
Local Competition and Enhanced Defense: How Metarhizium brunneum Inhibits Verticillium longisporum in Oilseed Rape Plants



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study System
2.2. In Vitro Confrontation Assay
2.3. In Planta Co-Inoculation Assay
- Control: roots mock-inoculated with 0.1% Tween 80.
- Mb: roots inoculated with M. brunneum Cb15III.
- Vl: roots inoculated with V. longisporum VL 43.
- Mb/Vl: roots inoculated with a mix of both fungi.
2.4. Split-Root Assay
- Control: C1 = mock inoculation, C2 = untreated soil.
- Mb: C1 = M. brunneum (Mb-L), C2 = mock inoculation (Mb-S).
- VL: C1 = V. longisporum (Vl-L), C2 = mock inoculation (Vl-S).
- Local: C1 = both fungi present (Mb-L/Vl-L) C2 = both fungi absent (Mb-S/Vl-S).
- Systemic: C1 = M. brunneum present, V. longisporum absent (Mb-L/Vl-S), C2 = V. longisporum present, M. brunneum absent (VL-L/Mb-S).
2.5. Disease Assessment
- 1 = no symptoms.
- 2 = slight symptoms on oldest leaves (yellowing, black veins).
- 3 = slight symptoms on next younger leaves.
- 4 = about 50% of leaves showing symptoms.
- 5 = more than 50% of leaves showing symptoms.
- 6 = up to 50% of leaves dead.
- 7 = more than 50% of leaves dead.
- 8 = only apical meristem still alive.
- 9 = plant dead.
2.6. Quantification of Rhizospheric and Endophytic Metarhizium brunneum
2.7. Gene Expression in Plant Roots
2.8. Data Analysis
3. Results
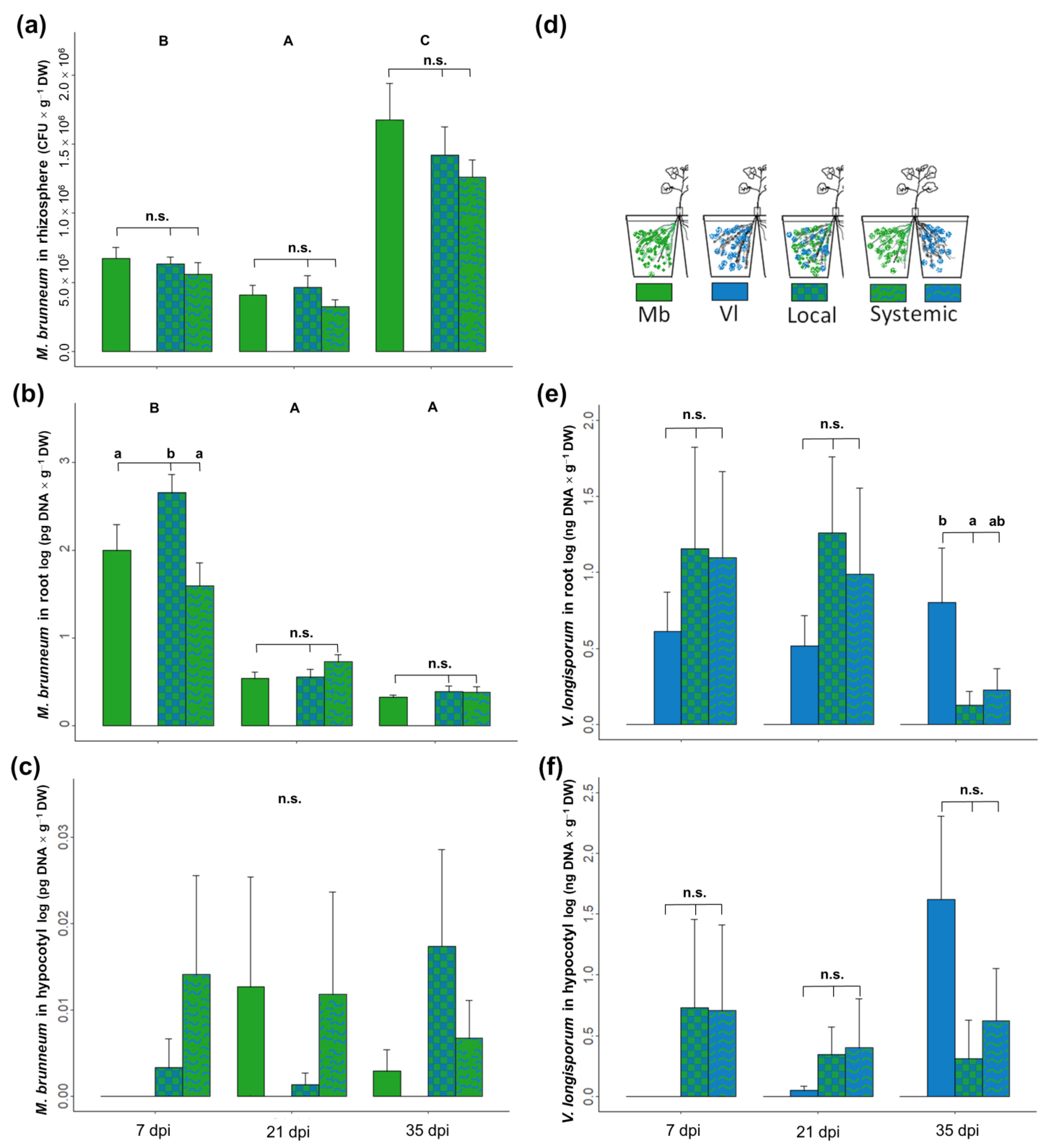
3.1. Direct and Systemic Effects of M. brunneum on V. longisporum Plant Colonization and Disease Development
3.2. Effect of Co-Inoculation M. brunneum on V. longisporum Disease Development
3.3. Direct and Systemic Effects of M. brunneum on V. longisporum Plant Colonization and Disease Development
3.4. Direct and Systemic Plant Responses to V. longisporum and M. brunneum: Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Torsvik, V.; Øvreås, L. Microbial Diversity and Function in Soil: From Genes to Ecosystems. Curr. Opin. Microbiol. 2002, 5, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Bakker, P.A.H.M.; Berendsen, R.L.; Doornbos, R.F.; Wintermans, P.C.A.; Pieterse, C.M.J. The Rhizosphere Revisited: Root Microbiomics. Front. Plant Sci. 2013, 4, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, A.; Schmid, M.; van Tuinen, D.; Berg, G. Plant-Driven Selection of Microbes. Plant Soil 2009, 321, 235–257. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced Systemic Resistance by Beneficial Microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The Rhizosphere Microbiome and Plant Health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Bruck, D.J. Ecology of Metarhizium anisopliae in Soilless Potting Media and the Rhizosphere: Implications for Pest Management. Biol. Control 2005, 32, 155–163. [Google Scholar] [CrossRef]
- Hu, S.; Bidochka, M.J. Root Colonization by Endophytic Insect-Pathogenic Fungi. J. Appl. Microbiol. 2021, 130, 570–581. [Google Scholar] [CrossRef]
- Sasan, R.K.; Bidochka, M.J. The Insect-Pathogenic Fungus Metarhizium robertsii (Clavicipitaceae) Is Also an Endophyte That Stimulates Plant Root Development. Am. J. Bot. 2012, 99, 101–107. [Google Scholar] [CrossRef]
- Behie, S.W.; Jones, S.J.; Bidochka, M.J. Plant Tissue Localization of the Endophytic Insect Pathogenic Fungi Metarhizium and Beauveria. Fungal Ecol. 2015, 13, 112–119. [Google Scholar] [CrossRef]
- Khan, A.L.; Hamayun, M.; Khan, S.A.; Kang, S.M.; Shinwari, Z.K.; Kamran, M.; ur Rehman, S.; Kim, J.G.; Lee, I.J. Pure Culture of Metarhizium anisopliae LHL07 Reprograms Soybean to Higher Growth and Mitigates Salt Stress. World J. Microbiol. Biotechnol. 2012, 28, 1483–1494. [Google Scholar] [CrossRef]
- Verma, P.; Khan, S.A.; Mathur, A.K.; Shanker, K.; Kalra, A. Fungal Endophytes Enhanced the Growth and Production Kinetics of Vinca minor Hairy Roots and Cell Suspensions Grown in Bioreactor. Plant Cell Tissue Organ Cult. 2014, 118, 257–268. [Google Scholar] [CrossRef]
- Ahmad, I.; del Mar Jiménez-Gasco, M.; Luthe, D.S.; Shakeel, S.N.; Barbercheck, M.E. Endophytic Metarhizium robertsii Promotes Maize Growth, Suppresses Insect Growth, and Alters Plant Defense Gene Expression. Biol. Control 2020, 144, 104167. [Google Scholar] [CrossRef]
- Jaber, L.R. Seed Inoculation with Endophytic Fungal Entomopathogens Promotes Plant Growth and Reduces Crown and Root Rot (CRR) Caused by Fusarium culmorum in Wheat. Planta 2018, 248, 1525–1535. [Google Scholar] [CrossRef]
- Jaber, L.R.; Enkerli, J. Fungal Entomopathogens as Endophytes: Can They Promote Plant Growth? Biocontrol Sci. Technol. 2017, 27, 28–41. [Google Scholar] [CrossRef]
- Sanchez-Rodriguez, A.R.; Barron, V.; Del Campillo, M.C.; Quesada-Moraga, E. The Entomopathogenic Fungus Metarhizium brunneum: A Tool for Alleviating Fe Chlorosis. Plant Soil 2016, 406, 295–310. [Google Scholar] [CrossRef]
- Behie, S.W.; Zelisko, P.M.; Bidochka, M.J. Endophytic Insect-Parasitic Fungi Translocate Nitrogen Directly from Insects to Plants. Science 2012, 336, 1576–1577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krell, V.; Unger, S.; Jakobs-Schoenwandt, D.; Patel, A.V. Importance of Phosphorus Supply through Endophytic Metarhizium brunneum for Root:Shoot Allocation and Root Architecture in Potato Plants. Plant Soil 2018, 430, 87–97. [Google Scholar] [CrossRef]
- Gange, A.C.; Koricheva, J.; Currie, A.F.; Jaber, L.R.; Vidal, S. Meta-Analysis of the Role of Entomopathogenic and Unspecialized Fungal Endophytes as Plant Bodyguards. New Phytol. 2019, 223, 2002–2010. [Google Scholar] [CrossRef] [Green Version]
- Vega, F.E. The Use of Fungal Entomopathogens as Endophytes in Biological Control: A Review. Mycologia 2018, 110, 4–30. [Google Scholar] [CrossRef]
- Sasan, R.K.; Bidochka, M.J. Antagonism of the Endophytic Insect Pathogenic Fungus Metarhizium robertsii against the Bean Plant Pathogen Fusarium solani f. Sp. Phaseoli. Can. J. Plant Pathol. 2013, 35, 288–293. [Google Scholar] [CrossRef]
- Miranda-Fuentes, P.; García-Carneros, A.B.; Montilla-Carmona, A.M.; Molinero-Ruiz, L. Evidence of Soil-located Competition as the Cause of the Reduction of Sunflower Verticillium Wilt by Entomopathogenic Fungi. Plant Pathol. 2020, 69, 1492–1503. [Google Scholar] [CrossRef]
- Jaber, L.R.; Alananbeh, K.M. Fungal Entomopathogens as Endophytes Reduce Several Species of Fusarium Causing Crown and Root Rot in Sweet Pepper (Capsicum annuum L.). Biol. Control 2018, 126, 117–126. [Google Scholar] [CrossRef]
- Rivas-Franco, F.; Hampton, J.G.; Morán-Diez, M.E.; Narciso, J.; Rostás, M.; Wessman, P.; Jackson, T.A.; Glare, T.R. Effect of Coating Maize Seed with Entomopathogenic Fungi on Plant Growth and Resistance against Fusarium graminearum and Costelytra giveni. Biocontrol Sci. Technol. 2019, 29, 877–900. [Google Scholar] [CrossRef]
- Gupta, R.; Keppanan, R.; Leibman-Markus, M.; Rav-David, D.; Elad, Y.; Ment, D.; Bar, M. The Entomopathogenic Fungi Metarhizium brunneum and Beauveria bassiana Promote Systemic Immunity and Confer Resistance to a Broad Range of Pests and Pathogens in Tomato. Phytopathology 2022, 112, 784–793. [Google Scholar] [CrossRef] [PubMed]
- St. Leger, R.J.; Wang, J.B. Metarhizium: Jack of All Trades, Master of Many. Open Biol. 2020, 10, 200307. [Google Scholar] [CrossRef]
- Ownley, B.H.; Gwinn, K.D.; Vega, F.E. Endophytic Fungal Entomopathogens with Activity against Plant Pathogens: Ecology and Evolution. BioControl 2010, 55, 113–128. [Google Scholar] [CrossRef]
- Moonjely, S.; Barelli, L.; Bidochka, M.J. Insect Pathogenic Fungi as Endophytes. Adv. Genet. 2016, 94, 107–135. [Google Scholar] [CrossRef]
- Jaber, L.R.; Ownley, B.H. Can We Use Entomopathogenic Fungi as Endophytes for Dual Biological Control of Insect Pests and Plant Pathogens? Biol. Control 2018, 116, 36–45. [Google Scholar] [CrossRef]
- Dreischhoff, S.; Das, I.S.; Jakobi, M.; Kasper, K.; Polle, A. Local Responses and Systemic Induced Resistance Mediated by Ectomycorrhizal Fungi. Front. Plant Sci. 2020, 11, 590063. [Google Scholar] [CrossRef]
- Hill, E.M.; Robinson, L.A.; Abdul-sada, A.; Vanbergen, A.J.; Hodge, A.; Hartley, S.E.; Hartley, S.E. Arbuscular Mycorrhizal Fungi and Plant Chemical Defence: Effects of Colonisation on Aboveground and Belowground Metabolomes. J. Chem. Ecol. 2018, 44, 198–208. [Google Scholar] [CrossRef] [Green Version]
- Shoresh, M.; Harman, G.E.; Mastouri, F. Induced Systemic Resistance and Plant Responses to Fungal Biocontrol Agents. Annu. Rev. Phytopathol. 2010, 48, 21–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, K.; Wang, F.; Nong, X.; McNeill, M.R.; Liu, S.; Wang, G.; Cao, G.; Zhang, Z. Response of Peanut Arachis hypogaea Roots to the Presence of Beneficial and Pathogenic Fungi by Transcriptome Analysis. Sci. Rep. 2017, 7, 964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raad, M.; Glare, T.R.; Brochero, H.L.; Müller, C.; Rostás, M. Transcriptional Reprogramming of Arabidopsis thaliana Defence Pathways by the Entomopathogen Beauveria bassiana Correlates with Resistance Against a Fungal Pathogen but Not Against Insects. Front. Microbiol. 2019, 10, 615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, X.; Zhao, X.; Huang, S.; Deng, J.; Li, X.; Luo, Z.; Zhang, Y. Pest Management via Endophytic Colonization of Tobacco Seedlings by the Insect Fungal Pathogen Beauveria bassiana. Pest Manag. Sci. 2021, 77, 2007–2018. [Google Scholar] [CrossRef]
- Cotes, B.; Thöming, G.; Amaya-Gómez, C.V.; Novák, O.; Nansen, C. Root-Associated Entomopathogenic Fungi Manipulate Host Plants to Attract Herbivorous Insects. Sci. Rep. 2020, 10, 22424. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Franco, F.; Hampton, J.G.; Narciso, J.; Rostás, M.; Wessman, P.; Saville, D.J.; Jackson, T.A.; Glare, T.R. Effects of a Maize Root Pest and Fungal Pathogen on Entomopathogenic Fungal Rhizosphere Colonization, Endophytism and Induction of Plant Hormones. Biol. Control 2020, 150, 104347. [Google Scholar] [CrossRef]
- Posada-Vergara, C.; Lohaus, K.; Alhussein, M.; Vidal, S.; Rostás, M. Root Colonization by Fungal Entomopathogen Systemically Primes Belowground Plant Defense against Cabbage Root Fly. J. Fungi 2022, 8, 969. [Google Scholar] [CrossRef]
- Cachapa, J.C.; Meyling, N.V.; Burow, M.; Hauser, T.P. Induction and Priming of Plant Defense by Root-Associated Insect-Pathogenic Fungi. J. Chem. Ecol. 2020, 47, 112–122. [Google Scholar] [CrossRef]
- Qing, Y.; Ourry, M.; Burow, M.; Meyling, N.V.; Hauser, T.P. Defense Priming in Cabbage (Brassica oleracea) by Insect-Pathogenic Fungi. Arthropod Plant Interact. 2023, 17, 275–287. [Google Scholar] [CrossRef]
- Depotter, J.R.L.L.; Deketelaere, S.; Inderbitzin, P.; Von Tiedemann, A.; Höfte, M.; Subbarao, K.V.; Wood, T.A.; Thomma, B.P.H.J. Verticillium longisporum, the Invisible Threat to Oilseed Rape and Other Brassicaceous Plant Hosts. Mol. Plant Pathol. 2016, 17, 1004–1016. [Google Scholar] [CrossRef] [Green Version]
- Eynck, C.; Koopmann, B.; Grunewaldt-Stoecker, G.; Karlovsky, P.; von Tiedemann, A. Differential Interactions of Verticillium longisporum and V. Dahliae with Brassica napus Detected with Molecular and Histological Techniques. Eur. J. Plant Pathol. 2007, 118, 259–274. [Google Scholar] [CrossRef]
- Eynck, C.; Koopmann, B.; Karlovsky, P.; Von Tiedemann, A. Internal Resistance in Winter Oilseed Rape Inhibits Systemic Spread of the Vascular Pathogen Verticillium longisporum. Phytopathology 2009, 99, 802–811. [Google Scholar] [CrossRef] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madden, L.V.; Hughes, G.; Bosch, F. The Study of Plant Disease Epidemics; The American Phytopathological Society: St. Paul, MI, USA, 2007. [Google Scholar]
- Strasser, H.; Forer, A.; Schinner, F. Development of Media for the Selective Isolation and Maintenance of Virulence of Beauveria brongniartii. In Proceedings of the 3rd International Workshop on Microbial Control of Soil Dwelling Pests, Lincoln, New Zealand, 21–23 February 1996; Jackson, T.A., Glare, T.A., Eds.; AgResearch Lincoln: Lincoln, New Zealand, 1996; pp. 125–130. [Google Scholar]
- Brandfass, C.; Karlovsky, P. Simultaneous Detection of Fusarium culmorum and F. graminearum in Plant Material by Duplex PCR with Melting Curve Analysis. BMC Microbiol. 2006, 6, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, S.; Rehner, S.A.; Widmer, F.; Enkerli, J. A PCR-Based Tool for Cultivation-Independent Detection and Quantification of Metarhizium Clade 1. J. Invertebr. Pathol. 2011, 108, 106–114. [Google Scholar] [CrossRef]
- Behrens, F.H.; Schenke, D.; Hossain, R.; Ye, W.; Schemmel, M.; Bergmann, T.; Häder, C.; Zhao, Y.; Ladewig, L.; Zhu, W.; et al. Suppression of Abscisic Acid Biosynthesis at the Early Infection Stage of Verticillium longisporum in Oilseed Rape (Brassica Napus). Mol. Plant Pathol. 2019, 20, 1645–1661. [Google Scholar] [CrossRef]
- Zheng, X.; Koopmann, B.; von Tiedemann, A. Role of Salicylic Acid and Components of the Phenylpropanoid Pathway in Basal and Cultivar-Related Resistance of Oilseed Rape (Brassica napus) to Verticillium longisporum. Plants 2019, 8, 491. [Google Scholar] [CrossRef] [Green Version]
- Koressaar, T.; Lepamets, M.; Kaplinski, L.; Raime, K.; Andreson, R.; Remm, M. Primer3_masker: Integrating Masking of Template Sequence with Primer Design Software. Bioinformatics 2018, 34, 1937–1938. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Mendiburu, F.; Yaseen, M. Agricolae: Statistical Procedures for Agricultural Research 2020. R Package Version 1.4.0. Available online: https://myaseen208.github.io/agricolae/https://cran.r-project.org/package=agricolae (accessed on 27 May 2021).
- Lozano-Tovar, M.D.; Ortiz-Urquiza, A.; Garrido-Jurado, I.; Trapero-Casas, A.; Quesada-Moraga, E. Assessment of Entomopathogenic Fungi and Their Extracts against a Soil-Dwelling Pest and Soil-Borne Pathogens of Olive. Biol. Control 2013, 67, 409–420. [Google Scholar] [CrossRef]
- Guigón López, C.; Muñoz Castellanos, L.N.; Flores Ortiz, N.A.; González González, J.A. Control of Powdery Mildew (Leveillula taurica) Using Trichoderma asperellum and Metarhizium anisopliae in Different Pepper Types. BioControl 2019, 64, 77–89. [Google Scholar] [CrossRef]
- Lozano-Tovar, M.D.; Garrido-Jurado, I.; Quesada-Moraga, E.; Raya-Ortega, M.C.; Trapero-Casas, A. Metarhizium brunneum and Beauveria bassiana Release Secondary Metabolites with Antagonistic Activity against Verticillium dahliae and Phytophthora megasperma Olive Pathogens. Crop Prot. 2017, 100, 186–195. [Google Scholar] [CrossRef]
- Hummadi, E.H.; Cetin, Y.; Demirbek, M.; Kardar, N.M.; Khan, S.; Coates, C.J.; Eastwood, D.C.; Dudley, E.; Maffeis, T.; Loveridge, J.; et al. Antimicrobial Volatiles of the Insect Pathogen Metarhizium brunneum. J. Fungi 2022, 8, 326. [Google Scholar] [CrossRef] [PubMed]
- Knüfer, J.; Lopisso, D.T.; Koopmann, B.; Karlovsky, P.; von Tiedemann, A. Assessment of Latent Infection with Verticillium longisporum in Field-Grown Oilseed Rape by QPCR. Eur. J. Plant Pathol. 2017, 147, 819–831. [Google Scholar] [CrossRef]
- Tyvaert, L.; França, S.C.; Debode, J.; Höfte, M. The Endophyte Verticillium Vt305 Protects Cauliflower against Verticillium Wilt. J. Appl. Microbiol. 2014, 116, 1563–1571. [Google Scholar] [CrossRef]
- Nakano, T.; Nishiuchi, T.; Suzuki, K.; Fujimura, T.; Shinshi, H. Studies on Transcriptional Regulation of Endogenous Genes by ERF2 Transcription Factor in Tobacco Cells. Plant Cell Physiol. 2006, 47, 554–558. [Google Scholar] [CrossRef] [Green Version]
- Hafiz, F.B.; Moradtalab, N.; Goertz, S.; Rietz, S.; Dietel, K.; Rozhon, W.; Humbeck, K.; Geistlinger, J.; Neumann, G.; Schellenberg, I. Synergistic Effects of a Root-Endophytic Trichoderma Fungus and Bacillus on Early Root Colonization and Defense Activation Against Verticillium longisporum in Rapeseed. Mol. Plant-Microbe Interact. 2022, 35, 380–392. [Google Scholar] [CrossRef]
- Azadi, N.; Shirzad, A.; Mohammadi, H. A Study of Some Biocontrol Mechanisms of Beauveria bassiana against Rhizoctonia disease on Tomato. Acta Biol. Szeged. 2016, 60, 119–127. [Google Scholar]
- Cabanás, C.G.L.; Sesmero, R.; Valverde-Corredor, A.; Javier López-Escudero, F.; Mercado-Blanco, J. A Split-Root System to Assess Biocontrol Effectiveness and Defense-Related Genetic Responses in above-Ground Tissues during the Tripartite Interaction Verticillium dahliae-Olive-Pseudomonas fluorescens PICF7 in Roots. Plant Soil 2017, 417, 433–452. [Google Scholar] [CrossRef]
- Yedidia, I.; Shoresh, M.; Kerem, Z.; Benhamou, N.; Kapulnik, Y.; Chet, I. Concomitant Induction of Systemic Resistance to Pseudomonas syringae Pv. Lachrymans in Cucumber by Trichoderma asperellum (T-203) and Accumulation of Phytoalexins. Appl. Environ. Microbiol. 2003, 69, 7343–7353. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Wang, H.; Xie, J.; Lv, J.; Zhang, G.; Hu, L.; Luo, S.; Li, L.; Yu, J. The Roles of Cruciferae Glucosinolates in Disease and Pest Resistance. Plants 2021, 10, 1097. [Google Scholar] [CrossRef] [PubMed]
- Bednarek, P.; Piślewska-Bednarek, M.; Svatoš, A.; Schneider, B.; Doubský, J.; Mansurova, M.; Humphry, M.; Consonni, C.; Panstruga, R.; Sanchez-Vallet, A.; et al. A Glucosinolate Metabolism Pathway in Living Plant Cells Mediates Broad-Spectrum Antifungal Defense. Science 2009, 323, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Vig, A.P.; Rampal, G.; Thind, T.S.; Arora, S. Bio-Protective Effects of Glucosinolates—A Review. LWT Food Sci. Technol. 2009, 42, 1561–1572. [Google Scholar] [CrossRef]
- Fröschel, C. In-Depth Evaluation of Root Infection Systems Using the Vascular Fungus Verticillium longisporum as Soil-Borne Model Pathogen. Plant Methods 2021, 17, 57. [Google Scholar] [CrossRef] [PubMed]
- Iven, T.; König, S.; Singh, S.; Braus-Stromeyer, S.A.; Bischoff, M.; Tietze, L.F.; Braus, G.H.; Lipka, V.; Feussner, I.; Dröge-Laser, W. Transcriptional Activation and Production of Tryptophan-Derived Secondary Metabolites in Arabidopsis Roots Contributes to the Defense against the Fungal Vascular Pathogen Verticillium longisporum. Mol. Plant 2012, 5, 1389–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nongbri, P.L.; Johnson, J.M.; Sherameti, I.; Glawischnig, E.; Halkier, B.A.; Oelmüller, R.; Botany, G.; Physiology, P.; Jena, F.; Str, D. Indole-3-Acetaldoxime-Derived Compounds Restrict Root Colonization in the Beneficial Interaction Between Arabidopsis Roots and the Endophyte Piriformospora indica. Mol. Plant Microbe Interact. 2012, 25, 1186–1197. [Google Scholar] [CrossRef] [Green Version]
- Harman, G.E.; Petzoldt, R.; Comis, A.; Chen, J. Interactions between Trichoderma harzianum Strain T22 and Maize Inbred Line Mo17 and Effects of These Interactions on Diseases Caused by Pythiuin ultimum and Colletotrichum graminicola. Phytopathology 2004, 94, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Newman, M.A.; Sundelin, T.; Nielsen, J.T.; Erbs, G. MAMP (Microbe-Associated Molecular Pattern) Triggered Immunity in Plants. Front. Plant Sci. 2013, 4, 139. [Google Scholar] [CrossRef] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Posada-Vergara, C.; Vidal, S.; Rostás, M. Local Competition and Enhanced Defense: How Metarhizium brunneum Inhibits Verticillium longisporum in Oilseed Rape Plants. J. Fungi 2023, 9, 796. https://doi.org/10.3390/jof9080796
Posada-Vergara C, Vidal S, Rostás M. Local Competition and Enhanced Defense: How Metarhizium brunneum Inhibits Verticillium longisporum in Oilseed Rape Plants. Journal of Fungi. 2023; 9(8):796. https://doi.org/10.3390/jof9080796
Chicago/Turabian StylePosada-Vergara, Catalina, Stefan Vidal, and Michael Rostás. 2023. "Local Competition and Enhanced Defense: How Metarhizium brunneum Inhibits Verticillium longisporum in Oilseed Rape Plants" Journal of Fungi 9, no. 8: 796. https://doi.org/10.3390/jof9080796
APA StylePosada-Vergara, C., Vidal, S., & Rostás, M. (2023). Local Competition and Enhanced Defense: How Metarhizium brunneum Inhibits Verticillium longisporum in Oilseed Rape Plants. Journal of Fungi, 9(8), 796. https://doi.org/10.3390/jof9080796