
Antimicrobial Activity the Essential Oil from Croton pluriglandulosus Carn. Leaves against Microorganisms of Clinical Interest

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. Plant Material and Extraction of Essential Oil
2.3. Characterization of CpEO by GC/MS Analysis
2.4. Antimicrobial Activity
2.5. Antibiofilm Assay
2.6. Mechanisms of Action
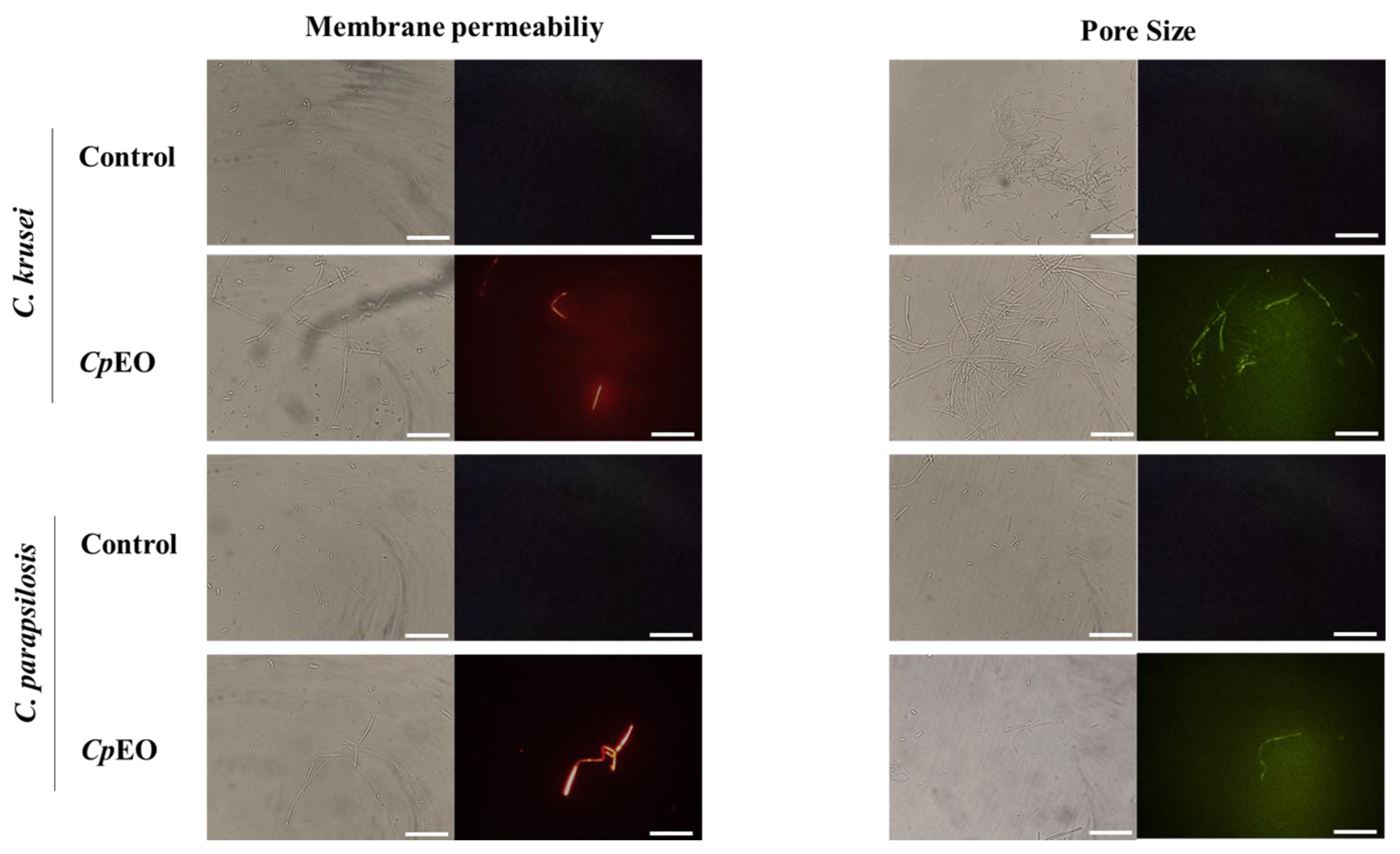
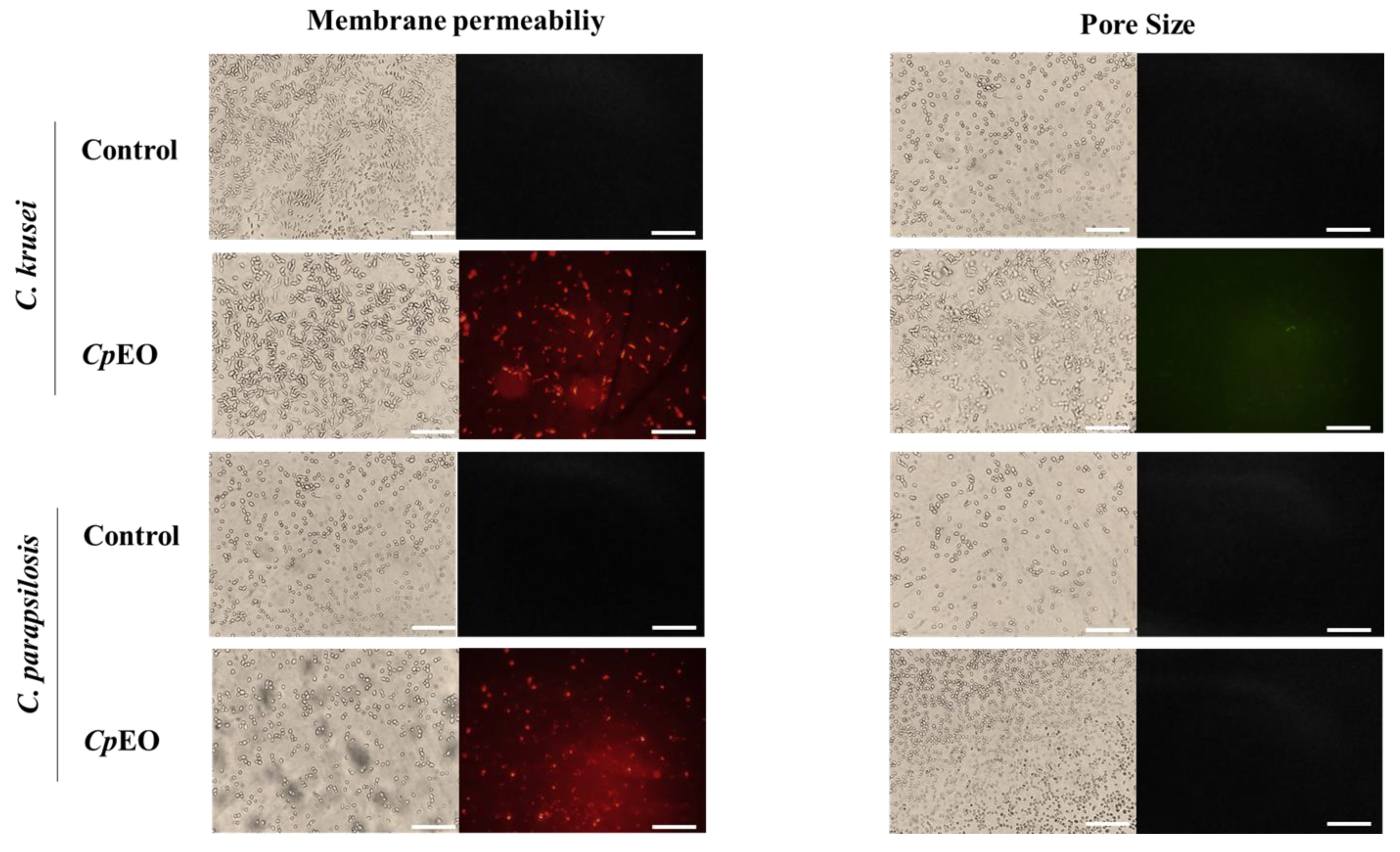
2.6.1. Cell Membrane Integrity
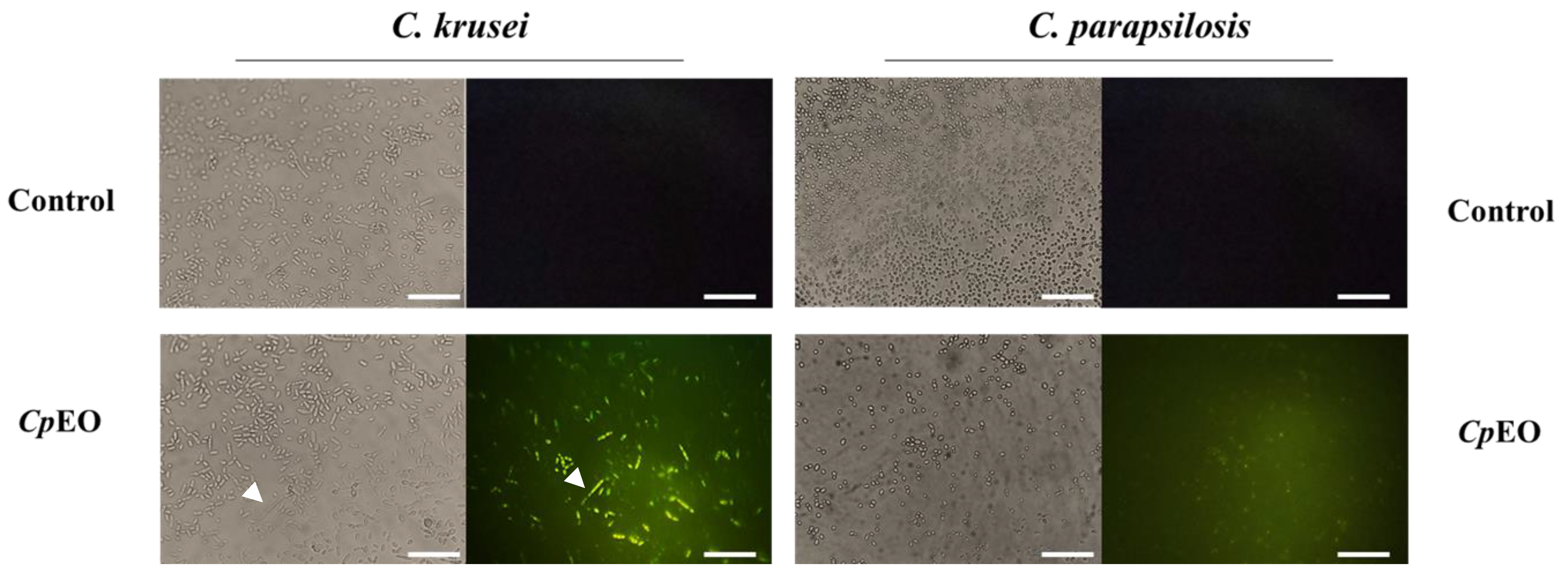
2.6.2. Detection of ROS Overproduction
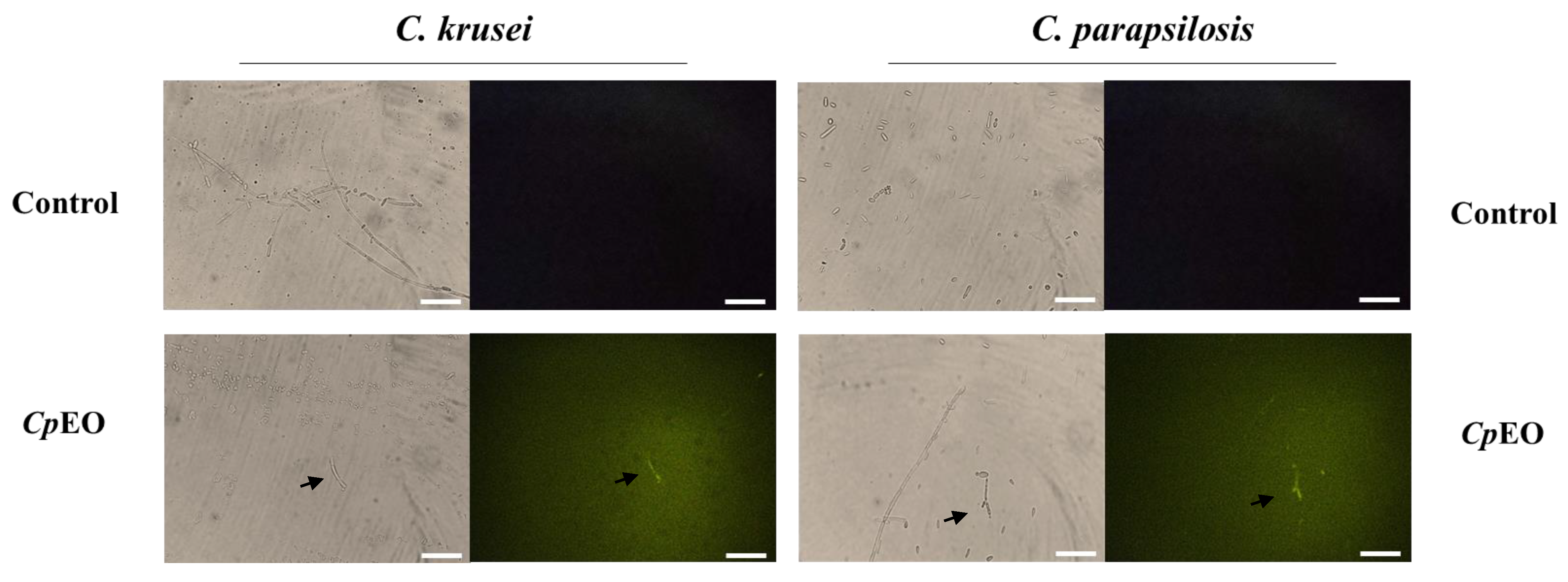
2.6.3. Induction of Apoptosis
2.7. Scanning Electron Microscopy (SEM)
2.8. Hemolytic Analysis
2.9. Statistical Analysis
3. Results and Discussion
3.1. GC-MS/MS Analysis of CpEO
3.2. Antimicrobial Activity of CpEO
3.3. Mechanisms of Action
3.3.1. Cell Membrane Integrity
3.3.2. ROS Overproduction and Induction of Apoptosis
3.4. Scanning Electron Microscopy (SEM)
3.5. Hemolytic Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Butler, G.; Rasmussen, M.D.; Lin, M.F.; Santos, M.A.S.; Sakthikumar, S.; Munro, C.A.; Rheinbay, E.; Grabherr, M.; Forche, A.; Reedy, J.L.; et al. Evolution of Pathogenicity and Sexual Reproduction in Eight Candida Genomes. Nature 2009, 459, 657–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, J.; Ding, Y.; Wei, B.; Wang, L.; Xu, S.; Qin, P.; Wei, L.; Jiang, L. Epidemiology of Candida Albicans and Non-C.Albicans of Neonatal Candidemia at a Tertiary Care Hospital in Western China. BMC Infect. Dis. 2017, 17, 329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, I.d.O.; Oliveira, R.d.A.G.; Lima, E.d.O.; Farias, N.M.P.; de Souza, E.L. Atividade Antifúngica de Óleos Essenciais Sobre Espécies de Candida. Rev. Bras. Farmacogn. 2006, 16, 197–201. [Google Scholar] [CrossRef]
- Vieira, F.; Nascimento, T. Resistência a Fármacos Antifúngicos Por Candida e Abordagem Terapêutica. Rev. Port. Farmacoter. 2017, 9, 29–36. [Google Scholar]
- Arendrup, M.C.; Patterson, T.F. Multidrug-Resistant Candida: Epidemiology, Molecular Mechanisms, and Treatment. J. Infect. Dis. 2017, 216, S445–S451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- do Vale, J.P.C.; Vasconcelos, M.A.; Arruda, F.V.S.; Firmino, N.C.S.; Pereira, A.L.; Andrade, A.L.; Saker-Sampaio, S.; Sampaio, A.H.; Marinho, E.S.; Teixeira, A.M.R.; et al. Evaluation of Antimicrobial and Antioxidant Potential of Essential Oil from Croton Piauhiensis Müll. Arg. Curr. Microbiol. 2021, 78, 1926–1938. [Google Scholar] [CrossRef] [PubMed]
- Benini, P.C.; Schwan-Estrada, K.R.F.; Klais, E.C.; Cruz, M.E.S.; Itako, A.T.; Mesquini, R.M.; Stangarlin, J.R.; Tolentino Júnior, J.B. Efeito in vitro do óleo essencial e extrato aquoso de Ocimum gratissimum colhido nas quatro estações do ano sobre fitopatógenos. Arq. Inst. Biol. 2010, 77, 677–683. [Google Scholar] [CrossRef]
- Souza, A.E.F.; Araújo, E.; Nascimento, L.C. Atividade Antifúngica de Extratos de Alho e Capim-Santo Sobre o Desenvolvimento de Fusarium Proliferatum Isolado de Grãos de Milho. Fitopatol. Bras. 2007, 32, 465–471. [Google Scholar] [CrossRef]
- Dudareva, N.; Klempien, A.; Muhlemann, J.K.; Kaplan, I. Biosynthesis, Function and Metabolic Engineering of Plant Volatile Organic Compounds. New Phytol. 2013, 198, 16–32. [Google Scholar] [CrossRef]
- Arrais, L.G.; Lyra, H.F.S.; Batista, D.C.A.; Coutinho, F.N.; Saraiva, A.M.; Pereira, R.C.A.; Pisciottano, M.N.C.; Xavier, H.S.; Melo, S.J. Atividade Antimicrobiana Dos Extratos Metanólicos Da Raiz, Caule e Folhas de Croton Pulegioides Baill. (Zabelê). Rev. Bras. Plantas Med. 2014, 16, 316–322. [Google Scholar] [CrossRef] [Green Version]
- Guerra, A.M.N.d.M.; Silva, D.d.S.; Santos, P.S.; dos Santos, L.B. Teste de Repelência de Óleos Essenciais Sobre Callosobruchus Maculatus. Rev. Bras. Agropecu. Sustent. 2019, 9, 583–586. [Google Scholar] [CrossRef]
- Da Costa, J.G.M.; Rodrigues, F.F.G.; Angélico, E.C.; Pereira, C.K.B.; de Souza, E.O.; Caldas, G.F.R.; Silva, M.R.; Santos, N.K.A.; Mota, M.L.; dos Santos, P.F. Composição Química e Avaliação Da Atividade Antibacteriana e Toxicidade Do Óleo Essencial de Croton Zehntneri (Variedade Estragol). Rev. Bras. Farmacogn. 2008, 18, 583–586. [Google Scholar] [CrossRef] [Green Version]
- De Vasconcelos, E.C.; Paganini, C.C.; de Figueiredo, E.A.T.; de Aragão, G.M.F. Ação Antimicrobiana Dos Compostos Voláteis Do Óleo Essencial Das Folhas Secas de Croton Blanchetianus Baill. Res. Soc. Dev. 2022, 11, e28011124785. [Google Scholar] [CrossRef]
- Riina, R.; Van Ee, B.W.; Beatriz Rossi Caruzo, M.; Santos Carneiro-Torres, D.; Freitas dos Santos, R.; Berry, P.E. The neotropical croton section geiseleria (Euphorbiaceae): Classification update, phylogenetic framework, and seven new species from South America 1. Ann. Mo. Bot. Gard. 2021, 106, 111–166. [Google Scholar] [CrossRef]
- De Oliveira, A.R.M.F.; Jezler, C.N.; Oliveira, R.A.; Mielke, M.S.; Costa, L.C.B. Determinação Do Tempo de Hidrodestilação e Do Horário de Colheita No Óleo Essencial de Menta. Hortic. Bras. 2012, 30, 155–159. [Google Scholar] [CrossRef] [Green Version]
- Malveira, E.A.; Souza, P.F.N.; Neto, N.A.S.; Aguiar, T.K.B.; Rodrigues, N.S.; Henrique, C.W.B.; Silva, A.F.B.; Lima, L.B.; Albuquerque, C.C.; Freitas, C.D.T. Essential Oil from Croton Blanchetianus Leaves: Anticandidal Potential and Mechanisms of Action. J. Fungi 2022, 8, 1147. [Google Scholar] [CrossRef]
- Waterman, P.G. Identification of Essential Oil Components by Gas Chromatography/Mass Spectroscopy. Biochem. Syst. Ecol. 1996, 6, 594. [Google Scholar] [CrossRef]
- Souza, P.F.N.; Marques, L.S.M.; Oliveira, J.T.A.; Lima, P.G.; Dias, L.P.; Neto, N.A.S.; Lopes, F.E.S.; Sousa, J.S.; Silva, A.F.B.; Caneiro, R.F.; et al. Synthetic Antimicrobial Peptides: From Choice of the Best Sequences to Action Mechanisms. Biochimie 2020, 175, 132–145. [Google Scholar] [CrossRef]
- Dias, L.P.; Souza, P.F.N.; Oliveira, J.T.A.; Vasconcelos, I.M.; Araújo, N.M.S.; Tilburg, M.F.V.; Guedes, M.I.F.; Carneiro, R.F.; Lopes, J.L.S.; Sousa, D.O.B. RcAlb-PepII, a Synthetic Small Peptide Bioinspired in the 2S Albumin from the Seed Cake of Ricinus Communis, Is a Potent Antimicrobial Agent against Klebsiella Pneumoniae and Candida Parapsilosis. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183092. [Google Scholar] [CrossRef]
- Oliveira, J.T.A.; Souza, P.F.N.; Vasconcelos, I.M.; Dias, L.P.; Martins, T.F.; Van Tilburg, M.F.; Guedes, M.I.F.; Sousa, D.O.B. Mo-CBP3-PepI, Mo-CBP3-PepII, and Mo-CBP3-PepIII Are Synthetic Antimicrobial Peptides Active against Human Pathogens by Stimulating ROS Generation and Increasing Plasma Membrane Permeability. Biochimie 2019, 157, 10–21. [Google Scholar] [CrossRef]
- Dikalov, S.I.; Harrison, D.G. Methods for Detection of Mitochondrial and Cellular Reactive Oxygen Species. Antioxid. Redox Signal. 2014, 20, 372–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staniszewska, M.; Bondaryk, M.; Swoboda-Kopec, E.; Siennicka, K.; Sygitowicz, G.; Kurzatkowski, W. Candida Albicans Morphologies Revealed by Scanning Electron Microscopy Analysis. Braz. J. Microbiol. 2013, 44, 813–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharifi-Rad, J.; Sureda, A.; Tenore, G.; Daglia, M.; Sharifi-Rad, M.; Valussi, M.; Tundis, R.; Sharifi-Rad, M.; Loizzo, M.; Ademiluyi, A.; et al. Biological Activities of Essential Oils: From Plant Chemoecology to Traditional Healing Systems. Molecules 2017, 22, 70. [Google Scholar] [CrossRef] [PubMed]
- Rossi, P.-G.; Bao, L.; Luciani, A.; Panighi, J.; Desjobert, J.-M.; Costa, J.; Casanova, J.; Bolla, J.-M.; Berti, L. (E)-Methylisoeugenol and Elemicin: Antibacterial Components of Daucus Carota L. Essential Oil against Campylobacter Jejuni. J. Agric. Food Chem. 2007, 55, 7332–7336. [Google Scholar] [CrossRef] [PubMed]
- Saldanha, A.A.; Vieira, L.; Ribeiro, R.I.M.d.A.; Thomé, R.G.; dos Santos, H.B.; Silva, D.B.; Carollo, C.A.; de Oliveira, F.M.; Lopes, D.d.O.; de Siqueira, J.M.; et al. Chemical Composition and Evaluation of the Anti-Inflammatory and Antinociceptive Activities of Duguetia Furfuracea Essential Oil: Effect on Edema, Leukocyte Recruitment, Tumor Necrosis Factor Alpha Production, INOS Expression, and Adenosinergic and Opioid. J. Ethnopharmacol. 2019, 231, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Nogueira Sobrinho, A.C.; de Morais, S.M.; Marinho, M.M.; de Souza, N.V.; Lima, D.M. Antiviral Activity on the Zika Virus and Larvicidal Activity on the Aedes Spp. of Lippia Alba Essential Oil and β-Caryophyllene. Ind. Crops Prod. 2021, 162, 113281. [Google Scholar] [CrossRef]
- Maia, D.S.; Lopes, C.F.; Saldanha, A.A.; Silva, N.L.; Sartori, Â.L.B.; Carollo, C.A.; Sobral, M.G.; Alves, S.N.; Silva, D.B.; de Siqueira, J.M. Larvicidal Effect from Different Annonaceae Species on Culex Quinquefasciatus. Environ. Sci. Pollut. Res. 2020, 27, 36983–36993. [Google Scholar] [CrossRef]
- Carev, I.; Gelemanović, A.; Glumac, M.; Tutek, K.; Dželalija, M.; Paiardini, A.; Prosseda, G. Centaurea Triumfetii Essential Oil Chemical Composition, Comparative Analysis, and Antimicrobial Activity of Selected Compounds. Sci. Rep. 2023, 13, 7475. [Google Scholar] [CrossRef]
- Dahham, S.S.; Tabana, Y.M.; Iqbal, M.A.; Ahamed, M.B.K.; Ezzat, M.O.; Majid, A.S.A.; Majid, A.M.S.A. The Anticancer, Antioxidant and Antimicrobial Properties of the Sesquiterpene β-Caryophyllene from the Essential Oil of Aquilaria Crassna. Molecules 2015, 20, 11808–11829. [Google Scholar] [CrossRef] [Green Version]
- Seol, G.H.; Kim, K.Y. Eucalyptol and Its Role in Chronic Diseases. In Drug Discovery from Mother Nature; Springer: Berlin/Heidelberg, Germany, 2016; pp. 389–398. [Google Scholar]
- Mishra, P.; Gupta, P.; Srivastava, A.K.; Poluri, K.M.; Prasad, R. Eucalyptol/β-Cyclodextrin Inclusion Complex Loaded Gellan/PVA Nanofibers as Antifungal Drug Delivery System. Int. J. Pharm. 2021, 609, 121163. [Google Scholar] [CrossRef]
- Sousa, O.V.; Del-Vechio-Vieira, G.; Alves, M.S.; Araújo, A.A.L.; Pinto, M.A.O.; Amaral, M.P.H.; Rodarte, M.P.; Kaplan, M.A.C. Chemical Composition and Biological Activities of the Essential Oils from Duguetia Lanceolata St. Hil. Barks. Molecules 2012, 17, 11056–11066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, C.-Y.; Yang, W.; Li, M.; Ying, J.; Tao, S.-J.; Li, K.; Dong, J.-H.; Wang, X.-S. Cell Apoptosis Induced by δ-Elemene in Colorectal Adenocarcinoma Cells via a Mitochondrial-Mediated Pathway. YAKUGAKU ZASSHI 2009, 129, 1403–1413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moremi, M.P.; Kamatou, G.P.; Viljoen, A.M.; Tankeu, S.Y. Croton Gratissimus—Essential Oil Composition and Chemometric Analysis of an Ethnomedicinally Important Tree from South Africa. S. Afr. J. Bot. 2021, 138, 141–147. [Google Scholar] [CrossRef]
- Moremi, M.P.; Makolo, F.; Viljoen, A.M.; Kamatou, G.P. A Review of Biological Activities and Phytochemistry of Six Ethnomedicinally Important South African Croton Species. J. Ethnopharmacol. 2021, 280, 114416. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Chalchat, J.C.; Michet, A.; Linhart, Y.B.; Ehlers, B. Qualitative and Quantitative Variation in Monoterpene Co-Occurrence and Composition in the Essential Oil of Thymus Vulgaris Chemotypes. J. Chem. Ecol. 2003, 29, 859–880. [Google Scholar] [CrossRef] [PubMed]
- Yammine, J.; Chihib, N.-E.; Gharsallaoui, A.; Dumas, E.; Ismail, A.; Karam, L. Essential Oils and Their Active Components Applied as: Free, Encapsulated and in Hurdle Technology to Fight Microbial Contaminations. A Review. Heliyon 2022, 8, e12472. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.K.; Yusoff, K.; Thomas, W.; Akseer, R.; Alhosani, M.S.; Abushelaibi, A.; Lim, S.H.E.; Lai, K.S. Lavender Essential Oil Induces Oxidative Stress Which Modifies the Bacterial Membrane Permeability of Carbapenemase Producing Klebsiella Pneumoniae. Sci. Rep. 2020, 10, 819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, S.; Sae-Tia, S.; Fries, B.C. Candidiasis and Mechanisms of Antifungal Resistance. Antibiotics 2020, 9, 312. [Google Scholar] [CrossRef] [PubMed]
- Asdadi, A.; Hamdouch, A.; Oukacha, A.; Moutaj, R.; Gharby, S.; Harhar, H.; El Hadek, M.; Chebli, B.; Idrissi Hassani, L.M. Study on Chemical Analysis, Antioxidant and in Vitro Antifungal Activities of Essential Oil from Wild Vitex Agnus-Castus L. Seeds Growing in Area of Argan Tree of Morocco against Clinical Strains of Candida Responsible for Nosocomial Infections. J. Mycol. Med. 2015, 25, e118–e127. [Google Scholar] [CrossRef]
- Taweechaisupapong, S.; Aieamsaard, J.; Chitropas, P.; Khunkitti, W. Inhibitory Effect of Lemongrass Oil and Its Major Constituents on Candida Biofilm and Germ Tube Formation. S. Afr. J. Bot. 2012, 81, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Da Costa, A.C.V.; Melo, G.F.A.; Madruga, M.S.; da Costa, J.G.M.; Garino Junior, F.; Queiroga Neto, V. Chemical Composition and Antibacterial Activity of Essential Oil of a Croton Rhamnifolioides Leaves Pax & Hoffm. Semin. Ciências Agrárias 2013, 34, 2853. [Google Scholar] [CrossRef] [Green Version]
- Filho, A.C.; Castro, C.F.; da Silva, A.; da Cruz, R. Efeito Antifúngico Sobre Candida Pelo Óleo Essencial Da Flor de Himatanthus Obovathus (Müell. Arg.) Woodson (Apocynaceae). Rev. Arq. Científicos 2021, 4, 72–78. [Google Scholar]
- Tariq, S.; Wani, S.; Rasool, W.; Shafi, K.; Bhat, M.A.; Prabhakar, A.; Shalla, A.H.; Rather, M.A. A Comprehensive Review of the Antibacterial, Antifungal and Antiviral Potential of Essential Oils and Their Chemical Constituents against Drug-Resistant Microbial Pathogens. Microb. Pathog. 2019, 134, 103580. [Google Scholar] [CrossRef]
- Raut, J.S.; Shinde, R.B.; Chauhan, N.M.; Mohan Karuppayil, S. Terpenoids of Plant Origin Inhibit Morphogenesis, Adhesion, and Biofilm Formation by Candida Albicans. Biofouling 2013, 29, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Neto, N.A.S.; Oliveira, J.T.A.; Aguiar, T.K.B.; Bezerra, L.P.; Branco, L.A.C.; Mesquita, F.P.; Freitas, C.D.T.; Souza, P.F.N. Synergistic Antibiofilm Activity between Synthetic Peptides and Ciprofloxacin against Staphylococcus Aureus. Pathogens 2022, 11, 995. [Google Scholar] [CrossRef] [PubMed]
- An, P.; Yang, X.; Yu, J.; Qi, J.; Ren, X.; Kong, Q. α-Terpineol and Terpene-4-Ol, the Critical Components of Tea Tree Oil, Exert Antifungal Activities in Vitro and in Vivo against Aspergillus Niger in Grapes by Inducing Morphous Damage and Metabolic Changes of Fungus. Food Control 2019, 98, 42–53. [Google Scholar] [CrossRef]
- Etxaniz, A.; González-Bullón, D.; Martín, C.; Ostolaza, H. Membrane Repair Mechanisms against Permeabilization by Pore-Forming Toxins. Toxins 2018, 10, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Billerbeck, V.G.; Roques, C.G.; Bessière, J.-M.; Fonvieille, J.-L.; Dargent, R. Effects of Cymbopogon nardus (L.) W. Watson Essential Oil on the Growth and Morphogenesis of Aspergillus Niger. Can. J. Microbiol. 2001, 47, 9–17. [Google Scholar] [CrossRef]
- Moo, C.L.; Yang, S.K.; Osman, M.A.; Yuswan, M.H.; Loh, J.Y.; Lim, W.M.; Lim, S.H.E.; Lai, K.S. Antibacterial Activity and Mode of Action of β-Caryophyllene on Bacillus Cereus. Polish J. Microbiol. 2020, 69, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Andrade, C.R.D.; Bel, O.; Amaral, L.; Neto, N.A.S.; Silva, R.G.G. Combined Antibiofilm Activity of Synthetic Peptides and Antifungal Drugs against Candida spp. Future Microbiol. 2022, 17, 1133–1146. [Google Scholar] [CrossRef]
- Bezerra, L.P.; Freitas, C.D.T.; Silva, A.F.B.; Amaral, J.L.; Neto, N.A.S.; Silva, R.G.G.; Parra, A.L.C.; Goldman, G.H.; Oliveira, J.T.A.; Mesquita, F.P.; et al. Synergistic Antifungal Activity of Synthetic Peptides and Antifungal Drugs against Candida Albicans and C. Parapsilosis Biofilms. Antibiotics 2022, 11, 553. [Google Scholar] [CrossRef]
- Ding, Y.; Zhang, K.; Yin, Y.; Wu, J. D319 Induced Antifungal Effects through ROS-Mediated Apoptosis and Inhibited Isocitrate Lyase in Candida Albicans. Biochim. Biophys. Acta Gen. Subj. 2022, 1866, 130050. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Shao, Y.-L.; Tang, Y.-J.; Zhou, W.-W. Antifungal Activity of Essential Oil Compounds (Geraniol and Citral) and Inhibitory Mechanisms on Grain Pathogens (Aspergillus flavus and Aspergillus ochraceus). Molecules 2018, 23, 2108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, L.; Chen, S.; Xia, W.; Li, T.; Dong, L.; Zhang, Y.; Zhang, Y.; Li, H.; Zhao, Y.; Fu, X. A New Strategy: Inhibitory Effect and Mechanism of Cymbopogon Martinii Essential Oil on Aspergillus Flavus. Food Biosci. 2023, 51, 102315. [Google Scholar] [CrossRef]
- Almshawit, H.; Macreadie, I. Fungicidal Effect of Thymoquinone Involves Generation of Oxidative Stress in Candida Glabrata. Microbiol. Res. 2017, 195, 81–88. [Google Scholar] [CrossRef]
- Čáp, M.; Váchová, L.; Palková, Z. Reactive Oxygen Species in the Signaling and Adaptation of Multicellular Microbial Communities. Oxid. Med. Cell. Longev. 2012, 2012, 976753. [Google Scholar] [CrossRef] [Green Version]
- Perrone, G.G.; Tan, S.X.; Dawes, I.W. Reactive Oxygen Species and Yeast Apoptosis. Biochim. Biophys. Acta Mol. Cell Res. 2008, 1783, 1354–1368. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.; Zhou, M.; Zuo, Z. Toxic Mechanism of Eucalyptol and β-Cyclocitral on Chlamydomonas Reinhardtii by Inducing Programmed Cell Death. J. Hazard. Mater. 2020, 389, 121910. [Google Scholar] [CrossRef]
- Janssens, S.; Tinel, A. The PIDDosome, DNA-Damage-Induced Apoptosis and Beyond. Cell Death Differ. 2012, 19, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Thakre, A.; Zore, G.; Kodgire, S.; Kazi, R.; Mulange, S.; Patil, R.; Shelar, A.; Santhakumari, B.; Kulkarni, M.; Kharat, K.; et al. Limonene Inhibits Candida Albicans Growth by Inducing Apoptosis. Med. Mycol. 2018, 56, 565–578. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CpEO (50 μg mL−1) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Anti-Candidal Activity | ||||||||
| Microorganisms | Inhibition of Planktonic Cells Growth (%) | Significance a | Inhibition of Biofilm Formation (%) | Significance a | Biomass Reduction of Preformed Biofilm (%) | Significance a | Itraconazole | Significance a |
| C. albicans | 00.00 | p > 0.05 | 00.00 | p > 0.05 | 00.00 | p > 0.05 | 00.00 | p > 0.05 |
| C. krusei | 89.3 ± 0.02 | p < 0.05 | 83.5 ± 0.01 | p < 0.05 | 00.00 | p > 0.05 | 74.5 ± 0.05 | p < 0.05 |
| C. parapsilosis | 80.70 ± 0.03 | p < 0.05 | 77.9 ± 0.07 | p < 0.05 | 00.00 | p > 0.05 | 80.0 ± 0.03 | p < 0.05 |
| Antibacterial Activity | Ciprofloxacin | |||||||
| B. subtilis | 00.00 | p > 0.05 | 00.00 | p > 0.05 | 00.00 | p > 0.05 | 67.5 ± 0.06 | p < 0.05 |
| E. aerogenes | 00.00 | p > 0.05 | 00.00 | p > 0.05 | 00.00 | p > 0.05 | 78.7 ± 0.02 | p < 0.05 |
| E. coli | 00.00 | p > 0.05 | 00.00 | p > 0.05 | 00.00 | p > 0.05 | 68.5 ± 0.007 | p < 0.05 |
| K. pneumoniae | 8.76 ± 0.09 | p > 0.05 | 00.00 | p > 0.05 | 00.00 | p > 0.05 | 72.8 ± 0.06 | p < 0.05 |
| P. aeruginosa | 14.92 ± 0.04 | p < 0.05 | 00.00 | p > 0.05 | 00.00 | p > 0.05 | 77.3 ± 0.07 | p < 0.05 |
| S. aureus | 14.30 ± 0.01 | p < 0.05 | 00.00 | p > 0.05 | 00.00 | p > 0.05 | 87.2 ± 0.07 | p < 0.05 |
| S. enterica | 6.07 ± 0.01 | p > 0.05 | 00.00 | p > 0.05 | 00.00 | p > 0.05 | 62.3 ± 0.01 | p < 0.05 |
| Blood Type | % of Hemolysis | ||||
|---|---|---|---|---|---|
| 0.1 % Triton | 5% DMSO | CpOE 1 mg mL−1 | CpOE 1.25 mg mL−1 | CpOE 3 mg mL−1 | |
| A | 100 | 0 | 0 | 0 | 0 |
| B | 100 | 0 | 0 | 0 | 0 |
| O | 100 | 0 | 0 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carvalho, R.J.P.; Souza, P.F.N.; Malveira, E.A.; Neto, N.A.S.; Silva, R.R.S.; Melo, G.L.C.; Silva, A.F.B.; Lima, L.B.; de Albuquerque, C.C.; Bastos, R.W.; et al. Antimicrobial Activity the Essential Oil from Croton pluriglandulosus Carn. Leaves against Microorganisms of Clinical Interest. J. Fungi 2023, 9, 756. https://doi.org/10.3390/jof9070756
Carvalho RJP, Souza PFN, Malveira EA, Neto NAS, Silva RRS, Melo GLC, Silva AFB, Lima LB, de Albuquerque CC, Bastos RW, et al. Antimicrobial Activity the Essential Oil from Croton pluriglandulosus Carn. Leaves against Microorganisms of Clinical Interest. Journal of Fungi. 2023; 9(7):756. https://doi.org/10.3390/jof9070756
Chicago/Turabian StyleCarvalho, Rayara J. P., Pedro F. N. Souza, Ellen A. Malveira, Nilton A. S. Neto, Romério R. S. Silva, Gabriel L. C. Melo, Ayrles F. B. Silva, Leandro B. Lima, Cynthia C. de Albuquerque, Rafael W. Bastos, and et al. 2023. "Antimicrobial Activity the Essential Oil from Croton pluriglandulosus Carn. Leaves against Microorganisms of Clinical Interest" Journal of Fungi 9, no. 7: 756. https://doi.org/10.3390/jof9070756
APA StyleCarvalho, R. J. P., Souza, P. F. N., Malveira, E. A., Neto, N. A. S., Silva, R. R. S., Melo, G. L. C., Silva, A. F. B., Lima, L. B., de Albuquerque, C. C., Bastos, R. W., Goldman, G. H., & de Freitas, C. D. T. (2023). Antimicrobial Activity the Essential Oil from Croton pluriglandulosus Carn. Leaves against Microorganisms of Clinical Interest. Journal of Fungi, 9(7), 756. https://doi.org/10.3390/jof9070756