
Inventory of the Secondary Metabolite Biosynthetic Potential of Members within the Terminal Clade of the Fusarium solani Species Complex

Abstract
:1. Introduction
2. Materials and Methods
2.1. Genomic Data of FSSC Isolates
2.2. Species Phylogeny of FSSC Isolates
2.3. Prediction of Secondary Metabolite Gene Clusters and Phylogenetic Analysis of PKS and NRPS Proteins
2.4. Similarity Network and Phylogenetic Analysis of the Terpene Synthase/Cyclase, DMATS, and Phosphoenolpyruvate Phosphomutase
2.5. Analysis of the Genetic Organization of the Conserved and Unique Biosynthetic Gene Clusters with Known Products
3. Results
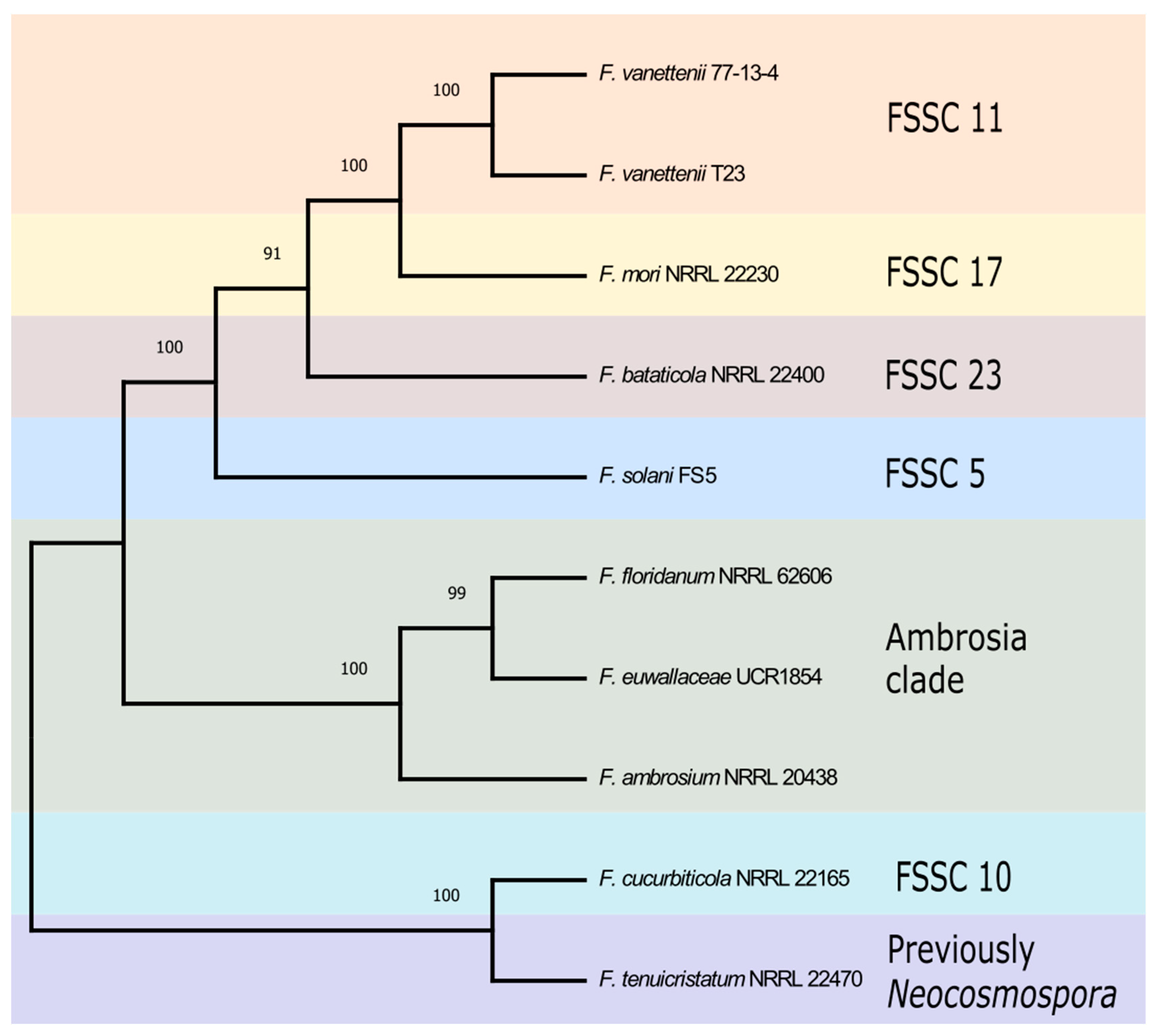
3.1. Phylogenetic Analysis of FSSC Isolates Used in This Study
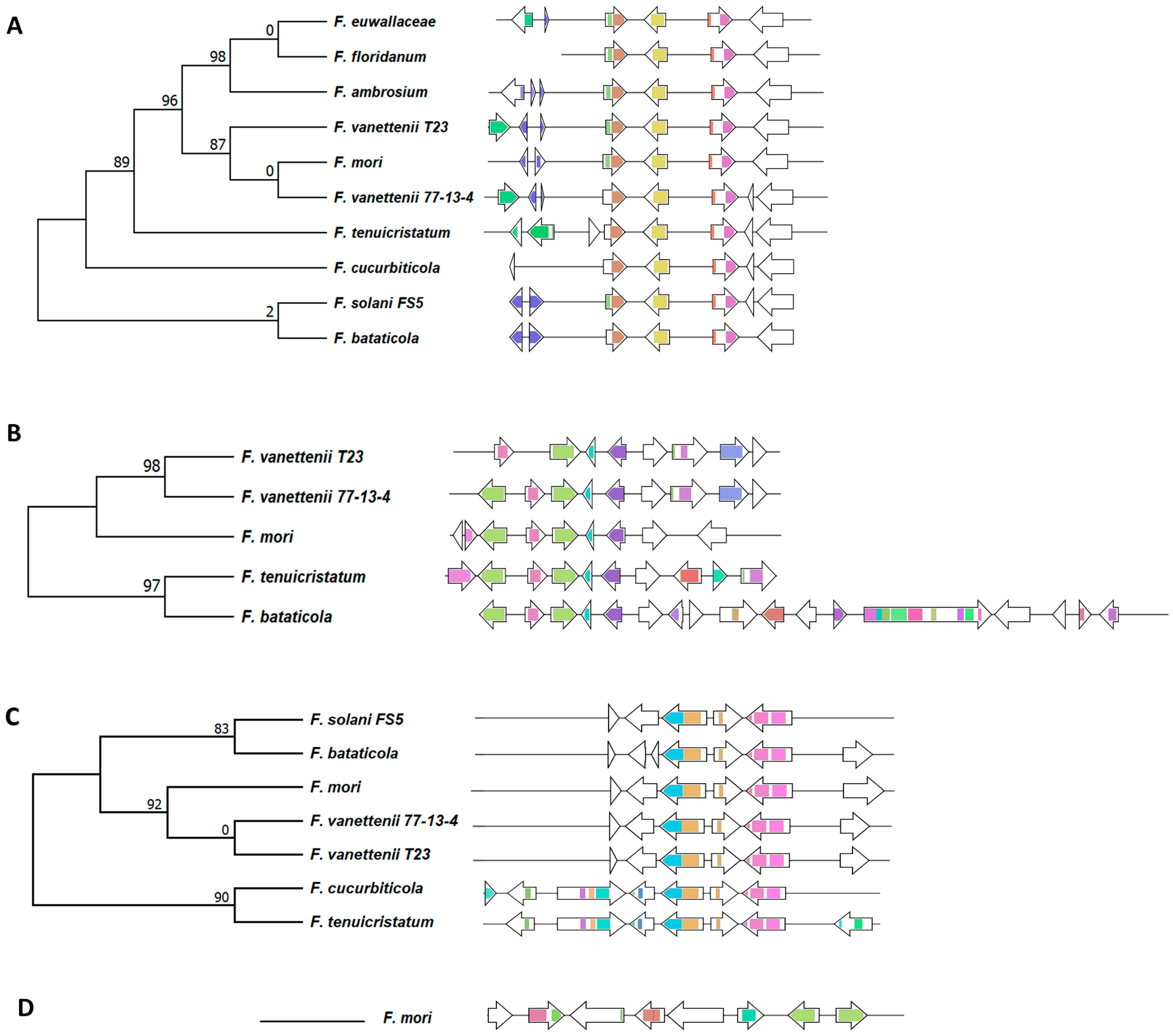
3.2. Identification of Secondary Metabolite Biosynthetic Clusters
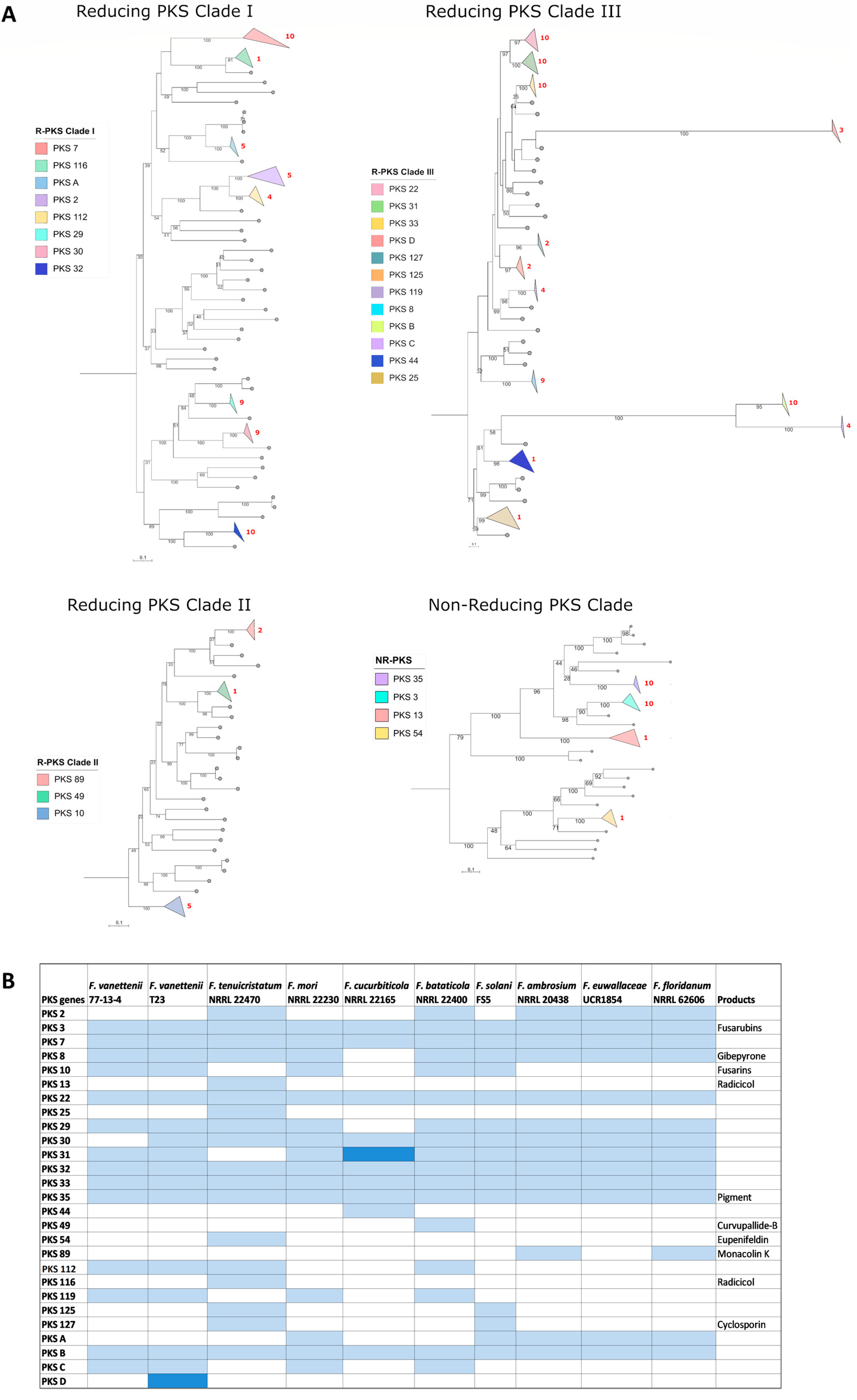
3.3. Comparison of the Polyketide Synthase (PKS)-Containing Clusters
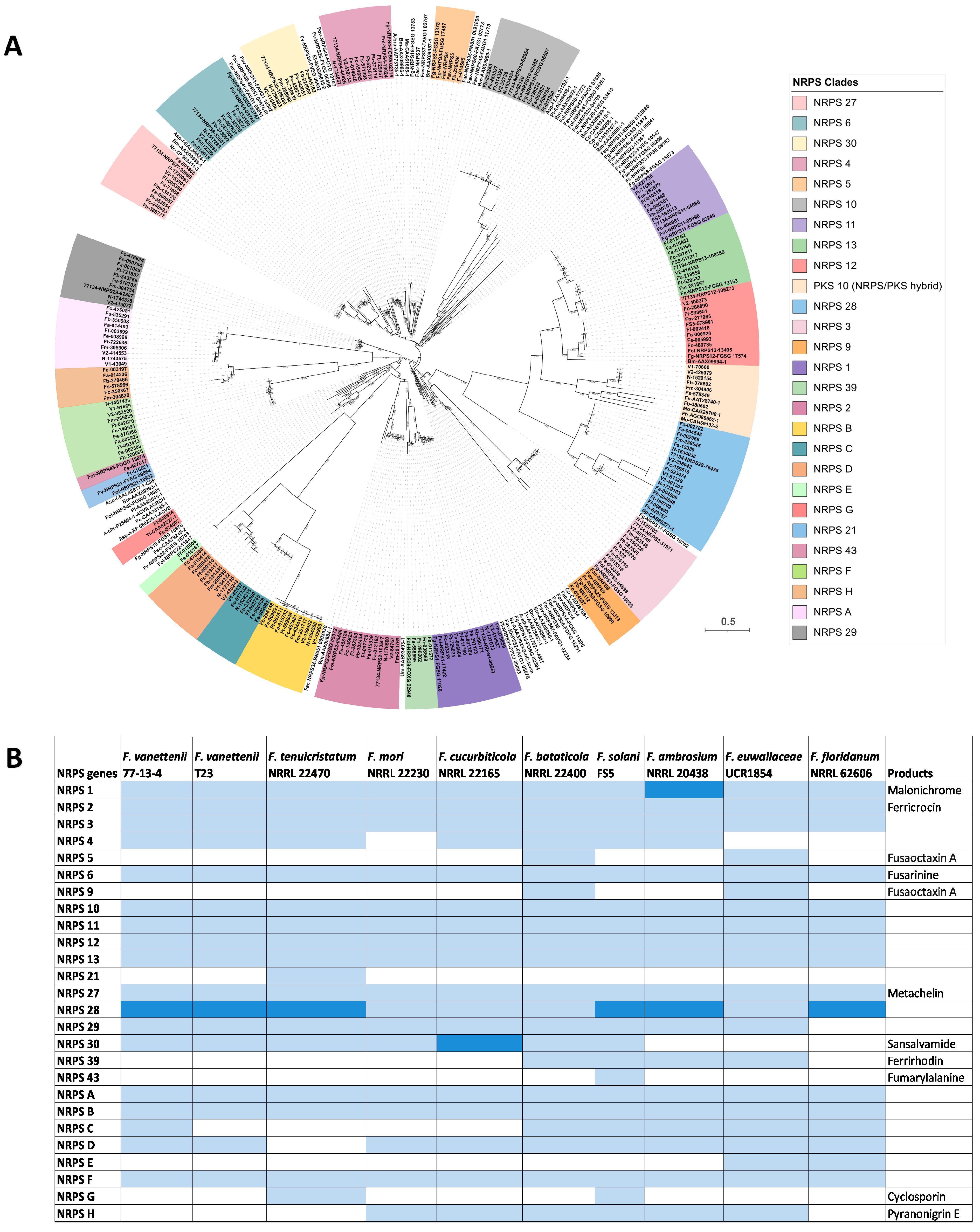
3.4. Comparison of the Nonribosomal Peptide Synthetase (NRPS)-Containing Clusters
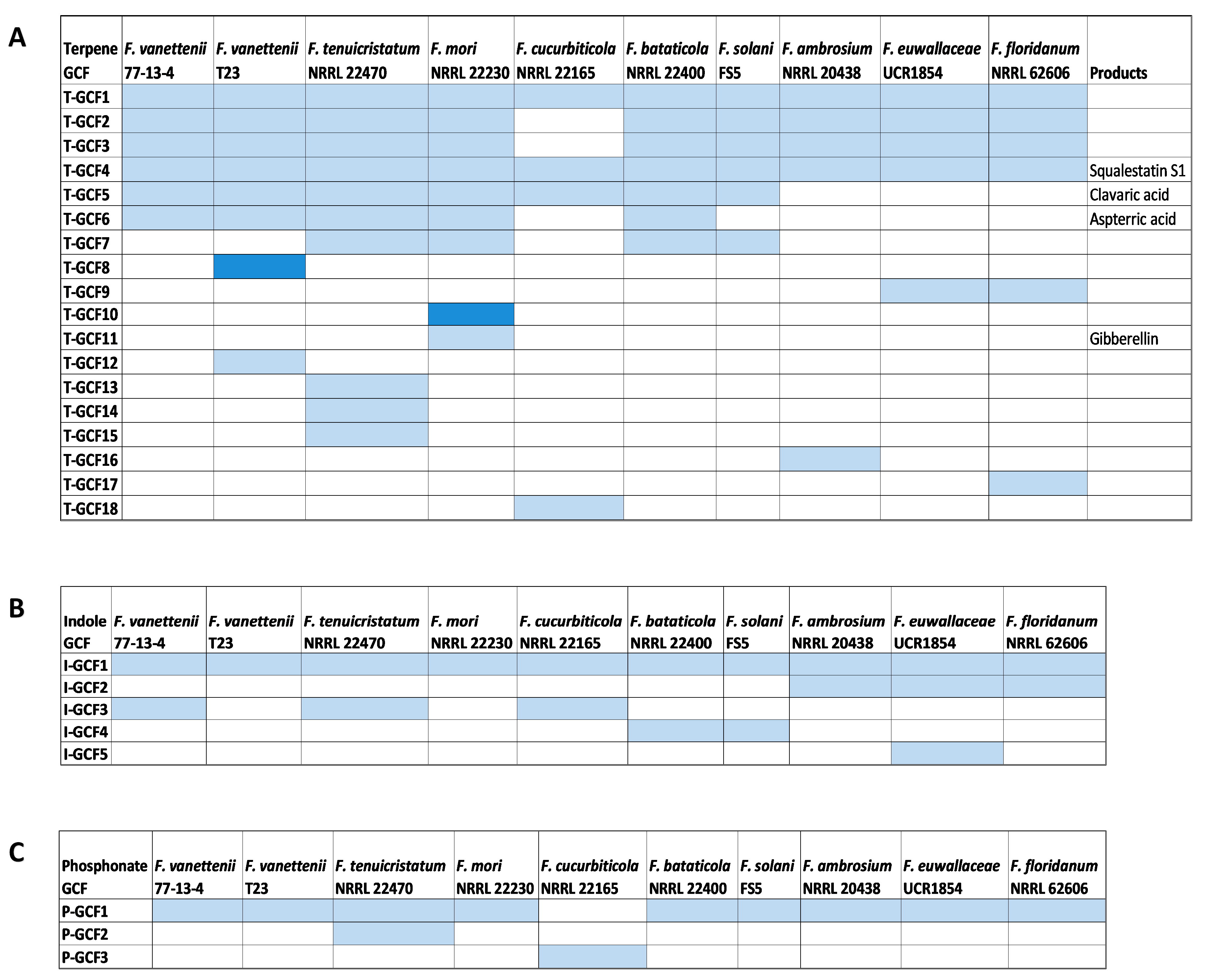
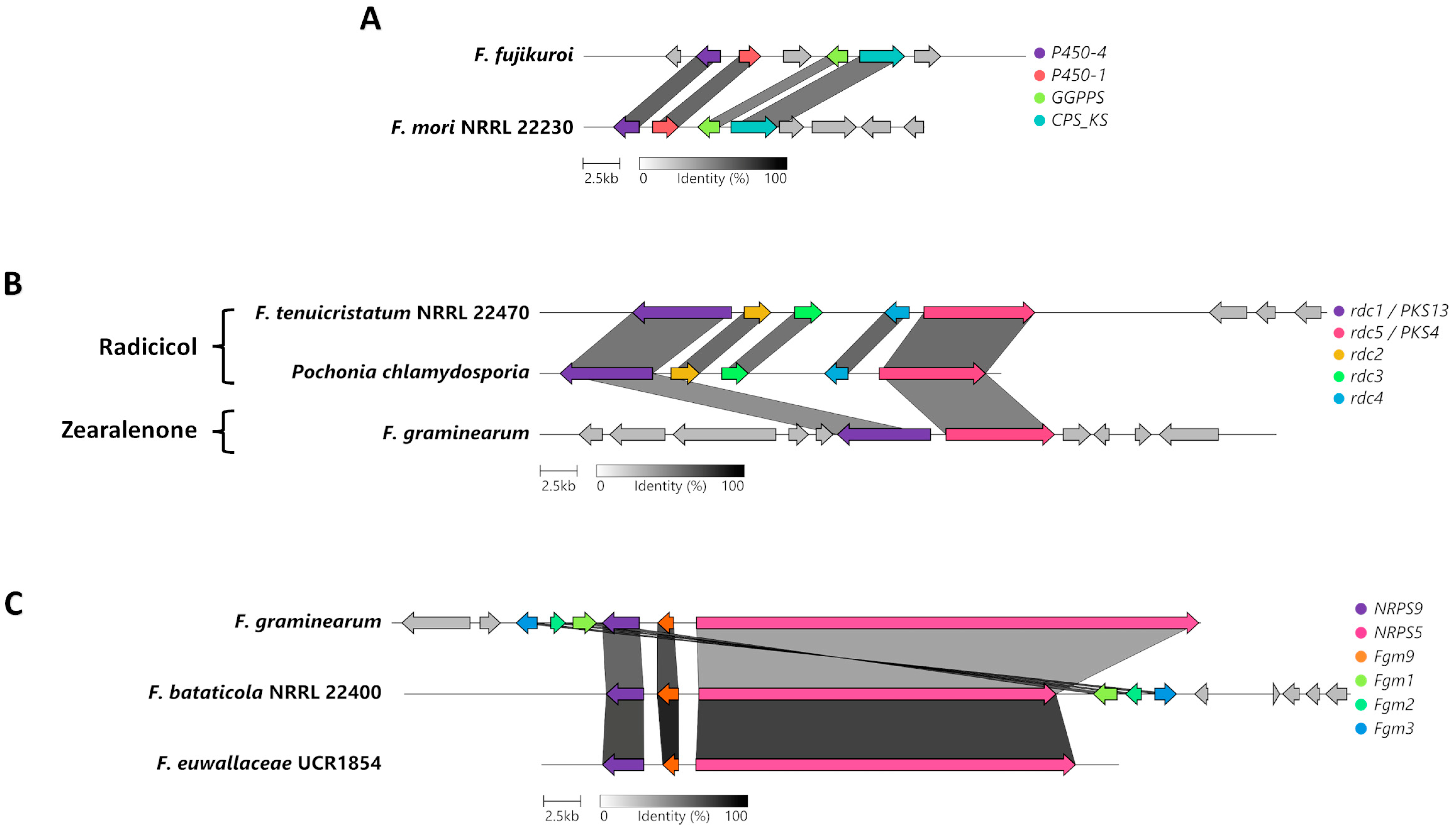
3.5. Comparison of the Terpene Synthase/Cyclase, DMATS, and Phosphonate-Producing BGCs
3.6. Unique and Specific BGCs of Interest
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Keller, N.P. Fungal secondary metabolism: Regulation, function and drug discovery. Nat. Rev. Microbiol. 2019, 17, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Geiser, D.M.; Aoki, T.; Bacon, C.W.; Baker, S.E.; Bhattacharyya, M.K.; Brandt, M.E.; Brown, D.W.; Burgess, L.W.; Chulze, S.; Coleman, J.J.; et al. One fungus, one name: Defining the genus Fusarium in a scientifically robust way that preserves longstanding use. Phytopathology 2013, 103, 400–408. [Google Scholar] [CrossRef] [Green Version]
- Geiser, D.M.; Al-Hatmi, A.; Aoki, T.; Arie, T.; Balmas, V.; Barnes, I.; Bergstrom, G.C.; Bhattacharyya, M.K.K.; Blomquist, C.L.; Bowden, R.; et al. Phylogenomic analysis of a 55.1 kb 19-gene dataset resolves a monophyletic Fusarium that includes the Fusarium solani Species Complex. Phytopathology 2020, 111, 1064–1079. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Rooney, A.P.; Proctor, R.H.; Brown, D.W.; McCormick, S.P.; Ward, T.J.; Frandsen, R.J.N.; Lysoe, E.; Rehner, S.A.; Aoki, T.; et al. Phylogenetic analyses of RPB1 and RPB2 support a middle Cretaceous origin for a clade comprising all agriculturally and medically important fusaria. Fungal Genet. Biol. 2013, 52, 20–31. [Google Scholar] [CrossRef]
- Coleman, J.J. The Fusarium solani species complex: Ubiquitous pathogens of agricultural importance. Mol. Plant Pathol. 2016, 17, 146–158. [Google Scholar] [CrossRef] [PubMed]
- Muhammed, M.; Anagnostou, T.; Desalermos, A.; Kourkoumpetis, T.K.; Carneiro, H.A.; Glavis-Bloom, J.; Coleman, J.J.; Mylonakis, E. Fusarium infection: Report of 26 cases and review of 97 cases from the literature. Medicine 2013, 92, 305–316. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; O’Donnell, K.; Sutton, D.A.; Nalim, F.A.; Summerbell, R.C.; Padhye, A.A.; Geiser, D.M. Members of the Fusarium solani species complex that cause infections in both humans and plants are common in the environment. J. Clin. Microbiol. 2006, 44, 2186–2190. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, K.; Sutton, D.A.; Fothergill, A.; McCarthy, D.; Rinaldi, M.G.; Brandt, M.E.; Zhang, N.; Geiser, D.M. Molecular phylogenetic diversity, multilocus haplotype nomenclature, and in vitro antifungal resistance within the Fusarium solani species complex. J. Clin. Microbiol. 2008, 46, 2477–2490. [Google Scholar] [CrossRef] [Green Version]
- Medentsev, A.G.; Akimenko, V.K. Naphthoquinone metabolites of the fungi. Phytochemistry 1998, 47, 935–959. [Google Scholar] [CrossRef]
- Short, D.P.G.; O’Donnell, K.; Thrane, U.; Nielsen, K.F.; Zhang, N.; Juba, J.H.; Geiser, D.M. Phylogenetic relationships among members of the Fusarium solani species complex in human infections and the descriptions of F. keratoplasticum sp nov and F. petroliphilum stat. nov. Fungal Genet. Biol. 2013, 53, 59–70. [Google Scholar] [CrossRef]
- Nielsen, M.R.; Sondergaard, T.E.; Giese, H.; Sørensen, J.L. Advances in linking polyketides and non-ribosomal peptides to their biosynthetic gene clusters in Fusarium. Curr. Genet. 2019, 65, 1263–1280. [Google Scholar] [CrossRef]
- Romans-Fuertes, P.; Sondergaard, T.E.; Sandmann, M.I.H.; Wollenberg, R.D.; Nielsen, K.F.; Hansen, F.T.; Giese, H.; Brodersen, D.E.; Sørensen, J.L. Identification of the non-ribosomal peptide synthetase responsible for biosynthesis of the potential anti-cancer drug sansalvamide in Fusarium solani. Curr. Genet. 2016, 62, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Graziani, S.; Vasnier, C.; Daboussi, M.J. Novel polyketide synthase from Nectria haematococca. Appl. Environ. Microbiol. 2004, 70, 2984–2988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, D.W.; Kim, H.-S.; McGovern, A.E.; Probyn, C.E.; Proctor, R.H. Genus-wide analysis of Fusarium polyketide synthases reveals broad chemical potential. Fungal Genet. Biol. 2022, 160, 103696. [Google Scholar] [CrossRef] [PubMed]
- Coleman, J.J.; Rounsley, S.D.; Rodriguez-Carres, M.; Kuo, A.; Wasmann, C.C.; Grimwood, J.; Schmutz, J.; Taga, M.; White, G.J.; Zhou, S.; et al. The genome of Nectria haematococca: Contribution of supernumerary chromosomes to gene expansion. PLOS Genet. 2009, 5, e1000618. [Google Scholar] [CrossRef] [Green Version]
- Coleman, J.J. Exploring the Supernumerary Chromosomes of the Fusarium Solani and Fusarium oxysporum Species Complexes: Reservoirs for Function Genetic Diversity; USDDOE Joint Genome Institute (JGI): Walnut Creek, CA, USA, 2013. [Google Scholar]
- Mesny, F.; Miyauchi, S.; Thiergart, T.; Pickel, B.; Atanasova, L.; Karlsson, M.; Hüttel, B.; Barry, K.W.; Haridas, S.; Chen, C.; et al. Genetic determinants of endophytism in the Arabidopsis root mycobiome. Nat. Commun. 2021, 12, 7227. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [Green Version]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema, M.H.; Weber, T. AntiSMASH 6.0: Improving Cluster Detection and Comparison Capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Brown, D.W.; Proctor, R.H. Insights into natural products biosynthesis from analysis of 490 polyketide synthases from Fusarium. Fungal Genet. Biol. 2016, 89, 37–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, F.T.; Gardiner, D.M.; Lysøe, E.; Fuertes, P.R.; Tudzynski, B.; Wiemann, P.; Sondergaard, T.E.; Giese, H.; Brodersen, D.E.; Sørensen, J.L. An Update to Polyketide Synthase and Non-Ribosomal Synthetase Genes and Nomenclature in Fusarium. Fungal Genet. Biol. 2015, 75, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Bushley, K.E.; Turgeon, B.G. Phylogenomics reveals subfamilies of fungal nonribosomal peptide synthetases and their evolutionary relationships. BMC Evol. Biol. 2010, 10, 26. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Criscuolo, A.; Gribaldo, S. BMGE (Block Mapping and Gathering with Entropy): A new software for selection of phylogenetic informative regions from multiple sequence alignments. BMC Evol. Biol. 2010, 10, 210. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. ProtTest 3: Fast selection of best-fit models of protein evolution. Bioinformatics 2011, 27, 1164–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML Version 8: A Tool for Phylogenetic Analysis and Post-Analysis of Large Phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) V5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Navarro-Muñoz, J.C.; Selem-Mojica, N.; Mullowney, M.W.; Kautsar, S.A.; Tryon, J.H.; Parkinson, E.I.; De Los Santos, E.L.C.; Yeong, M.; Cruz-Morales, P.; Abubucker, S.; et al. A computational framework to explore large-scale biosynthetic diversity. Nat. Chem. Biol. 2020, 16, 60–68. [Google Scholar] [CrossRef]
- Terlouw, B.R.; Blin, K.; Navarro-Muñoz, J.C.; Avalon, N.E.; Chevrette, M.G.; Egbert, S.; Lee, S.; Meijer, D.; Recchia, M.J.J.; Reitz, Z.L.; et al. MIBiG 3.0: A community-driven effort to annotate experimentally validated biosynthetic gene clusters. Nucleic Acids Res. 2023, 51, D603–D610. [Google Scholar] [CrossRef]
- Gilchrist, C.M.; Chooi, Y.-H. clinker & clustermap.js: Automatic generation of gene cluster comparison figures. Bioinformatics 2021, 37, 2473–2475. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, Y.; Barr, J.R.; Barr, D.B.; Brock, J.W.; Elie, C.M.; Ueno, Y.; Patterson, D.G.; Potter, M.E.; Reiss, E. Physiological characteristics and mycotoxins of human clinical isolates of Fusarium species. Mycol. Res. 1999, 103, 1462–1468. [Google Scholar] [CrossRef]
- Jia, L.-J.; Tang, H.-Y.; Wang, W.-Q.; Yuan, T.-L.; Wei, W.-Q.; Pang, B.; Gong, X.-M.; Wang, S.-F.; Li, Y.-J.; Zhang, D.; et al. A linear nonribosomal octapeptide from Fusarium graminearum facilitates cell-to-cell invasion in wheat. Nat. Commun. 2019, 10, 922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coleman, J.J.; Ghosh, S.; Okoli, I.; Mylonakis, E. Antifungal activity of microbial secondary metabolites. PLoS ONE 2011, 6, e25321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, H.; Eisendle, M.; Turgeon, B.G. Siderophores in fungal physiology and virulence. Annu. Rev. Phytopathol. 2008, 46, 149–187. [Google Scholar] [CrossRef]
- Oide, S.; Berthiller, F.; Wiesenberger, G.; Adam, G.; Turgeon, B.G. Individual and combined roles of malonichrome, ferricrocin, and TAFC siderophores in Fusarium graminearum pathogenic and sexual development. Front. Microbiol. 2015, 5, 759. [Google Scholar] [CrossRef]
- Lee, B.-N.; Kroken, S.; Chou, D.Y.T.; Robbertse, B.; Yoder, O.C.; Turgeon, B.G. Functional analysis of all nonribosomal peptide synthetases in Cochliobolus heterostrophus reveals a factor, NPS6, involved in virulence and resistance to oxidative stress. Eukaryot. Cell 2005, 4, 545–555. [Google Scholar] [CrossRef] [Green Version]
- Krasnoff, S.B.; Keresztes, I.; Donzelli, B.G.G.; Gibson, D.M. Metachelins, mannosylated and N-oxidized coprogen-type siderophores from Metarhizium robertsii. J. Nat. Prod. 2014, 77, 1685–1692. [Google Scholar] [CrossRef]
- Chang, H.-X.; Domier, L.L.; Radwan, O.; Yendrek, C.R.; Hudson, M.E.; Hartman, G.L. Identification of multiple phytotoxins produced by Fusarium virguliforme including a phytotoxic effector (FvNIS1) associated with sudden death syndrome foliar symptoms. Mol. Plant-Microbe Interact. 2016, 29, 96–108. [Google Scholar] [CrossRef] [Green Version]
- Piper, P.W.; Millson, S.H. Spotlight on the microbes that produce heat shock protein 90-targeting antibiotics. Open Biol. 2012, 2, 120138. [Google Scholar] [CrossRef]
- Wicklow, D.T.; Jordan, A.M.; Gloer, J.B. Antifungal metabolites (monorden, monocillins I, II, III) from Colletotrichum graminicola, a systemic vascular pathogen of maize. Mycol. Res. 2009, 113, 1433–1442. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-T.; Lee, Y.-R.; Jin, J.; Han, K.-H.; Kim, H.; Kim, J.-C.; Lee, T.; Yun, S.-H.; Lee, Y.-W. Two different polyketide synthase genes are required for synthesis of zearalenone in Gibberella zeae. Mol. Microbiol. 2005, 58, 1102–1113. [Google Scholar] [CrossRef] [PubMed]
- Gaffoor, I.; Trail, F. Characterization of two polyketide synthase genes involved in zearalenone biosynthesis in Gibberella zeae. Appl. Environ. Microbiol. 2006, 72, 1793–1799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiemann, P.; Sieber, C.M.K.; von Bargen, K.W.; Studt, L.; Niehaus, E.-M.; Espino, J.J.; Huß, K.; Michielse, C.B.; Albermann, S.; Wagner, D.; et al. Deciphering the cryptic genome: Genome-wide analyses of the rice pathogen Fusarium fujikuroi reveal complex regulation of secondary metabolism and novel metabolites. PLOS Pathog. 2013, 9, e1003475. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Gavia, D.J.; Tang, Y. Biosynthesis of fungal indole alkaloids. Nat. Prod. Rep. 2014, 31, 1474–1487. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Doroghazi, J.R.; Janga, S.C.; Zhang, J.K.; Circello, B.; Griffin, B.M.; Labeda, D.P.; Metcalf, W.W. Diversity and abundance of phosphonate biosynthetic genes in nature. Proc. Natl. Acad. Sci. USA 2013, 110, 20759–20764. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain | Genome Size (Mbp) | Number of Proteins | Source/Reference |
|---|---|---|---|---|
| F. vanettenii | 77-13-4 | 54.59 | 16,929 | [15] |
| F. vanettenii | T23 | 61.89 | 18,783 | [16] |
| F. tenuicristatum | NRRL 22470 | 61.51 | 17,800 | [16] |
| F. mori | NRRL 22230 | 43.64 | 14,523 | [16] |
| F. cucurbiticola | NRRL 22165 | 42.45 | 12,147 | [16] |
| F. bataticola | NRRL 22400 | 50.41 | 16,777 | [16] |
| F. solani | FS5 | 52.93 | 17,656 | [17] |
| F. ambrosium | NRRL 20438 | 49.04 | 17,262 | NCBI |
| F. euwallaceae | UCR1854 | 50.55 | 17,630 | NCBI |
| F. floridanum | NRRL 62606 | 47.42 | 16,762 | NCBI |
| Species | Strain | Total BGC a | Total b | Hybrid BGCs | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PKS | NRPS | Terpene | DMATS | Phosphonate | PKS–PKS Hybrid | NRPS–NRPS Hybrid | NRPS–PKS Hybrid | PKS–Terpene Hybrid | NRPS–Terpene Hybrid | NRPS–DMATS Hybrid | |||
| Fusarium vanettenii | 77-13-4 | 39 | 14 | 18 | 6 | 2 | 1 | 1 | 1 | ||||
| Fusarium vanettenii | T23 | 42 | 16 | 17 | 8 | 1 | 1 | 1 | |||||
| Fusarium tenuicristatum | NRRL 22470 | 48 | 18 | 18 | 10 | 2 | 2 | 1 | 1 | ||||
| Fusarium mori | NRRL 22230 | 41 | 15 | 17 | 9 | 1 | 1 | 1 | 1 | ||||
| Fusarium cucurbiticola | NRRL 22165 | 35 | 10 | 18 | 4 | 2 | 1 | ||||||
| Fusarium bataticola | NRRL 22400 | 45 | 17 | 22 | 7 | 2 | 1 | 1 | 1 | 1 | 1 | ||
| Fusarium solani | FS5 | 43 | 15 | 22 | 6 | 2 | 1 | 2 | 1 | ||||
| Fusarium ambrosium | NRRL 20438 | 40 | 14 | 19 | 5 | 2 | 1 | 1 | |||||
| Fusarium euwallaceae | UCR1854 | 40 | 13 | 21 | 5 | 3 | 1 | 1 | 1 | 1 | |||
| Fusarium floridanum | NRRL 62606 | 38 | 14 | 16 | 6 | 2 | 1 | 1 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pokhrel, A.; Coleman, J.J. Inventory of the Secondary Metabolite Biosynthetic Potential of Members within the Terminal Clade of the Fusarium solani Species Complex. J. Fungi 2023, 9, 799. https://doi.org/10.3390/jof9080799
Pokhrel A, Coleman JJ. Inventory of the Secondary Metabolite Biosynthetic Potential of Members within the Terminal Clade of the Fusarium solani Species Complex. Journal of Fungi. 2023; 9(8):799. https://doi.org/10.3390/jof9080799
Chicago/Turabian StylePokhrel, Ambika, and Jeffrey J. Coleman. 2023. "Inventory of the Secondary Metabolite Biosynthetic Potential of Members within the Terminal Clade of the Fusarium solani Species Complex" Journal of Fungi 9, no. 8: 799. https://doi.org/10.3390/jof9080799
APA StylePokhrel, A., & Coleman, J. J. (2023). Inventory of the Secondary Metabolite Biosynthetic Potential of Members within the Terminal Clade of the Fusarium solani Species Complex. Journal of Fungi, 9(8), 799. https://doi.org/10.3390/jof9080799