken, W. Infection of the malaria mosquito Anopheles gambiae with the entomopathogenic fungus Metarhizium anisopliae reduces blood feeding and fecundity. J. Invertebr. Pathol. 2006, 91, 43–49. [Google Scholar] [CrossRef]
- Scholte, E.-J.; Takken, W.; Knols, B.G.J. Infection of adult Aedes aegypti and Ae. Albopictus mosquitoes with the entomopathogenic fungus Metarhizium anisopliae. Acta Trop. 2007, 102, 151–158. [Google Scholar] [CrossRef]
- Ghosh, S.K.; Podder, D.; Mukherjee, A. An insight of Anopheline larvicidal mechanism of Trichoderma asperellum (TaspSKGN2). Sci. Rep. 2021, 11, 16029. [Google Scholar] [CrossRef]
- Carolino, A.T.; Gomes, S.A.; Pontes Teodoro, T.B.; Mattoso, T.C.; Samuels, R.I. Aedes aegypti pupae are highly susceptible to infection by Metarhizium anisopliae blastospores. J. Pure Appl. Microbiol. 2019, 13, 1629–1634. [Google Scholar] [CrossRef] [Green Version]
- Lai, Y.; Cao, X.; Chen, J.; Wang, L.; Wei, G.; Wang, S. Coordinated regulation of infection-related morphogenesis by the KMT2-Cre1-Hyd4 regulatory pathway to facilitate fungal infection. Sci. Adv. 2020, 6, eaaz1659. [Google Scholar] [CrossRef] [Green Version]
- Vivero-Gomez, R.J.; Castañeda-Monsalve, V.A.; Atencia, M.C.; Hoyos-Lopez, R.; Hurst, G.D.; Cadavid-Restrepo, G.; Moreno-Herrera, C.X. Molecular phylogeny of heritable symbionts and microbiota diversity analysis in phlebotominae sand flies and Culex nigripalpus from Colombia. PLoS Negl. Trop. Dis. 2021, 15, e0009942. [Google Scholar] [CrossRef] [PubMed]
- Angleró-Rodríguez, Y.I.; Blumberg, B.J.; Dong, Y.; Sandiford, S.L.; Pike, A.; Clayton, A.M.; Dimopoulos, G. A natural Anopheles-associated Penicillium chrysogenum enhances mosquito susceptibility to Plasmodium infection. Sci. Rep. 2016, 6, 34084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, G.; Lai, Y.; Wang, G.; Chen, H.; Li, F.; Wang, S. Insect pathogenic fungus interacts with the gut microbiota to accelerate mosquito mortality. Proc. Natl. Acad. Sci. USA 2017, 114, 5994–5999. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.; Wang, Y.; Liu, J.; Zhao, J.; Sun, P.; Wang, S. A fungal pathogen deploys a small silencing RNA that attenuates mosquito immunity and facilitates infection. Nat. Commun. 2019, 10, 4298. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Luo, S.; Clarke, B.B.; Belanger, F.C. The Epichloë festucae antifungal protein Efe-AfpA is also a possible effector protein required for the interaction of the fungus with its host grass Festuca rubra subsp. rubra. Microorganisms 2021, 9, 140. [Google Scholar] [CrossRef] [PubMed]
- Greenfield, B.P.J.; Lord, A.M.; Dudley, E.; Butt, T.M. Conidia of the Insect pathogenic fungus, Metarhizium anisopliae, fail to adhere to mosquito larval cuticle. R. Soc. Open Sci. 2014, 1, 140193. [Google Scholar] [CrossRef] [Green Version]
- Bawin, T.; Seye, F.; Boukraa, S.; Zimmer, J.Y.; Raharimalala, F.N.; Ndiaye, M.; Compere, P.; Delvigne, F.; Francis, F. Histopathological effects of Aspergillus clavatus (Ascomycota: Trichocomaceae) on larvae of the southern house mosquito, Culex quinquefasciatus (Diptera: Culicidae). Fungal Biol. 2016, 120, 489–499. [Google Scholar] [CrossRef]
- Alkhaibari, A.M.; Carolino, A.T.; Yavasoglu, S.I.; Maffeis, T.; Mattoso, T.C.; Bull, J.C.; Samuels, R.I.; Butt, T.M. Metarhizium brunneum blastospore pathogenesis in Aedes aegypti larvae: Attack on several fronts accelerates mortality. PLoS Pathog. 2016, 12, e1005715. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira Barbosa Bitencourt, R.; Reis Dos Santos Mallet, J.; Mesquita, E.; Silva Gôlo, P.; Fiorotti, J.; Rita Elias Pinheiro Bittencourt, V.; Guedes Pontes, E.; Da Costa Angelo, I. Larvicidal activity, route of interaction and ultrastructural changes in Aedes aegypti exposed to entomopathogenic fungi. Acta Trop. 2021, 213, 105732. [Google Scholar] [CrossRef]
- De Oliveira Barbosa Bitencourt, R.; Corrêa, T.A.; Santos-Mallet, J.; Santos, H.A.; Lowenberger, C.; Moreira, H.V.S.; Gôlo, P.S.; Bittencourt, V.R.E.P.; Da Costa Angelo, I. Beauveria bassiana interacts with gut and hemocytes to manipulate Aedes aegypti immunity. Parasit Vectors 2023, 16, 17. [Google Scholar] [CrossRef] [PubMed]
- Bitencourt, R.D.O.B.; Santos-Mallet, J.R.D.; Lowenberger, C.; Ventura, A.; Gôlo, P.S.; Bittencourt, V.R.E.P.; Angelo, I.D.C. A novel model of pathogenesis of Metarhizium anisopliae propagules through the midguts of Aedes aegypti larvae. Insects 2023, 14, 328. [Google Scholar] [CrossRef] [PubMed]
- Vivekanandhan, P.; Kavitha, T.; Karthi, S.; Senthil-Nathan, S.; Shivakumar, M. Toxicity of Beauveria bassiana-28 mycelial extracts on larvae of Culex quinquefasciatus mosquito (Diptera: Culicidae). Int. J. Environ. Res. Public Health 2018, 15, 440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vivekanandhan, P.; Bedini, S.; Shivakumar, M.S. Isolation and identification of entomopathogenic fungus from Eastern Ghats of South Indian forest soil and their efficacy as biopesticide for mosquito control. Parasitol. Int. 2020, 76, 102099. [Google Scholar] [CrossRef]
- Vivekanandhan, P.; Swathy, K.; Murugan, A.C.; Krutmuang, P. Insecticidal efficacy of Metarhizium anisopliae derived chemical constituents against disease-vector mosquitoes. J. Fungi 2022, 8, 300. [Google Scholar] [CrossRef]
- Ragavendran, C.; Natarajan, D. Insecticidal potency of Aspergillus terreus against larvae and pupae of three mosquito species Anopheles stephensi, Culex quinquefasciatus, and Aedes aegypti. Environ. Sci. Pollut. Res. Int. 2015, 22, 17224–17237. [Google Scholar] [CrossRef]
- Kumar, A.; Srivastava, P.; Sirisena, P.; Dubey, S.K.; Kumar, R.; Shrinet, J.; Sunil, S. Mosquito Innate Immunity. Insects 2018, 9, 95. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.H.; Chang, M.M.; Wang, X.L.; Zheng, A.H.; Zou, Z. The immune strategies of mosquito Aedes aegypti against microbial infection. Dev. Comp. Immunol. 2018, 83, 12–21. [Google Scholar] [CrossRef]
- Ramirez, J.L.; Muturi, E.J.; Barletta, A.B.F.; Rooney, A.P. The Aedes aegypti IMD pathway is a critical component of the mosquito antifungal immune response. Dev. Comp. Immunol. 2019, 95, 1–9. [Google Scholar] [CrossRef]
- Wang, Y.; Chang, M.; Wang, M.; Ji, Y.; Sun, X.; Raikhel, A.S.; Zou, Z. OTU7B modulates the mosquito immune response to Beauveria bassiana Infection via deubiquitination of the Toll adaptor TRAF4. Microbiol. Spectr. 2023, 11, e03123-22. [Google Scholar] [CrossRef]
- Wang, Y.; Cui, C.; Wang, G.; Li, Y.; Wang, S. Insects defend against fungal infection by employing microRNAs to silence virulence-related genes. Proc. Natl. Acad. Sci. USA 2021, 118, e2023802118. [Google Scholar] [CrossRef]
- Hamama, H.M.; Zyaan, O.H.; Abu Ali, O.A.; Saleh, D.I.; Elakkad, H.A.; El-Saadony, M.T.; Farag, S.M. Virulence of entomopathogenic fungi against Culex pipiens: Impact on biomolecules availability and life table parameters. Saudi J. Biol. Sci. 2022, 29, 385–393. [Google Scholar] [CrossRef]
- Falvo, M.L.; Musso, A.; Ordoqui, E.; López Lastra, C.C.; Luz, C.; García, J.J. Adulticidal activity of Metarhizium anisopliae s.l. (Hypocreales: Clavicipitaceae) native strains from argentina against Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2020, 57, 636–640. [Google Scholar] [CrossRef]
- Ondiaka, S.N.; Masinde, E.W.; Koenraadt, C.J.; Takken, W.; Mukabana, W.R. Effects of fungal infection on feeding and survival of Anopheles gambiae (Diptera: Culicidae) on plant sugars. Parasit Vectors 2015, 8, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tawidian, P.; Kang, Q.; Michel, K. The potential of a new Beauveria bassiana isolate for mosquito larval control. J. Med. Entomol. 2023, 60, 131–147. [Google Scholar] [CrossRef]
- Kirsch, J.M.; Tay, J.W. Larval Mortality and ovipositional preference in Aedes albopictus (Diptera: Culicidae) induced by the entomopathogenic fungus Beauveria bassiana (Hypocreales: Cordycipitaceae). J. Med. Entomol. 2022, 59, 1687–1693. [Google Scholar] [CrossRef] [PubMed]
- Koodalingam, A.; Dayanidhi, M.K. Studies on biochemical and synergistic effects of immunosuppressive concentration of imidacloprid with Beauveria bassiana and Metarhizium anisopliae for enhancement of virulence against vector mosquito Culex quinquefasciatus. Pestic. Biochem. Physiol. 2021, 176, 104882. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.L.; Dunlap, C.A.; Muturi, E.J.; Barletta, A.B.F.; Rooney, A.P. Entomopathogenic fungal infection leads to temporospatial modulation of the mosquito immune system. PLoS Negl. Trop. Dis. 2018, 12, e0006433. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, J.L.; Schumacher, M.K.; Ower, G.; Palmquist, D.E.; Juliano, S.A. Impacts of fungal entomopathogens on survival and immune responses of Aedes albopictus and Culex pipiens mosquitoes in the context of native Wolbachia infections. PLoS Negl. Trop. Dis. 2021, 15, e0009984. [Google Scholar] [CrossRef]
- Valero-Jiménez, C.A.; Van Kan, J.A.L.; Koenraadt, C.J.M.; Zwaan, B.J.; Schoustra, S.E. Experimental evolution to increase the efficacy of the entomopathogenic fungus Beauveria bassiana against malaria mosquitoes: Effects on mycelial growth and virulence. Evol. Appl. 2017, 10, 433–443. [Google Scholar] [CrossRef]
- Blanford, S.; Jenkins, N.E.; Christian, R.; Chan, B.H.; Nardini, L.; Osae, M.; Koekemoer, L.; Coetzee, M.; Read, A.F.; Thomas, M.B. Storage and persistence of a candidate fungal biopesticide for use against adult malaria vectors. Malar. J. 2012, 11, 354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanford, S.; Jenkins, N.E.; Read, A.F.; Thomas, M.B. Evaluating the lethal and pre-lethal effects of a range of fungi against adult Anopheles stephensi mosquitoes. Malar. J. 2012, 11, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omoya, F.O. Susceptibility of Anopheles arabiensis mosquito pupal stage to bioactivities of Varicosporium elodeae and Articulospora inflata. Biosci. Biotech. Res. Asia 2011, 8, 131–135. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, J.; Rocha, L.F.N.; Martinez, J.M.; Montalva, C.; Humber, R.A.; Luz, C. Clonostachys Spp., Natural mosquito antagonists, and their prospects for biological control of Aedes aegypti. Parasitol. Res. 2022, 121, 2979–2984. [Google Scholar] [CrossRef] [PubMed]
- Montalva, C.; Silva, J.J.; Rocha, L.F.N.; Luz, C.; Humber, R.A. Characterization of Tolypocladium cylindrosporum (Hypocreales, Ophiocordycipitaceae) isolates from Brazil and their efficacy against Aedes aegypti (Diptera, Culicidae). J. Appl. Microbiol. 2019, 126, 266–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrar, A.; Abbas, M.; Mehmood, S.; Ghani, N.; Fatima, A.; Shahzadi, R. Scanning electron microscopy for identification of local strain of Aspergillus parasiticus and its larvicidal efficacy against Aedes aegypti and non-target toxicity testing on fingerlings of Hypophthalmichthys molitrix. Microsc. Res. Tech. 2022, 85, 3187–3192. [Google Scholar] [CrossRef]
- Vasantha-Srinivasan, P.; Karthi, S.; Chellappandian, M.; Ponsankar, A.; Thanigaivel, A.; Senthil-Nathan, S.; Chandramohan, D.; Ganesan, R. Aspergillus flavus (Link) toxins reduces the fitness of dengue vector Aedes aegypti (Linn.) and their non-target toxicity against aquatic predator. Microb. Pathog. 2019, 128, 281–287. [Google Scholar] [CrossRef]
- Podder, D.; Ghosh, S.K. A new application of Trichoderma asperellum as an anopheline larvicide for eco friendly management in medical science. Sci. Rep. 2019, 9, 1108. [Google Scholar] [CrossRef] [Green Version]
- Montalva, C.; Rocha, L.F.N.; Fernandes, É.K.K.; Luz, C.; Humber, R.A. Conidiobolus macrosporus (Entomophthorales), a mosquito pathogen in Central Brazil. J. Invertebr. Pathol. 2016, 139, 102–108. [Google Scholar] [CrossRef]
- Deletre, E.; Martin, T.; Duménil, C.; Chandre, F. Insecticide resistance modifies mosquito response to DEET and natural repellents. Parasit Vectors 2019, 12, 89. [Google Scholar] [CrossRef] [Green Version]
- Samal, R.R.; Panmei, K.; Lanbiliu, P.; Kumar, S. Reversion of CYP450 monooxygenase-mediated acetamiprid larval resistance in dengue fever mosquito, Aedes aegypti L. Bull. Entomol. Res. 2022, 112, 557–566. [Google Scholar] [CrossRef]
- Ismail, H.M.; Freed, S.; Naeem, A.; Malik, S.; Ali, N. The effect of entomopathogenic fungi on enzymatic activity in chlorpyrifos-resistant mosquitoes, Culex quinquefasciatus (Diptera: Culicidae). J. Med. Entomol. 2020, 57, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Zoh, M.G.; Gaude, T.; Prud’homme, S.M.; Riaz, M.A.; David, J.P.; Reynaud, S. Molecular bases of P450-mediated resistance to the neonicotinoid insecticide imidacloprid in the mosquito Ae. Aegypti. Aquat. Toxicol. 2021, 236, 105860. [Google Scholar] [CrossRef]
- Salem, H.H.A.; Mohammed, S.H.; Eltaly, R.I.; Moustafa, M.A.M.; Fónagy, A.; Farag, S.M. Co-application of entomopathogenic fungi with chemical insecticides against Culex pipiens. J. Invertebr. Pathol. 2023, 198, 107916. [Google Scholar] [CrossRef]
- Paula, A.R.; Carolino, A.T.; Paula, C.O.; Samuels, R.I. The combination of the entomopathogenic fungus Metarhizium anisopliae with the insecticide imidacloprid increases virulence against the dengue vector Aedes aegypti (Diptera: Culicidae). Parasit Vectors 2011, 4, 8. [Google Scholar] [CrossRef] [Green Version]
- Araújo, I.F.; Marinho, V.H.D.S.; Sena, I.D.S.; Curti, J.M.; Ramos, R.D.S.; Ferreira, R.M.A.; Souto, R.N.P.; Ferreira, I.M. Larvicidal activity against Aedes aegypti and molecular docking studies of compounds extracted from the endophytic fungus Aspergillus sp. isolated from Bertholletia excelsa Humn. & Bonpl. Biotechnol. Lett. 2022, 44, 439–459. [Google Scholar] [PubMed]
- Noskov, Y.A.; Kabilov, M.R.; Polenogova, O.V.; Yurchenko, Y.A.; Belevich, O.E.; Yaroslavtseva, O.N.; Alikina, T.Y.; Byvaltsev, A.M.; Rotskaya, U.N.; Morozova, V.V.; et al. A neurotoxic insecticide promotes fungal infection in Aedes aegypti larvae by altering the bacterial community. Microb. Ecol. 2021, 81, 493–505. [Google Scholar] [CrossRef] [PubMed]
- Derua, Y.A.; Tungu, P.K.; Malima, R.C.; Mwingira, V.; Kimambo, A.G.; Batengana, B.M.; Machafuko, P.; Sambu, E.Z.; Mgaya, Y.D.; Kisinza, W.N. Laboratory and semi-field evaluation of the efficacy of Bacillus thuringiensis var. israelensis (Bactivec®) and Bacillus sphaericus (Griselesf®) for control of mosquito vectors in northeastern Tanzania. Curr. Res. Parasitol. Vector Borne Dis. 2022, 2, 100089. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Niu, G.; Guan, L.; Ramelow, J.; Zhan, Z.; Zhou, X.; Li, J. Discovery of mosquitocides from fungal extracts through a high-throughput cytotoxicity-screening approach. Parasites Vectors 2021, 14, 595. [Google Scholar] [CrossRef]
- Niu, G.; Wang, B.; Zhang, G.; King, J.B.; Cichewicz, R.H.; Li, J. Targeting mosquito FREP1 with a fungal metabolite blocks malaria transmission. Sci. Rep. 2015, 5, 14694. [Google Scholar] [CrossRef] [Green Version]
- Perera, D.S.; Tharaka, W.G.H.; Amarasinghe, D.; Wickramarachchi, S.R. Extracellular extracts of antagonistic fungi, Trichoderma longibrachiatum and Trichoderma viride, as larvicides against dengue vectors, Aedes aegypti and Aedes albopictus. Acta Trop. 2023, 238, 106747. [Google Scholar] [CrossRef] [PubMed]
- Correa, R.; Coronado, L.M.; Garrido, A.C.; Durant-Archibold, A.A.; Spadafora, C. Volatile organic compounds associated with Plasmodium falciparum infection in vitro. Parasites Vectors 2017, 10, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dormont, L.; Mulatier, M.; Carrasco, D.; Cohuet, A. Mosquito attractants. J. Chem. Ecol. 2021, 47, 351–393. [Google Scholar] [CrossRef] [PubMed]
- Geedi, R.; Canas, L.; Reding, M.E.; Ranger, C.M. Attraction of Myzus persicae (Hemiptera: Aphididae) to volatiles emitted from the entomopathogenic fungus Beauveria bassiana. Environ. Entomol. 2023, 52, 31–38. [Google Scholar] [CrossRef]
- Paula, A.R.; Silva, L.E.I.; Ribeiro, A.; Butt, T.M.; Silva, C.P.; Samuels, R.I. Improving the delivery and efficiency of fungus-impregnated cloths for control of adult Aedes aegypti using a synthetic attractive lure. Parasit Vectors 2018, 11, 285. [Google Scholar] [CrossRef]
- Mostafiz, M.M.; Ryu, J.; Akintola, A.A.; Choi, K.S.; Hwang, U.W.; Hassan, E.; Lee, K.Y. Larvicidal activity of methyl benzoate, a volatile organic compound, against the mosquitoes Aedes albopictus and Culex pipiens (Diptera: Culicidae). J. Med. Entomol. 2022, 59, 788–794. [Google Scholar] [CrossRef]
- De Oliveira Barbosa Bitencourt, R.; De Souza Faria, F.; Marchesini, P.; Reis Dos Santos-Mallet, J.; Guedes Camargo, M.; Rita Elias Pinheiro Bittencourt, V.; Guedes Pontes, E.; Baptista Pereira, D.; Siqueira De Almeida Chaves, D.; Da Costa Angelo, I. Entomopathogenic fungi and Schinus molle essential oil: The combination of two eco-friendly agents against Aedes aegypti larvae. J. Invertebr. Pathol. 2022, 194, 107827. [Google Scholar] [CrossRef]
- Becher, P.G.; Hagman, A.; Verschut, V.; Chakraborty, A.; Rozpędowska, E.; Lebreton, S.; Bengtsson, M.; Flick, G.; Witzgall, P.; Piškur, J. Chemical signaling and insect attraction is a conserved trait in yeasts. Ecol. Evol. 2018, 8, 2962–2974. [Google Scholar] [CrossRef] [Green Version]
- Hapairai, L.K.; Mysore, K.; James, L.D.; Scheel, N.D.; Realey, J.S.; Sun, L.; Gerber, L.E.; Feng, R.S.; Romero-Severson, E.; Mohammed, A.; et al. Evaluation of large volume yeast interfering RNA lure-and-kill ovitraps for attraction and control of Aedes mosquitoes. Med. Vet. Entomol. 2021, 35, 361–370. [Google Scholar] [CrossRef]
- Zamora-Avilés, N.; Orozco-Flores, A.A.; Gomez-Flores, R.; Domínguez-Gámez, M.; Rodríguez-Pérez, M.A.; Tamez-Guerra, P. Increased attraction and stability of Beauveria bassiana-formulated microgranules for Aedes aegypti Biocontrol. J. Fungi 2022, 8, 828. [Google Scholar] [CrossRef]
- Schorkopf, D.L.P.; Spanoudis, C.G.; Mboera, L.E.G.; Mafra-Neto, A.; Ignell, R.; Dekker, T. Combining attractants and larvicides in biodegradable matrices for sustainable mosquito vector control. PLoS Negl. Trop. Dis. 2016, 10, e0005043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girard, M.; Martin, E.; Vallon, L.; Raquin, V.; Bellet, C.; Rozier, Y.; Desouhant, E.; Hay, A.-E.; Luis, P.; Valiente Moro, C.; et al. Microorganisms associated with mosquito oviposition sites: Implications for habitat selection and insect life histories. Microorganisms 2021, 9, 1589. [Google Scholar] [CrossRef]
- Ponnusamy, L.; Schal, C.; Wesson, D.M.; Arellano, C.; Apperson, C.S. Oviposition responses of Aedes mosquitoes to bacterial isolates from attractive bamboo infusions. Parasites Vectors 2015, 8, 486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eneh, L.K.; Saijo, H.; Borg-Karlson, A.-K.; Lindh, J.M.; Rajarao, G.K. Cedrol, a malaria mosquito oviposition attractant is produced by fungi isolated from rhizomes of the grass Cyperus rotundus. Malar. J. 2016, 15, 478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaalan, E.A.; Canyon, D.V. Aquatic insect predators and mosquito control. Trop. Biomed. 2009, 26, 223–261. [Google Scholar]
- Vinogradov, D.D.; Sinev, A.Y.; Tiunov, A.V. Predators as control agents of mosquito larvae in micro-reservoirs (review). Inland Water Biol. 2022, 15, 39–53. [Google Scholar] [CrossRef]
- Karthi, S.; Vasantha-Srinivasan, P.; Ganesan, R.; Ramasamy, V.; Senthil-Nathan, S.; Khater, H.F.; Radhakrishnan, N.; Amala, K.; Kim, T.J.; El-Sheikh, M.A.; et al. Target activity of Isaria tenuipes (Hypocreales: Clavicipitaceae) fungal strains against dengue vector Aedes aegypti (Linn.) and its non-target activity against aquatic predators. J. Fungi 2020, 6, 196. [Google Scholar] [CrossRef]
- Alkhaibari, A.M.; Maffeis, T.; Bull, J.C.; Butt, T.M. Combined use of the entomopathogenic fungus, Metarhizium brunneum, and the mosquito predator, Toxorhynchites brevipalpis, for control of mosquito larvae: Is this a risky biocontrol strategy? J. Invertebr. Pathol. 2018, 153, 38–50. [Google Scholar] [CrossRef]
- Ong’wen, F.; Onyango, P.O.; Bukhari, T. Direct and indirect effects of predation and parasitism on the Anopheles gambiae mosquito. Parasites Vectors 2020, 13, 43. [Google Scholar] [CrossRef] [Green Version]
- Kamareddine, L.; Fan, Y.; Osta, M.A.; Keyhani, N.O. Expression of trypsin modulating oostatic factor (TMOF) in an entomopathogenic fungus increases its virulence towards Anopheles gambiae and reduces fecundity in the target mosquito. Parasites Vectors 2013, 6, 22. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; St. Leger, R.J. A collagenous protective coat enables Metarhizium anisopliae to evade insect immune responses. Proc. Natl. Acad. Sci. USA 2006, 103, 6647–6652. [Google Scholar] [CrossRef]
- Bilgo, E.; Lovett, B.; Fang, W.; Bende, N.; King, G.F.; Diabate, A.; St. Leger, R.J. Improved efficacy of an arthropod toxin expressing fungus against insecticide-resistant malaria-vector mosquitoes. Sci. Rep. 2017, 7, 3433. [Google Scholar] [CrossRef] [PubMed]
- Bilgo, E.; Lovett, B.; Bayili, K.; Millogo, A.S.; Saré, I.; Dabiré, R.K.; Sanon, A.; St. Leger, R.J.; Diabate, A. Transgenic Metarhizium pingshaense synergistically ameliorates pyrethroid-resistance in wild-caught, malaria-vector mosquitoes. PLoS ONE 2018, 13, e0203529. [Google Scholar] [CrossRef] [PubMed]
- Cui, G.; Huang, C.; Bi, X.; Wang, Y.; Yin, K.; Zhu, L.; Jiang, Z.; Chen, B.; Deng, Y.Z. Aminotransferase SsAro8 regulates tryptophan metabolism essential for filamentous growth of sugarcane smut fungus Sporisorium scitamineum. Microbiol. Spectr. 2022, 10, e00570-22. [Google Scholar] [CrossRef]
- Tu, C.; Zhang, Y.; Zhu, P.; Sun, L.; Xu, P.; Wang, T.; Luo, J.; Yu, J.; Xu, L. Enhanced toxicity of entomopathogenic fungi Beauveria bassiana with bacteria expressing immune suppressive dsRNA in a leaf beetle. Pestic. Biochem. Physiol. 2023, 193, 105431. [Google Scholar] [CrossRef]
- Yordanova, I.A.; Zakovic, S.; Rausch, S.; Costa, G.; Levashina, E.; Hartmann, S. Micromanaging immunity in the murine host vs. the mosquito vector: Microbiota-dependent immune responses to intestinal parasites. Front. Cell. Infect. Microbiol. 2018, 8, 308. [Google Scholar] [CrossRef]
- Wang, S.; Jacobs-Lorena, M. Genetic approaches to interfere with malaria transmission by vector mosquitoes. Trends Biotechnol. 2013, 31, 185–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mancini, M.V.; Spaccapelo, R.; Damiani, C.; Accoti, A.; Tallarita, M.; Petraglia, E.; Rossi, P.; Cappelli, A.; Capone, A.; Peruzzi, G.; et al. Paratransgenesis to control malaria vectors: A semi-field pilot study. Parasites Vectors 2016, 9, 140. [Google Scholar] [CrossRef] [Green Version]
- Fang, W.; Vega-Rodríguez, J.; Ghosh, A.K.; Jacobs-Lorena, M.; Kang, A.; St. Leger, R.J. Development of transgenic fungi that kill human malaria parasites in mosquitoes. Science 2011, 331, 1074–1077. [Google Scholar] [CrossRef] [Green Version]
- Shane, J.L.; Grogan, C.L.; Cwalina, C.; Lampe, D.J. Blood meal-induced inhibition of vector-borne disease by transgenic microbiota. Nat. Commun. 2018, 9, 4127. [Google Scholar] [CrossRef] [Green Version]
- Falvo, M.L.; Albornoz Medina, P.; Rodrigues, J.; López Lastra, C.C.; García, J.J.; Fernandes, É.K.K.; Luz, C. Effect of UV-B irradiation on water-suspended Metarhizium anisopliae s.l. (Hypocreales: Clavicipitaceae) conidia and their larvicidal activity in Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2018, 55, 1330–1333. [Google Scholar] [CrossRef]
- Falvo, M.L.; Pereira-Junior, R.A.; Rodrigues, J.; López Lastra, C.C.; García, J.J.; Fernandes, É.K.K.; Luz, C. UV-B Radiation reduces in vitro germination of Metarhizium anisopliae s.l. but does not affect virulence in fungus-treated Aedes aegypti adults and development on dead mosquitoes. J. Appl. Microbiol. 2016, 121, 1710–1717. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.; St. Leger, R.J. Enhanced UV resistance and improved killing of malaria mosquitoes by photolyase transgenic entomopathogenic fungi. PLoS ONE 2012, 7, e43069. [Google Scholar] [CrossRef] [Green Version]
- Lovett, B.; St. Leger, R.J. Stress is the rule rather than the exception for Metarhizium. Curr. Genet. 2015, 61, 253–261. [Google Scholar] [CrossRef]
- Vandenberg, J.D.; Cantone, F.A. Effect of serial transfer of three strains of Paecilomyces fumosoroseus on growth in vitro, virulence, and host specificity. J. Invertebr. Pathol. 2004, 85, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Hu, X.; Huang, H. Recent patents on biosafety strategies of selectable marker genes in genetically modified crops. Recent. Pat. Food Nutr. Agric. 2014, 6, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, S.; Turna, J. Excision of selectable marker genes from transgenic crops as a concern for environmental biosafety. J. Sci. Food Agric. 2007, 87, 2547–2554. [Google Scholar] [CrossRef]
- Ying, S.H.; Feng, M.G.; Keyhani, N.O. Use of uridine auxotrophy (ura3) for markerless transformation of the mycoinsecticide Beauveria bassiana. Appl. Microbiol. Biotechnol. 2013, 97, 3017–3025. [Google Scholar] [CrossRef]
- Rovner, A.J.; Haimovich, A.D.; Katz, S.R.; Li, Z.; Grome, M.W.; Gassaway, B.M.; Amiram, M.; Patel, J.R.; Gallagher, R.R.; Rinehart, J.; et al. Recoded organisms engineered to depend on synthetic amino acids. Nature 2015, 518, 89–93. [Google Scholar] [CrossRef] [Green Version]
- Fang, W.; Lu, H.L.; King, G.F.; St. Leger, R.J. Construction of a hypervirulent and specific mycoinsecticide for locust control. Sci. Rep. 2014, 4, 7345. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, M.R.; Katak, R.D.M.; Da Silva, G.F.; Marinotti, O.; Terenius, O.; Tadei, W.P.; De Souza, A.D.L.; De Souza, A.Q.L. Extracts of Amazonian fungi with larvicidal activities against Aedes aegypti. Front. Microbiol. 2021, 12, 743246. [Google Scholar] [CrossRef] [PubMed]
- Vyas, N.; Dua, K.; Prakash, S. Larvicidal activity of metabolites of Metarhizium anisopliae against Aedes and Culex mosquitoes. Entomol. Ornithol. Herpetol. 2015, 4, 162. [Google Scholar] [CrossRef] [Green Version]
- Ragavendran, C.; Manigandan, V.; Kamaraj, C.; Balasubramani, G.; Prakash, J.S.; Perumal, P.; Natarajan, D. Larvicidal, histopathological, antibacterial activity of indigenous fungus Penicillium sp. against Aedes aegypti L and Culex quinquefasciatus (Say) (Diptera: Culicidae) and its acetylcholinesterase inhibition and toxicity assessment of Zebrafish (Danio rerio). Front. Microbiol. 2019, 10, 427. [Google Scholar] [PubMed] [Green Version]
- Baskar, K.; Chinnasamy, R.; Pandy, K.; Venkatesan, M.; Sebastian, P.J.; Subban, M.; Thomas, A.; Kweka, E.J.; Devarajan, N. Larvicidal and histopathology effect of endophytic fungal extracts of Aspergillus tamarii against Aedes aegypti and Culex quinquefasciatus. Heliyon 2020, 6, e05331. [Google Scholar] [CrossRef] [PubMed]
- Ragavendran, C.; Mariappan, T.; Natarajan, D. Larvicidal, histopathological efficacy of Penicillium daleae against larvae of Culex quinquefasciatus and Aedes aegypti plus biotoxicity on Artemia nauplii a non-target aquatic organism. Front. Pharmacol. 2017, 8, 773. [Google Scholar] [CrossRef] [Green Version]
- Arunthirumeni, M.; Vinitha, G.; Shivakumar, M.S. Antifeedant and larvicidal activity of bioactive compounds isolated from entomopathogenic fungi Penicillium sp. for the control of agricultural and medically important insect pest (Spodoptera litura and Culex quinquefasciatus). Parasitol. Int. 2023, 92, 83102688. [Google Scholar] [CrossRef]


{kind=link}
| Fungal Strain | Stage | Mosquito Species | Mosquito Killing Effect (Mortality) * | Reference |
|---|---|---|---|---|
| M. humberi | Egg | Ae. aegypti | 25–30% eclosion | [29] |
| M. anisopliae | Pupae | Ae. aegypti | 1 × 107 conidia/mL: 43%/24 h; 77%/48 h | [33] |
| Larvae | Ae. aegypti | 1 × 108 spores/mL: ST50 = 48 h | [43] | |
| Larvae | Cx. pipiens | 1 × 108 conidia/mL: 88%; LT50 = 22.6 h | [55] | |
| Larvae | Cx. qinquefasciatus | 1 × 106–1 × 1010 conidia/mL; LT50 = 3.25 d | [47] | |
| Larvae | Ae. albopictus | Pupation was delayed by 2.75% | [15] | |
| Larvae | An. Stephensi | Pupation was delayed by 83.3% | [12] | |
| Adults | Ae. aegypti | LC50 = 2.4 conidia/mL: ST50 = 5 d | [56] | |
| Adults | An. gambiae | 1 × 1011 conidia/m2: 100%/7 d | [57] | |
| Adults | An. stephensi | 1 × 107 conidia/mL: 57.5%/ST50 = 10 d | [12] | |
| B. bassiana | Pupae | An. gambiae | No effects on pupae | [58] |
| Pupae | Ae. albopictus | 2.5 × 108 conidia/mL: 14.0–40.5% | [58] | |
| Larvae | Ae. aegypti | 1 × 108 conidia/mL: ST50 = 2 d | [43] | |
| Larvae | Ae. albopictus | 1 × 106 conidia/mL: LT50 = 3.68 d | [59] | |
| Larvae | An. gambiae | 1.25–2.5 × 108 conidia/mL: 97.2–100% | [58] | |
| Larvae | An. stephensi | 1 × 106–1 × 1010 conidia/mL: LT50 = 6.18 d | [47] | |
| Larvae | Cx. pipiens | 1 × 108 conidia/m:73.33%; LT50 = 38.35 h | [55] | |
| Larvae | Cx. qinquefasciatus | 1 × 107 conidia/mL; 36.47%/92 h | [60] | |
| Adults | Ae. albopictus | 5 × 108 conidia/mL: S50 = 5 d | [14] | |
| Adults | Ae. aegypti | 1 × 108 conidia/mL: 95%/11 d; LT50 = 4.5 d | [61] | |
| Adults | Cx. pipiens | 1 × 108 conidia/mL: LT50 = 7.9 d | [62] | |
| Adults | An. coluzzii | 1 × 108 conidia/mL: ST50 = 5–7 d | [63] | |
| Adults | An. gambiae | 1 × 106 spores/mL: LT50 = 5 d | [64] | |
| Adults | An. stephensi | 1 × 106 conidia/mL: LT50 = 4 d | [65] | |
| V. elodeae | Pupae | An. gambiae | LC50 = 2.64 sfu/mL | [66] |
| A. inflata | Pupae | An. gambiae | LC50 = 5.486 sfu/mL | [66] |
| C. eriocamporesii | Eggs | Ae. aegypti | 5 × 106 conidia/cm2: 89% eclosion | [67] |
| Larvae | Ae. aegypti | 1 × 107 conidia/mL: LT50 = 0.9 d | [67] | |
| Adults | Ae. aegypti | 1 × 107 conidia/cm2: LT50 = 18.2 d | [67] | |
| T. cylindrosporum | Eggs | Ae. aegypti | 1 × 105 conidia/cm2: 85% eclosion | [68] |
| A. parasiticus | Larvae | Ae. Aegypti | LC50 = 1.0 × 107 conidia/mL; 24 h | [69] |
| LC50 = 2.99 × 105 conidia/mL; 48 h | ||||
| A. flavus | Larvae | Ae. aegypti | 2 × 108 conidia/mL: >90% | [70] |
| T. asperellum | Larvae | Aedes spp. | 2.68 × 108 conidia/mL; LT50 = 12.33 h | [71] |
| C. clavisporus | Larvae | An. stephensi | 1 × 106 conidia/mL: LT50 = 1.3 d | [9] |
| A. clavatus | Larvae | Cx. quinquefasciatus | 0.5–2.5 × 108 spores/mL: 17.0–74.3%/48 h | [9] |
| Ae. aegypti | ||||
| An. gambiae | ||||
| C. macrosporus | Larvae | Ae. aegypti | 8.3 × 104 conidia/cm2: 100%/72 h | [72] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, Y.; Liu, X.; Peng, G.; Xia, Y.; Cao, Y. Recent Advancements in Pathogenic Mechanisms, Applications and Strategies for Entomopathogenic Fungi in Mosquito Biocontrol. J. Fungi 2023, 9, 746. https://doi.org/10.3390/jof9070746
Qin Y, Liu X, Peng G, Xia Y, Cao Y. Recent Advancements in Pathogenic Mechanisms, Applications and Strategies for Entomopathogenic Fungi in Mosquito Biocontrol. Journal of Fungi. 2023; 9(7):746. https://doi.org/10.3390/jof9070746
Chicago/Turabian StyleQin, Yujie, Xiaoyu Liu, Guoxiong Peng, Yuxian Xia, and Yueqing Cao. 2023. "Recent Advancements in Pathogenic Mechanisms, Applications and Strategies for Entomopathogenic Fungi in Mosquito Biocontrol" Journal of Fungi 9, no. 7: 746. https://doi.org/10.3390/jof9070746
APA StyleQin, Y., Liu, X., Peng, G., Xia, Y., & Cao, Y. (2023). Recent Advancements in Pathogenic Mechanisms, Applications and Strategies for Entomopathogenic Fungi in Mosquito Biocontrol. Journal of Fungi, 9(7), 746. https://doi.org/10.3390/jof9070746