
Pathogenicity, Mycotoxin Production, and Control of Potato Dry Rot Caused by Fusarium spp.: A Review

Abstract
:1. Introduction
2. Dry Rot of Potato Tubers
2.1. The Causal Agent Causing Dry Rot
2.2. Pathogen Infection and the Symptoms of Potato Dry Rot
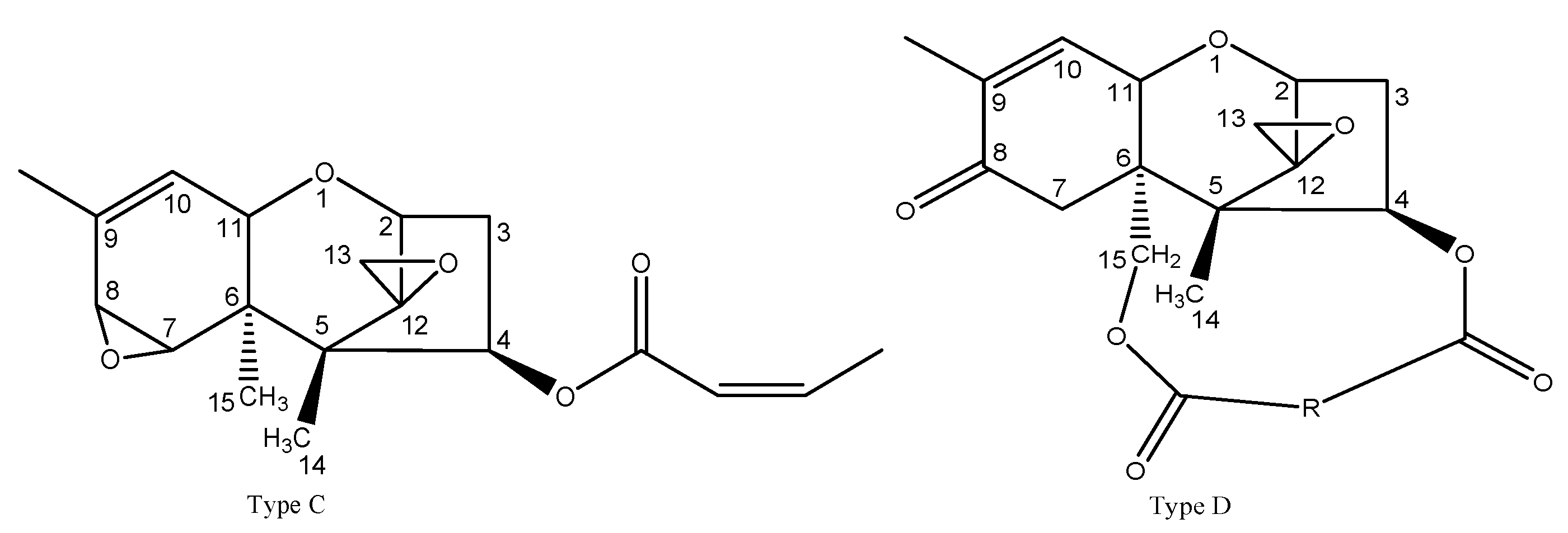
2.3. Mycotoxin Accumulation Associated with Fusarium Dry Rot
2.4. Dry Rot Control
2.4.1. Variety Screening
2.4.2. Cultivation Patterns
2.4.3. Harvesting, Grading and Packing
2.4.4. Storage Cellar Management
2.4.5. Physical, Chemical, and Biological Treatment
Physical Treatment
Chemical Treatment
Biological Treatment
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, J.; Sun, Z.Q.; Zou, Y.P.; Li, W.H.; He, F.Y.; Huang, X.Y.; Lin, C.L.; Cai, Q.N.; Wisniewski, M.; Wu, X.H. Pre- and postharvest measures used to control decay and mycotoxigenic fungi in potato (Solanum tuberosum L.) during storage. Crit. Rev. Food Sci. Nutr. 2022, 62, 415–428. [Google Scholar] [CrossRef]
- Li, L.; Zhu, T.; Song, Y.; Feng, L.; Kear, P.J.; Riseh, R.S.; Sitohy, M.; Datla, R.; Ren, M. Salicylic acid fights against Fusarium wilt by inhibiting target of rapamycin signaling pathway in Fusarium oxysporum. J. Adv. Res. 2022, 39, 1–13. [Google Scholar] [CrossRef]
- Li, Y.; Xia, X.; Zhao, Q.; Dong, P. The biocontrol of potato dry rot by microorganisms and bioactive substances: A review. Physiol. Mol. Plant Pathol. 2022, 122, 101919. [Google Scholar] [CrossRef]
- Xue, H.L.; Bi, Y.; Wei, J.M.; Tang, Y.M.; Zhao, Y.; Wang, Y. A new method for the simultaneous analysis of types A and B trichothecenes by ultrahigh-performance liquid chromatography coupled with tandem mass spectrometry in potato tubers inoculated with Fusarium sulphureum. J. Agric. Food Chem. 2013, 61, 9333–9338. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Zhang, W.; Kang, Y.; Shi, M.; Yang, X.; Yu, H.; Zhang, R.; Liu, Y.; Qin, S. Physiological and dynamic transcriptome analysis of two potato varieties reveal response of lignin and MAPK signal to dry rot caused by Fusarium sulphureum. Sci. Hortic. 2021, 289, 110470. [Google Scholar] [CrossRef]
- Wang, W.Z.; Min, F.X.; Yang, S.; Wei, Q.; Guo, M.; Gao, Y.F.; Hu, L.S.; Sheng, W.M. Research progress on potato dry rot disease in China and its control measures. China Veg. 2020, 4, 22–29. [Google Scholar]
- Xue, H.L.; Bi, Y.; Zong, Y.Y.; Alejandro, C.U.; Wang, H.J.; Pu, L.M.; Wang, Y.; Li, Y.C. Effects of elicitors on trichothecene accumulation and Tri genes expression in potato tubers inoculated with Fusarium sulphureum. Eur. J. Plant Pathol. 2017, 148, 673–685. [Google Scholar]
- Li, Y.C.; Bi, Y.; Ge, Y.H.; Sun, X.J.; Wang, Y. Antifungal activity of sodium silicate on Fusarium sulphureum and its effect on dry rot of potato tubers. J. Food Sci. 2009, 74, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Yaganza, E.S.; Tweddell, R.J.; Arul, J. Postharvest application of organic and inorganic salts to control potato (Solanum tuberosum L.) storage soft rot: Plant tissue-salt physicochemical interactions. J. Agric. Food Chem. 2014, 62, 9223–9231. [Google Scholar] [CrossRef]
- Li, Y.C.; Sun, X.J.; Bi, Y.; Ge, Y.H.; Wang, Y. Antifungal activity of chitosan on Fusarium sulphureum in relation to dry rot of potato tuber. Agric. Sci. China 2009, 8, 597–604. [Google Scholar] [CrossRef]
- Li, X.D.; Xue, H.L. Antifungal activity of the essential oil of Zanthoxylum bungeanum and its major constituent on Fusarium sulphureum and dry rot of potato tubers. Phytoparasitica 2014, 42, 509–517. [Google Scholar]
- Wei, J.; Bi, Y.; Xue, H. Antifungal activity of cinnamaldehyde against Fusarium sambucinum involves inhibition of ergosterol biosynthesis. J. Appl. Microbiol. 2020, 129, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Al-Mughrabi, K.I.; Vikram, A.; Peters, R.D.; Howard, R.J.; Grant, T.L.; Barasubiye, K.; Lynch, R.; Poirier, K.A.; Drake, I.K. Efficacy of Pseudomonas syringae in the management of critical reviews in food science and nutrition potato tuber diseases in storage. Biol. Control 2013, 64, 315–322. [Google Scholar] [CrossRef]
- Aydin, M.H. Evaluation of some Trichoderma species in biological control of potato dry rot caused by Fusarium sambucinum fuckel isolates. Appl. Ecol. Environ. Res. 2019, 17, 533–546. [Google Scholar] [CrossRef]
- Tiwari, R.K.; Kumar, R.; Sharma, S.; Sagar, V.; Aggarwal, R.; Naga, K.C.; Lal, M.K.; Chourasia, K.N.; Kumar, D.; Kumar, M. Potato dry rot disease: Current status, pathogenomics and management. Biotechnology 2020, 10, 503. [Google Scholar] [CrossRef] [PubMed]
- Ray, H.; Hammerschmidt, R. Responses of potato tuber to infection by Fusarium sambucinum. Physiol. Mol. Plant Pathol. 1998, 53, 82–91. [Google Scholar] [CrossRef]
- Wharton, P.S.; Tumbalam, P.; Kirk, W.W. First report of potato tuber sprout rot caused by Fusarium sambucinum in Michigan. Plant Dis. 2006, 90, 1460. [Google Scholar] [CrossRef] [PubMed]
- Heltoft, P.; Molteberg, E.L.; Nastad, R.; Hermansen, A. Effect of maturity level and potato cultivar on development of Fusarium dry rot in Norway. Potato Res. 2015, 58, 205–219. [Google Scholar] [CrossRef]
- Oosterhaven, K.; Chambel Leitao, A.; Gorris, L.G.M.; Smid, E.J. Comparative study on the action of S-(+)-carvone, in situ, on the potato storage fungi Fusarium solani var. coeruleum and F. sulphureum. J. Appl. Bacteriol. 1996, 80, 535–539. [Google Scholar] [CrossRef]
- Heltoft, P.; Brurberg, M.B.; Skogen, M.; Le, V.H.; Razzaghian, J.; Hermansen, A. Fusarium spp. causing dry rot on potatoes in Norway and development of a real-time PCR method for detection of Fusarium coeruleum. Potato Res. 2016, 59, 67–80. [Google Scholar] [CrossRef]
- Carnegie, S.F.; Cameron, A.M. Occurrence of Polyscytalum pustulans, Phoma foveata and Fusarium solani var. coeruleum in field soils in Scotland. Plant Pathol. 1990, 39, 517–523. [Google Scholar] [CrossRef]
- Ali, S.; Rivera, V.V.; Secor, G.A. First Report of Fusarium graminearum causing dry rot of potato in North Dakota. Plant Dis. 2005, 89, 105. [Google Scholar] [CrossRef] [PubMed]
- Villarino, M.; Cal, A.D.; Melgarejo, P.; Larena, I. Development of a multiplex PCR for the identification of Fusarium solani and F. oxysporum in a single step. J. Plant Dis. Prot. 2021, 128, 1275–1290. [Google Scholar] [CrossRef]
- Theron, D.J.; Holz, G. Effect of temperature on dry rot development of potato tubers inoculated with different Fusarium spp. Potato Res. 1990, 33, 109–117. [Google Scholar] [CrossRef]
- Esfahani, M.N. Present status of Fusarium dry rot of potato tubers in Isfahan. Indian Phytopathol. 2012, 59, 2. [Google Scholar]
- Gherbawy, Y.A.; Hussein, M.A.; El-dawy, E.G.A. Identification of Fusarium spp. associated with potato tubers in upper Egypt by morphological and molecular characters. Asian J. Biochem. Genet. Mol. Biol. 2019, 2, 1–14. [Google Scholar] [CrossRef]
- Du, M.; Ren, X.; Sun, Q.; Wang, Y.; Zhang, R. Characterization of Fusarium spp. causing potato dry rot in China and susceptibility evaluation of Chinese potato germplasm to the pathogen. Potato Res. 2012, 55, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Xue, H.L.; Bi, Y.; Tang, Y.M.; Zhao, Y.; Wang, Y. Effect of cultivars, Fusarium strains and storage temperature on trichothecenes production in inoculated potato tubers. Food Chem. 2014, 151, 236–242. [Google Scholar] [CrossRef]
- Yang, Z.M.; Bi, Y.; Li, Y.C.; Kou, Z.H.; Bao, G.H.; Liu, C.K.; Wang, Y.; Wang, D. Changes of cell wall degrading enzymes in potato tuber tissue slices infected by Fusarium sulphureum. Sci. Agric. Sin. 2012, 45, 127–134. [Google Scholar]
- Chai, Y.; Xu, Y.Q.; Fu, Y.; Li, X.Y.; He, F.M.; Han, Y.Q.; Feng, Z.; Li, F.L. Characteristics of cell wall degradation enzyme produced by main pathogenic Fusarium spp. in potato dry rot. Crops 2018, 4, 154–160. [Google Scholar]
- Bao, G.H.; Bi, Y.; Li, Y.C.; Kou, Z.H.; Hu, L.G.; Ge, Y.H.; Wang, Y.; Wang, D. Overproduction of reactive oxygen species involved in the pathogenicity of Fusarium in potato tubers, Physiol. Mol. Plant Pathol. 2014, 86, 35–42. [Google Scholar] [CrossRef]
- Zhang, X.F.; Wu, C.Y.; Hua, H.H.; Cai, Q.N.; Wu, X.H. Characterization of the first Alternavirus identified in Fusarium avenaceum, the causal agent of potato dry rot. Viruses 2023, 15, 145. [Google Scholar] [CrossRef] [PubMed]
- Vatankhah, M.; Saberi Riseh, R.; Moradzadeh Eskandari, M.; Sedaghati, E.; Alaie, H.; Afzali, H. Biological control of Fusarium dry rot of potato using some probiotic bacteria. J. Agric. Sci. Technol. 2019, 21, 1301–1312. [Google Scholar]
- Schultz, B.; Ellner, F.M.; Gossmann, M.; Buettner, C. Investigation into virulence and mycotoxin formation of the dry rot causing pathogen Fusarium sambucinum on potatoes. Mycotoxin Res. 2008, 23, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Xue, H.L.; Bi, Y.; Zhang, R.; Carelle, J.K.; Liu, Q.L.; Nan, M.N.; Pu, L.P.; Dov, P. Ozone treatment inhibits dry rot development and diacetoxyscirpenol accumulation in inoculated potato tuber by influencing growth of Fusarium sulphureum and ergosterol biosynthesis. Postharvest Biol. Technol. 2022, 185, 111796. [Google Scholar] [CrossRef]
- Song, H.H.; Lee, H.S.; Jeong, J.H.; Park, H.S.; Lee, C. Diversity in beauvericin and enniatins H, I, and MK1688 by Fusarium oxysporum isolated from potato. Int. J. Food Microbiol. 2006, 122, 296–301. [Google Scholar] [CrossRef]
- Stefańczyk, E.; Sobkowiak, S.; Brylińska, M.; Śliwka, J. Diversity of Fusarium spp. associated with dry rot of potato tubers in Poland. Eur. J. Plant Pathol. 2016, 145, 871–884. [Google Scholar] [CrossRef] [Green Version]
- Golinski, P.; Vesonder, R.F.; Latus-Zietkiewicz, D.; Perkowski, J. Formation of fusarenone X, nivalenol, zearalenone, alpha-trans-zearalenol, beta-trans-zearalenol, and fusarin C by Fusarium crookwellense. Appl. Environ. Microbiol. 1998, 54, 2147–2148. [Google Scholar] [CrossRef]
- Latus-Zietkiewicz, D.; Perkowski, J.; Tanaka, T.; Yamamoto, S.; Kawamura, O.; Sugiura, Y.; Ueno, Y. Formation of trichothecenes and zearalenone by Fusarium isolated from potato tubers. Microbiol. Aliment. Nutr. 1990, 8, 143–147. [Google Scholar]
- Sydenham, E.W.; Marasas, W.F.O.; Thiel, P.G.; Shephard, G.S.; Nieuwenhuis, J.J. Production of mycotoxins by selected Fusarium graminearum and F. crookwellense isolates. Food Addit. Contam. 1991, 8, 31–41. [Google Scholar] [CrossRef]
- El-Hassan, K.I.; El-Saman, M.G.; Mosa, A.A.; Mostafa, M.H. Variation among Fusarium spp. the causal of potato tuber dry rot in their pathogenicity and mycotoxins production. Egypt. J. Phytopathol. 2007, 35, 53–68. [Google Scholar]
- Kim, J.C.; Lee, Y.W. Sambutoxin, a new mycotoxin produced by toxic Fusarium isolates obtained from rotted potato tuber. Appl. Environ. Microbiol. 1994, 60, 4380–4386. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.C.; Lee, Y.W.; Tamura, H.; Yoshizawa, T. Sambutoxin: A new mycotoxin isolated from Fusarium sambucinum. Tetrahedron Lett. 1995, 36, 1047–1050. [Google Scholar] [CrossRef]
- Kim, J.C.; Lee, Y.W.; Yu, S.H. Sambutoxin-producing isolates of Fusarium species and occurrence of sambutoxin in rotten potato tubers. Appl. Environ. Microbiol. 1995, 61, 3750–3751. [Google Scholar] [CrossRef] [PubMed]
- Venter, S.L.; Steyn, P.J.; Hester, S.F. Production of fusaric acid by Fusarium oxysporum. Potato Res. 1996, 39, 79–83. [Google Scholar] [CrossRef]
- Venter, S.L.; Steyn, P.J. Correlation between fusaric acid production and virulence of isolates of Fusarium oxysporum that causes potato dry rot in South Africa. Potato Res. 1998, 41, 289–294. [Google Scholar] [CrossRef]
- Zhang, R.; Xue, H.L.; Li, L.; Bi, Y.; Zong, Y.Y.; Jimdjio, K.C. Study on the adsorption effect of diatomite on neosolaniol (NEO) in muskmelon fruits inoculated with Fusarium sulphureum. Int. J. Food Eng. 2021, 17, 257–264. [Google Scholar] [CrossRef]
- Tang, Y.M.; Xue, H.L.; Bi, Y.; Li, Y.C.; Wang, Y.; Zhao, Y.; Shen, K.P. A method of analysis for T-2 toxin and neosolaniol by UPLC-MS/MS in apple fruit inoculated with Trichothecium roseum. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2015, 32, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Pierron, A.; Neves, M.; Puel, S.; Lippi, Y.; Soler, L.; Miller, J.D.; Oswald, I.P. Intestinal toxicity of the new type A trichothecenes, NX and 3ANX. Chemosphere 2022, 288, 132415. [Google Scholar] [CrossRef] [PubMed]
- Ellner, F.M. Mycotoxins in potato tubers infected by Fusarium sambucinum. Mycotoxin Res. 2002, 18, 57–61. [Google Scholar] [CrossRef]
- Delgado, J.A.; Schwarz, P.B.; Gillespie, J.; Rivera-Varas, V.V.; Secor, G.A. Trichothecene mycotoxins associated with potato dry rot caused by Fusarium graminearum. Phytopathology 2010, 100, 290–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desjardins, A.E.; Plattner, R.D. Trichothecene toxin production by strains of Gibberella pulicaris (Fusarium sambucinum) in liquid culture and in potato tubers. J. Agric. Food Chem. 1989, 37, 388–392. [Google Scholar] [CrossRef]
- Jelen, H.H.; Mirocha, C.J.; Wasowicz, E.; Kaminski, E. Production of volatile sesquiterpenes by Fusarium sambucinum strains with different abilities to synthesize trichothecenes. Appl. Environ. Microbiol. 1995, 61, 3815–3820. [Google Scholar] [CrossRef]
- El-Banna, A.A.; Scott, P.M.; Lau, P.Y.; Sakuma, T.; Platt, H.W.; Campbell, V. Formation of trichothecenes by Fusarium solani var. coeruleum and Fusarium sambucinum in potatoes. Appl. Environ. Microbiol. 1984, 47, 1169–1171. [Google Scholar] [CrossRef]
- Lenc, L.; Lukanowski, A.; Sadowski, C. The use of PCR amplification in determining the toxigenic potential of Fusarium sambucinum and F. solani isolated from potato tubers with symptoms of dry rot. Phytopathol. Pol. 2008, 2008, 48. [Google Scholar]
- Vesonder, R.F.; Golinski, P.; Plattner, R.; Zietkiewicz, D.L. Mycotoxin formation by different geographic isolates of Fusarium crookwellense. Mycopathologia 1991, 113, 11–14. [Google Scholar] [CrossRef]
- Nielsen, K.F.; Thrane, U. Fast methods for screening of trichothecenes in fungal cultures using gas chromatography–tandem mass spectrometry. J. Chromatogr. A 2001, 929, 75–87. [Google Scholar] [CrossRef]
- Burlakoti, R.R.; Estrada, R.; Rivera, V.V.; Boddeda, A.; Secor, G.A.; Adhikari, T.B. Real-time PCR quantification and mycotoxin production of Fusarium graminearum in wheat inoculated with isolates collected from potato, sugar beet, and wheat. Phytopathology 2007, 97, 835–841. [Google Scholar] [CrossRef] [Green Version]
- Burlakoti, R.R.; Ali, S.; Secor, G.A.; Neate, S.M.; McMullen, M.P.; Adhikari, T.B. Genetic relationships among populations of Gibberella zeae from barley, wheat, potato, and sugar beet in the Upper Midwest of the United States. Phytopathology 2008, 98, 969–976. [Google Scholar] [CrossRef] [Green Version]
- Latus-Zietkiewicz, D.; Perkowski, J.; Chełkowski, J. Fusarium species as pathogens of potato tubers during storage and their ability to produce mycotoxins. Mycotoxin Res. 1987, 3, 99–104. [Google Scholar] [CrossRef]
- Shams, M.; Mitterbauer, R.; Corradini, R.; Wiesenberger, G.; Dall’Asta, C.; Schuhmacher, R.; Krska, R.; Adam, G.; Berthiller, F. Isolation and characterization of a new less-toxic derivative of the Fusarium mycotoxin diacetoxyscirpenol after thermal treatment. J. Agric. Food Chem. 2011, 59, 9709–9714. [Google Scholar] [CrossRef] [PubMed]
- Zaheer, K.; Akhtar, M.H. Potato production, usage, and Nutrition—A Review. Crit. Rev. Food Sci. Nutr. 2016, 56, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Trabelsi, B.M.; Abdallah, R.A.B.; Ammar, N.; Kthiri, Z.; Hamada, W. Bio-suppression of Fusarium wilt disease in potato using nonpathogenic potatoassociated fungi. J. Plant Pathol. Microbiol. 2016, 7, 347–354. [Google Scholar]
- Esfahani, M.N. Susceptibility assessment of potato cultivars to Fusarium dry rot species. Potato Res. 2005, 48, 215–226. [Google Scholar] [CrossRef]
- Yilma, S.; Vales, M.I.; Charlton, B.A.; Hane, D.C.; James, S.R.; Shock, C.C.; Mosley, A.R.; Culp, D.; Feibert, E.; Leroux, L.; et al. Owyhee Russet: A variety with high yields of U.S. No. 1 tubers, excellent processing quality, and moderate resistance to Fusarium dry rot (Fusarium solani var. coeruleum). Am. J. Potato Res. 2012, 89, 175–183. [Google Scholar] [CrossRef]
- Valluru, R.; Christ, B.J.; Haynes, K.G.; Vinyard, B.T. Inheritance and stability of resistance to Fusarium tuber rot in tetraploid potatoes. Am. J. Potato Res. 2006, 83, 335–341. [Google Scholar] [CrossRef]
- Burkhart, C.R.; Christ, B.J.; Haynes, K.G. Non-additive genetic variance governs resistance to Fusarium dry rot in a diploid hybrid potato population. Am. J. Potato Res. 2007, 84, 199–204. [Google Scholar] [CrossRef]
- Mejdoub-Trabelsi, B.; Jabnoun-Khiareddine, H.; Daami-Remadi, M. Effect of Fusarium species and temperature of storage on the susceptibility ranking of potato cultivars to tuber dry rot biocontrol of soilborne fungal diseases of vegetable crops. Pest Technol. 2012, 6, 41–46. [Google Scholar]
- Chen, D.; Nahar, K.; Bizimungu, B. A simple and efficient inoculation method for Fusarium dry rot evaluations in potatoes. Am. J. Potato Res. 2020, 97, 265–271. [Google Scholar] [CrossRef]
- Nadakuduti, S.S.; Starker, C.G.; Voytas, D.F.; Buell, C.R.; Douches, D.S. Genome editing in potato with CRISPR/Cas9. Methods Mol. Biol. 2019, 1917, 183–201. [Google Scholar]
- Veillet, F.; Perrot, L.; Chauvin, L.; Kermarrec, M.P.; Guyon-Debast, A.; Chauvin, J.E.; Nogue, F.; Mazier, M. Transgene-free genome editing in tomato and potato plants using agrobacterium-mediated delivery of a CRISPR/Cas9 cytidine base editor. Int. J. Mol. Sci. 2019, 20, 402. [Google Scholar] [CrossRef] [Green Version]
- Bojanowski, A.; Avis, T.J.; Pelletier, S.; Tweddell, R.J. Management of potato dry rot. Postharvest Biol. Technol. 2013, 84, 99–109. [Google Scholar] [CrossRef]
- Tiwari, R.K.; Kumar, R.; Sharma, S. Continuous and emerging challenges of silver scurf disease in potato. Int. J. Pest Manag. 2021, 68, 89–101. [Google Scholar] [CrossRef]
- Qin, J.H.; Bian, C.S.; Duan, S.G.; Wang, W.X.; Li, G.C.; Jin, L.P. Effects of different rotation cropping systems on potato yield, rhizosphere microbial community and soil biochemical properties. Front. Plant Sci. 2022, 13, 999730. [Google Scholar] [CrossRef] [PubMed]
- Kwambai, T.K.; Griffin, D.; Nyongesa, M.; Byrne, S.; Gorman, M.; Struik, P.C. Dormancy and physiological age of seed tubers from a diverse set of potato cultivars grown at different altitudes and in different seasons in Kenya. Potato Res. 2023, 23, 1–25. [Google Scholar] [CrossRef]
- Zhou, H.L.; Lei, Y.; Wang, P.; Liu, M.Y.; Hu, X.X. Development of SYBR Green real-time PCR and nested RT-PCR for the detection of Potato Mop-top Virus (PMTV) and viral surveys in Progeny tubers derived from PMTV infected Potato tubers. Mol. Cell. Probes 2019, 47, 101438. [Google Scholar] [CrossRef]
- Knowles, N.R.; Plissey, E.S. Maintaining tuber health during harvest, storage, and post-storage handling. In Potato Health Management; Johnson, D.A., Ed.; APS Press: St. Paul, MN, USA, 2008; pp. 79–99. [Google Scholar]
- Han, Y.; Yang, R.R.; Wang, Q.H.; Wang, B.; Prusky, D. Sodium silicate promotes wound healing by inducing the deposition of suberin polyphenolic and lignin in potato tubers, Front. Plant Sci. 2022, 13, 942022. [Google Scholar]
- Han, Y.; Yang, R.R.; Zhang, X.J.; Wang, Q.H.; Wang, B.; Zheng, X.Y.; Li, Y.C.; Prusky, D.; Bi, Y. Brassinosteroid accelerates wound healing of potato tubers by activation of reactive oxygen metabolism and phenylpropanoid metabolism. Foods 2022, 11, 906. [Google Scholar] [CrossRef]
- Jiang, H.; Li, X.; Ma, L.; Ren, Y.Y.; Bi, Y.; Prusky, D. Transcriptome sequencing and differential expression analysis of natural and BTH-treated wound healing in potato tubers (Solanum tuberosum L.). BMC Genom. 2022, 23, 263. [Google Scholar] [CrossRef]
- Su, Q.H.; Kondo, N.; Li, M.Z.; Sun, H.; Al Riza, D.F.; Habaragamuwa, H. Potato quality grading based on machine vision and 3D shape analysis. Comput. Electron. Agric. 2018, 152, 261–268. [Google Scholar] [CrossRef]
- Pinhero, R.G.; Coffin, R.; Yada, R.Y. Post-harvest storage of potatoes. In Advances in Potato Chemistry and Technology; Elsevier: Amsterdam, The Netherlands, 2009; pp. 339–370. [Google Scholar]
- Romanazzi, G.; Smilanick, J.L.; Feliziani, E.; Droby, S. Integrated management of postharvest gray mold on fruit crops. Postharvest Biol. Technol. 2016, 113, 69–76. [Google Scholar] [CrossRef]
- Jakubowski, T. Use of UV-C radiation for reducing storage losses of potato tubers. Bangladesh J. Bot. 2018, 47, 533–537. [Google Scholar] [CrossRef] [Green Version]
- Ranganna, B.; Kushalappa, A.C.; Raghavan, G.S.V. Ultraviolet irradiance to control dry rot and soft rot of potato in storage. Can. J. Plant Pathol. 1997, 19, 30–35. [Google Scholar] [CrossRef]
- Yu, B.; Nan, Y.Y.; Kong, N.N.; Dang, R.M.; Bai, Z.L.; Liu, J. Effect of short-wave Ultraviolet irradiation on dry rot and relevant enzymes activities of post-harvest potato tubers. J. Nucl. Agric. Sci. 2017, 31, 1730–1736. [Google Scholar]
- Jakubowski, T.; Krolczyk, J.B. Method for the reduction of natural losses of potato tubers during their long-term storage. Sustainability 2020, 12, 1048. [Google Scholar] [CrossRef] [Green Version]
- Chudinova, E.M.; Kokaeva, Y.L.; Elansky, S.N.; Kutuzova, I.A.; Pertsev, A.S.; Pobendinskaya, M.A. The occurrence of thiabendazole-resistant isolates of Helminthosporium solani on potato seed tubers in Russia. J. Plant Dis. Prot. 2020, 127, 421–423. [Google Scholar] [CrossRef]
- Gachango, E.; Hanson, L.E.; Rojas, A. Fusarium spp. causing dry rot of seed potato tubers in Michigan and their sensitivity to fungicides. Plant Dis. 2012, 96, 1767–1774. [Google Scholar] [CrossRef] [Green Version]
- Malyuga1, A.A.; Chulikova, N.S.; Ilyin, M.M.; Khalikov, S.S. Fludioxonil-based preparations for protecting potatoes from diseases and their effectiveness. Russ. Agric. Sci. 2022, 48, S74–S83. [Google Scholar] [CrossRef]
- Raigond, P.; Sagar, V.; Mishra, T. Chitosan: A safe alternative to synthetic fungicides to manage dry rot in stored potatoes. Potato Res. 2019, 62, 393–409. [Google Scholar] [CrossRef]
- Xue, H.; Bi, Y.; Prusky, D. The mechanism of induced resistance against Fusarium dry rot in potato tubers by the T-2 toxin. Postharvest Biol. Technol. 2019, 153, 69–78. [Google Scholar] [CrossRef]
- Ma, L.; Jiang, H.; Ren, Y.Y.; Yang, J.W.; Han, Y.; Si, H.J.; Prusky, D.; Bi, Y.; Wang, Y. Overexpression of StCDPK23 promotes wound healing of potato tubers by regulating StRbohs. Plant Physiol. Biochem. 2022, 185, 279–289. [Google Scholar] [CrossRef]
- Hay, W.T.; Fanta, G.F.; Rich, J.O. Antifungal activity of a fatty ammonium chloride amylose inclusion complex against Fusarium sambucinum; control of dry rot on multiple potato varieties. Am. J. Potato Res. 2019, 96, 79–85. [Google Scholar] [CrossRef]
- Mahmoud, G.A.; El-Tobgy, K.M.K.; Abo-El-Seoud, M.A. Utilisation of biocides for controlling pest attacks on potato tubers. Arch Phytopathol. Plant Prot. 2010, 43, 251–258. [Google Scholar] [CrossRef]
- Velluti, A.; Sanchis, V.; Ramos, A.J.; Turon, C.; Marin, S. Impact of essential oils on growth rate, zearalenone and deoxynivalenol production by Fusarium graminearum under different temperature and water activity conditions in maize grain. J. Appl. Microbiol. 2004, 96, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Bhavaniramya, S.; Vishnupriya, S.; Al-Aboody, M.S.; Vijayakumar, R.; Baskaran, D. Role of essential oils in food safety: Antimicrobial and antioxidant applications. Grain Oil Sci. Technol. 2019, 2, 49–55. [Google Scholar] [CrossRef]
- Othmane, M.; Djazouli, Z.; Zebib, B. Aqueous extract of algerian nettle (Urtica dioïca L.) as possible alternative pathway to control some plant diseases. Iran. J. Sci. Technol. Sci. Trans. A 2021, 45, 463–468. [Google Scholar]
- Boivin, M.; Nathalie, B.; Simon, B.; Isabel, D. Black spruce extracts reveal antimicrobial and sprout suppressive potentials to prevent potato (Solanum tuberosum L.) losses during storage. J. Agric. Food Sci. 2021, 5, 100187. [Google Scholar] [CrossRef]
- Mvuemba, H.; Green, S.; Tsopmo, A.; Avis, T. Antimicrobial efficacy of cinnamon, ginger, horseradish and nutmeg extracts against spoilage pathogens. Phytoprotection 2009, 90, 65–70. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Tian, S.L.; Shen, J.; Wang, X.Z.; Cheng, J.X.; Li, S.Q.; Ge, X.; Tian, J.C. Effects of chlorine dioxide on morphology and ultrastructure of Fusarium sulphureum and its virulence to potato tubers. Int. J. Agric. Biol. Eng. 2017, 10, 242–250. [Google Scholar]
- Liu, Q.L.; Zhang, R.; Xue, H.L.; Bi, Y.; Li, L.; Zhang, Q.Q.; Carelle, J.K.; Nan, M.N.; Prusky, D. Ozone controls potato dry rot development and diacetoxyscirpenol accumulation by targeting the cell membrane and affecting the growth of Fusarium sulphureus. Physiol. Mol. Plant Pathol. 2022, 118, 101785. [Google Scholar] [CrossRef]
- Liu, Z.G.; Yang, X.; Xue, H.L.; Bi, Y.; Zhang, Q.Q.; Liu, Q.L.; Chen, J.Y.; Nan, M.N.; Prusky, D. Reactive oxygen species metabolism and diacetoxyscirpenol biosynthesis modulation in potato tuber inoculated with ozone-treated Fusarium sulphureum. J. Food Process. Preserv. 2023, 2023, 4823679. [Google Scholar] [CrossRef]
- Schisler, D.A.; Slininger, P.J. Selection and performance of bacterial strains for biologically controlling Fusarium dry rot of potatoes incited by Gibberella pulicaris. Plant Dis. 1994, 78, 251–255. [Google Scholar] [CrossRef]
- Schisler, D.A.; Slininger, P.J.; Bothast, R.J. Effects of antagonist cell concentration and two-strain mixtures on biological control of Fusarium dry rot of potatoes. Phytopathology 1997, 87, 177–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Çakar, G.; Tozlu, E. The biological control of Fusarium oxysporum, the causal agent of potato rot. Gesunde Pflanz. 2022, 74, 305–315. [Google Scholar] [CrossRef]
- Daami-Remadi, M.; Hibar, K.; Jabnoun-Khiareddine, H.; Ayed, F.; El Mahjoub, M. Effect of two Trichoderma species on severity of potato tuber dry rot caused by Tunisian Fusarium complex. Int. J. Agric. Res. 2006, 1, 432–441. [Google Scholar]
- El-Kot, G.A.N. Biological control of black scurf and dry rot of potato. Egypt J. Phytopathol. 2008, 36, 45–56. [Google Scholar]
- Paul, N.C.; Park, S.; Liu, H.F.; Lee, J.G.; Han, G.H.; Kim, H.; Sang, H. Fungi associated with postharvest diseases of sweet potato storage roots and in vitro antagonistic assay of Trichoderma harzianum against the diseases. J. Fungi 2021, 7, 927. [Google Scholar] [CrossRef]
- Wharton, P.S.; Kirk, W.W. Evaluation of biological seed treatments in combination with management practices for the control of Fusarium dry rot of potato. Biocontrol Control 2014, 73, 23–30. [Google Scholar] [CrossRef]
- Hussain, T.; Khan, A.A.; Mohamed, H.I. Metabolites composition of Bacillus subtilis Hussaint-Amu determined by LC-MS and their effect on Fusarium dry rot of potato seed tuber. Phyton-Int. J. Exp. Bot. 2023, 92, 783–799. [Google Scholar] [CrossRef]
- Yu, X.Y.; Bi, Y.; Yan, L.; Liu, X.; Wang, Y.; Shen, K.P.; Li, Y.C. Activation of phenylpropanoid pathway and PR of potato tuber against Fusarium sulphureum by fungal elicitor from Trichothecium roseum. World J. Microbiol. Biotechnol. 2016, 32, 142. [Google Scholar] [CrossRef]
- Recep, K.; Fikrettin, S.; Erkol, D.; Cafer, E. Biological control of the potato dry rot caused by Fusarium species using PGPR strains. Biol. Control 2009, 50, 194–198. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fusarium Species | Region | Reference |
|---|---|---|
| F. sambucinum | North American and some regions of Europe | [16,17,18] |
| F. coeruleum | United Kingdom and Great Britain | [19,20,21] |
| F. graminearum | North Dakota | [22] |
| F. solani and | South Africa | [23,24] |
| F. oxysporum | Michigan | |
| F. sulphureum and F. solani | Iran | [25] |
| F. sambucinum | Egypt | [26] |
| F. oxysporum | ||
| F. verticillioides | ||
| F. incarnatum | ||
| F. avenaceum, | Heilongjiang Province and Inner Mongolia Autonomous Region | [27] |
| F. oxysporum, | ||
| F. sporotrichiodes | ||
| F. solani, | ||
| F. trichothecioides, | ||
| F. solani var. coeruleum | ||
| F. sambucinum | ||
| F. semitectum, | ||
| F. solani | ||
| F. sambucinum | ||
| F. culmorum, | ||
| F. gibbosum, | ||
| F. macroceras, | ||
| F. solani var. coeruleum, F. acuminatum, | ||
| F. equiseti and F. redolens | ||
| F. sambucinem | North of China | [27] |
| F. avenaceum | Shanxi Province | |
| F. solani var. coeruleum, | ||
| F. oxysporum, | ||
| F. acuminatum | ||
| F. sambucinem | Northwest of China, | [7,8,10,11,12,27] |
| F. avenaceum | Gansu Province, Ningxia Hui Autonomous Region | |
| F. graminearum | ||
| F. solani | ||
| F. sulphureum | ||
| F. tricinctum, | Northwest of China, Qinghai Province | [6] |
| F. avenaceum, | ||
| F. oxysporum, | ||
| F. solani, | ||
| F. acuminatum, | ||
| F. equiseti | ||
| F. solani, | South of China Zhejiang Province | [6] |
| F. moniliform, | ||
| F. redolens |
| Fusarium Species | Non-Trichothecenes | Reference |
|---|---|---|
| F. oxysporum | BEA, ENNs | [36] |
| F. sambucinum | ZEA, FUS | [37] |
| F. solani | ZEA, FUS | [37] |
| F. oxysporum | ZEA, FUS | [37] |
| F. crookwellense | ZEA, FUS | [38] |
| F. crookwellense | ZEA, FUS | [39] |
| F. crookwellense | ZEA, FUS | [40] |
| F. graminearum | ZEA | [40] |
| F. equiseti | FUM, ZEA | [41] |
| F. oxysporum | FA, FUM, ZEA | [41] |
| F. sambucinum | SAM | [42] |
| F. sambucinum | SAM | [43] |
| F. sambucinum | SAM | [44] |
| F. oxysporum | FA | [45] |
| F. oxysporum | FA | [46] |
| Fusarium Species | Trichothecenes | Reference |
|---|---|---|
| F. sambucinum | trichothecene | [37] |
| F. solani | trichothecene | [37] |
| F. oxysporum | trichothecene | [37] |
| F. crookwellense | NIV, FX | [38] |
| F. sambucinum | DAS | [39] |
| F. crookwellense | NIV, FX | [39] |
| F. sulphureum | 3-ADON, T-2, FUS, DAS | [4] |
| F. solani | 3-ADON, T-2, FUS, DAS | [4] |
| F. sambucinum | 3-ADON, T-2, FUS, DAS | [4] |
| F. equiseti | T-2 | [41] |
| F. oxysporum | T-2 | [41] |
| F. sambucinum | DAS | [50] |
| F. sambucinum | DAS, MAS, NEO, T-2, HT-2 | [52] |
| F. sambucinum | 4,15-DAS, 15-MAS, 4-MASc | [53] |
| F. solani | DON, HT-2, 3-ADON | [54] |
| F. sambucinum | DON, NIV, HT-2 | [54] |
| F. sambucinum | MAS, DAS | [55] |
| F. crookwellense | NIV, DAS | [56] |
| F. culmorum | NIV, FX, 3-ADON, DON | [57] |
| F. crookwellense | FX | [57] |
| F. equiseti | NIV, FX, 4-MAS, 15-MAS, DAS, SCR | [57] |
| F. graminearum | NIV, FX, DON, 3-ADON, 15-ADON | [57] |
| F. sambucinum | SCR, 4-MAS, 15-MAS, DAS, NEO, HT-2,T-2 | [57] |
| F. graminearum | DON, NIV, FX, 3-ADON, 15- ADON | [51] |
| F. graminearum | NIV, T-2, 3-ADON, 15-ADON,15-SCR | [58] |
| F. graminearum | DON, 3ADON, 15-ADON | [59] |
| F. culmorum | DON, 3-ADON | [60] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, H.; Liu, Q.; Yang, Z. Pathogenicity, Mycotoxin Production, and Control of Potato Dry Rot Caused by Fusarium spp.: A Review. J. Fungi 2023, 9, 843. https://doi.org/10.3390/jof9080843
Xue H, Liu Q, Yang Z. Pathogenicity, Mycotoxin Production, and Control of Potato Dry Rot Caused by Fusarium spp.: A Review. Journal of Fungi. 2023; 9(8):843. https://doi.org/10.3390/jof9080843
Chicago/Turabian StyleXue, Huali, Qili Liu, and Zhimin Yang. 2023. "Pathogenicity, Mycotoxin Production, and Control of Potato Dry Rot Caused by Fusarium spp.: A Review" Journal of Fungi 9, no. 8: 843. https://doi.org/10.3390/jof9080843
APA StyleXue, H., Liu, Q., & Yang, Z. (2023). Pathogenicity, Mycotoxin Production, and Control of Potato Dry Rot Caused by Fusarium spp.: A Review. Journal of Fungi, 9(8), 843. https://doi.org/10.3390/jof9080843