
Exploring the Diversity and Systematics of Phaeosphaeriaceae: Taxonomic Novelties from Ecologically Diverse Habitats and Their Phylogenetic Resolution



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolates and Specimens
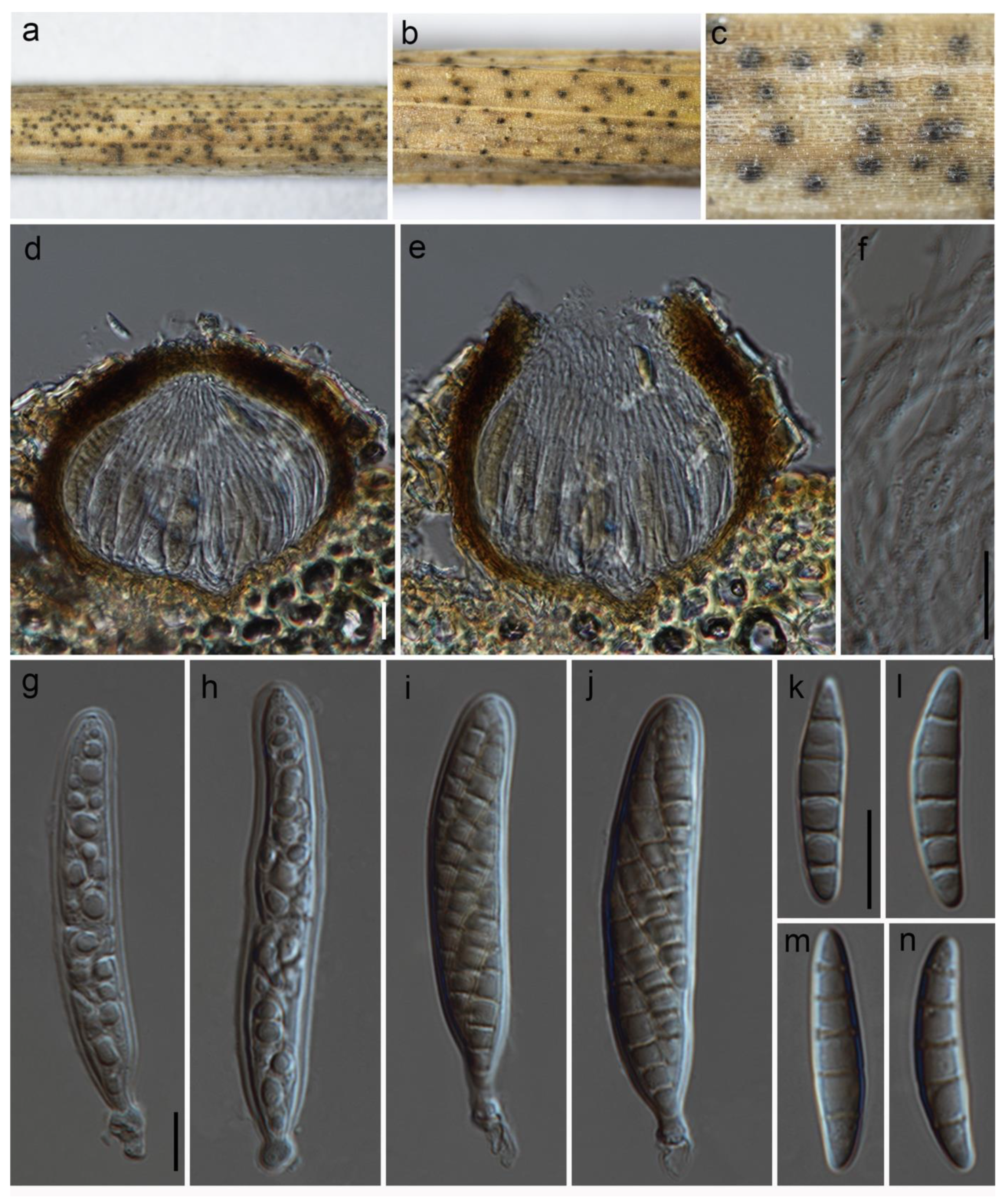
2.2. Morphological Observations
2.3. DNA Extraction, PCR Amplifications, and Sequencing
2.4. Molecular Phylogenetic Analyses
2.4.1. Sequencing and Sequence Alignment
2.4.2. Phylogenetic Analyses
3. Results
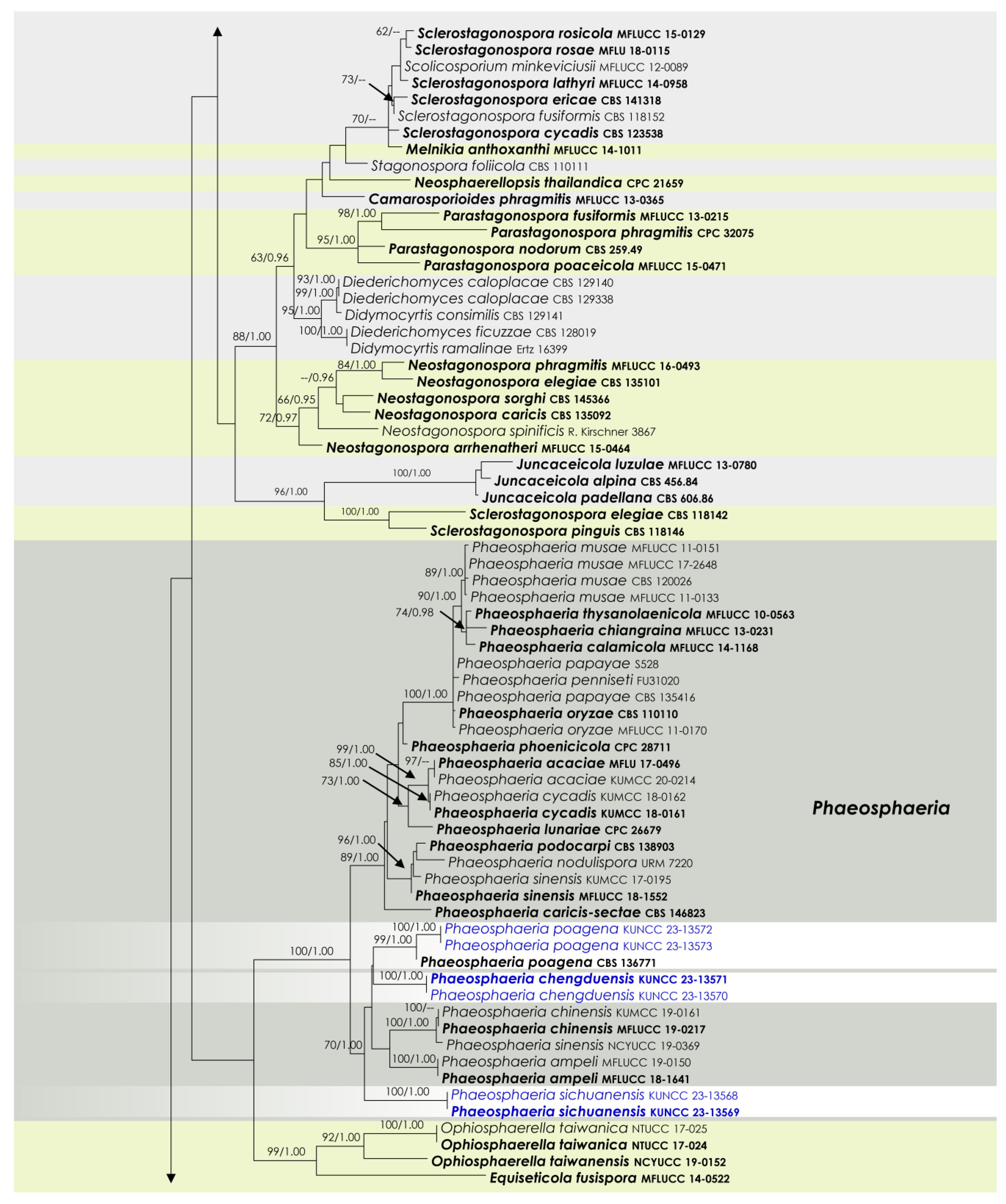
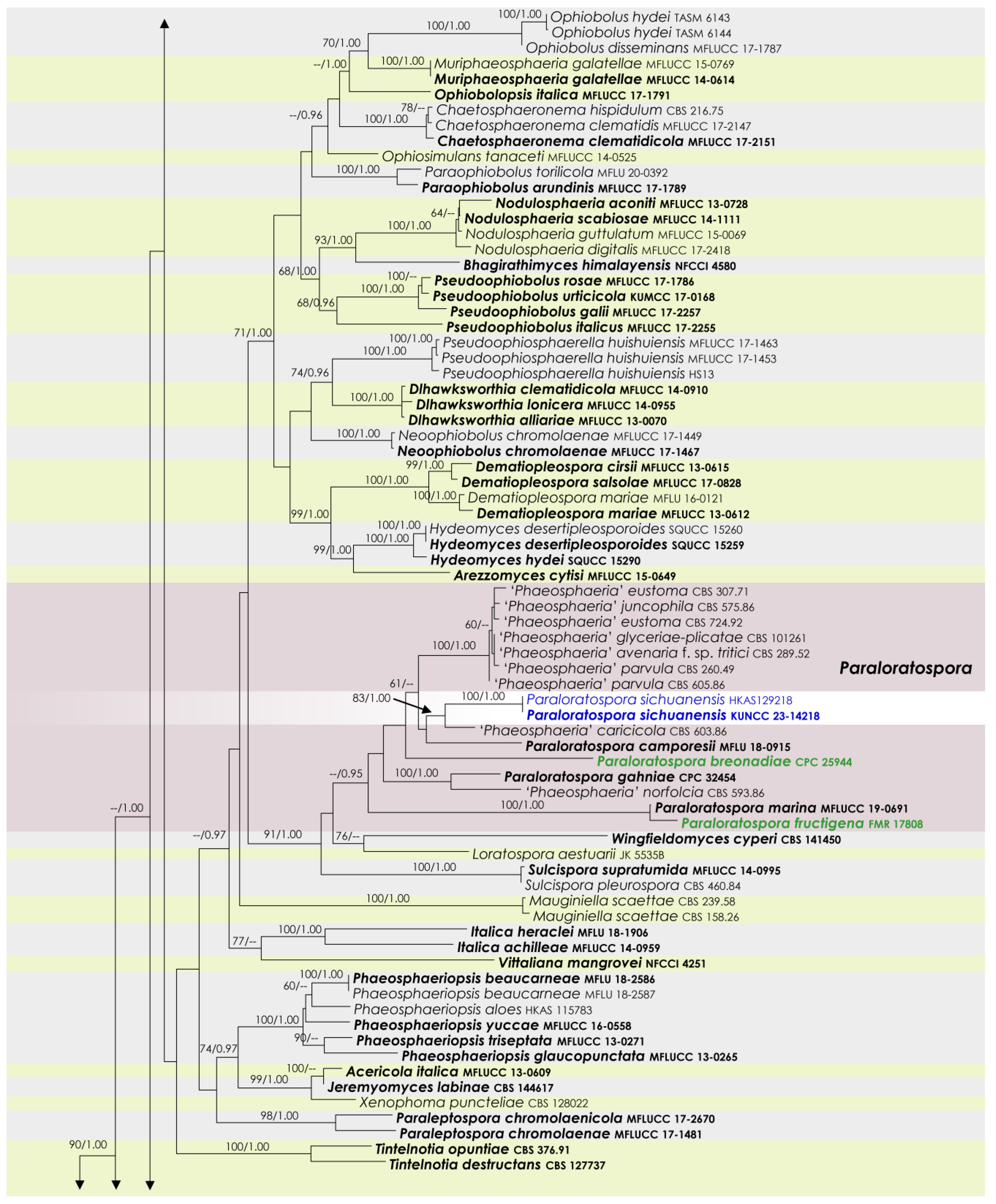
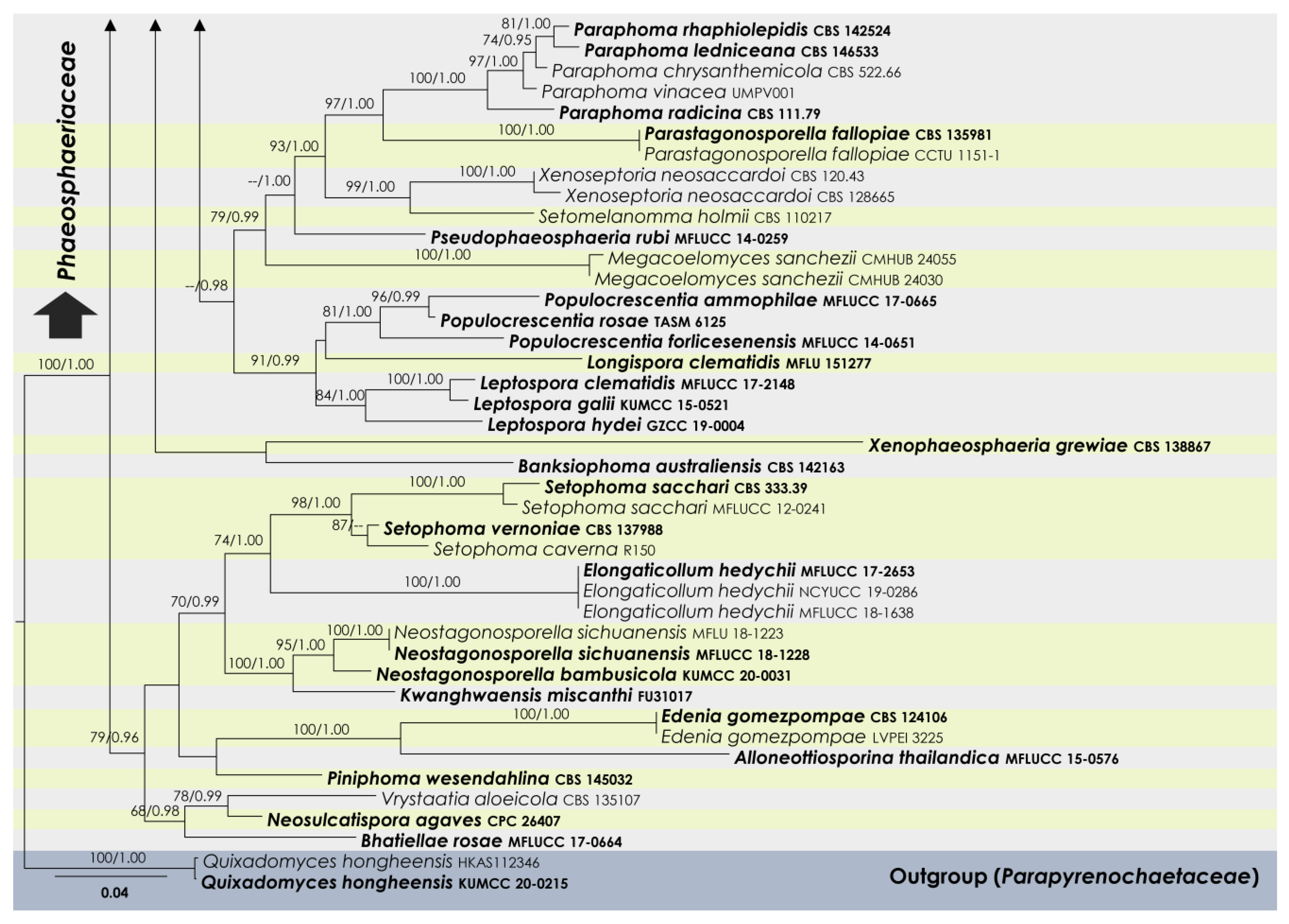
3.1. Phylogenetic Analyses
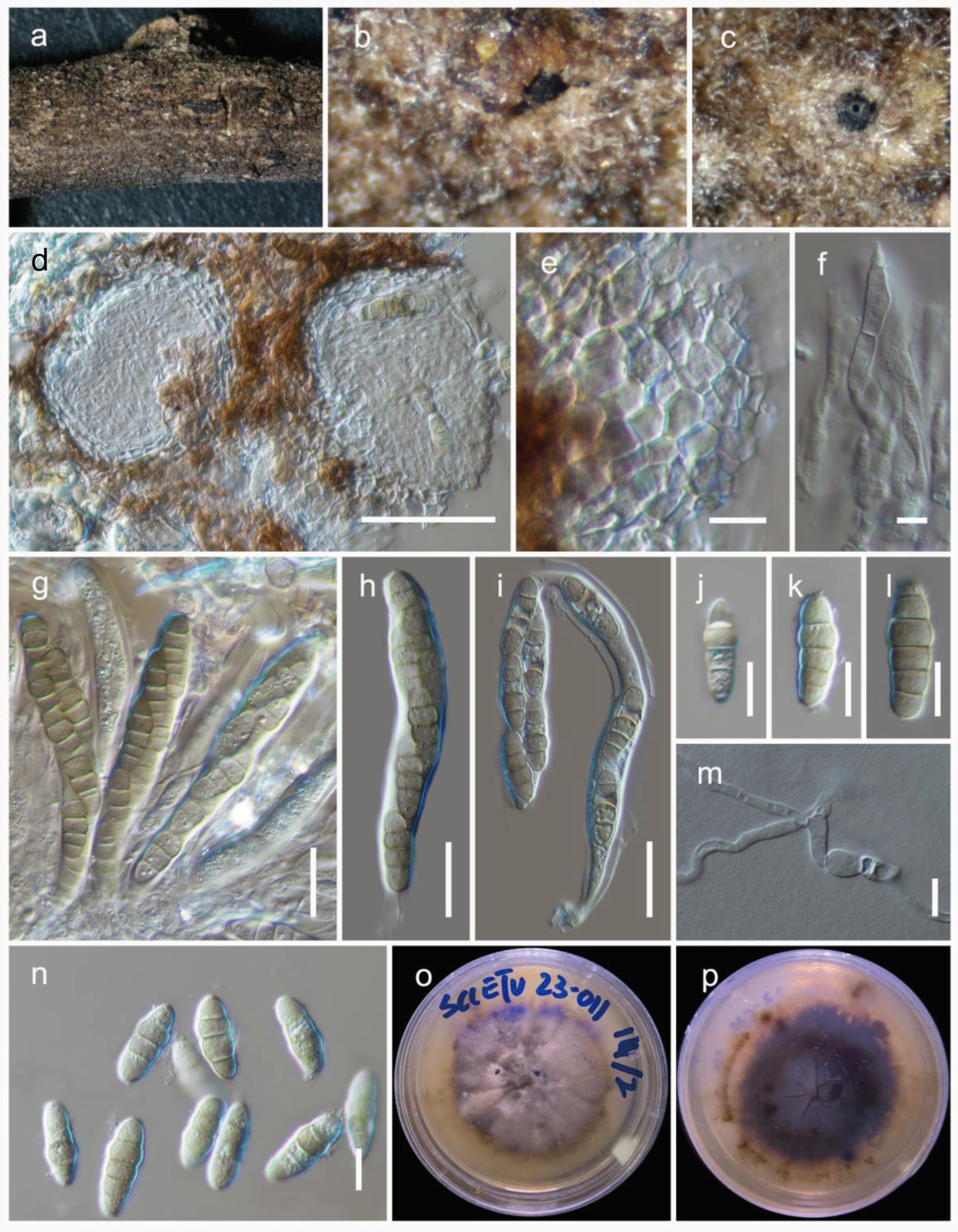
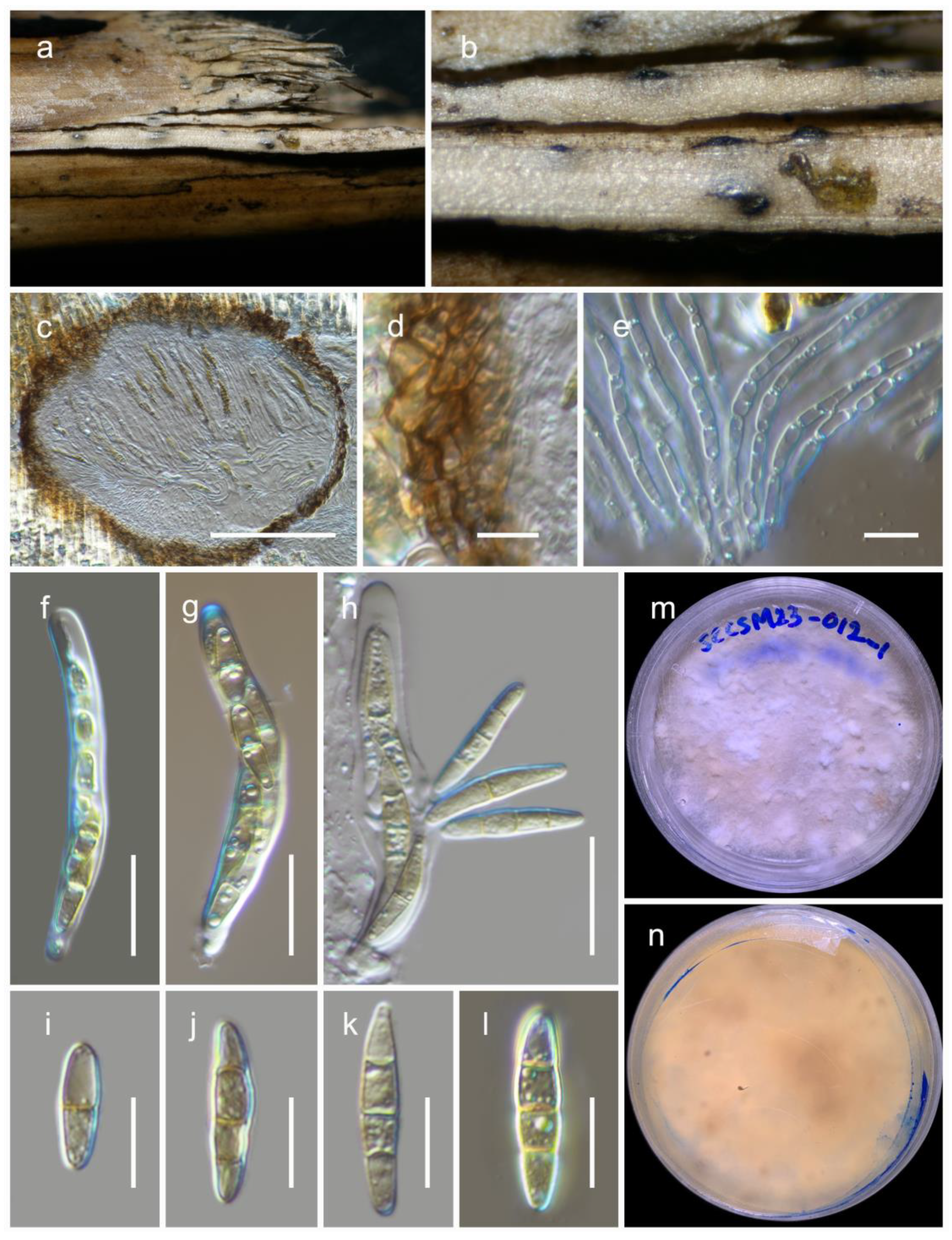
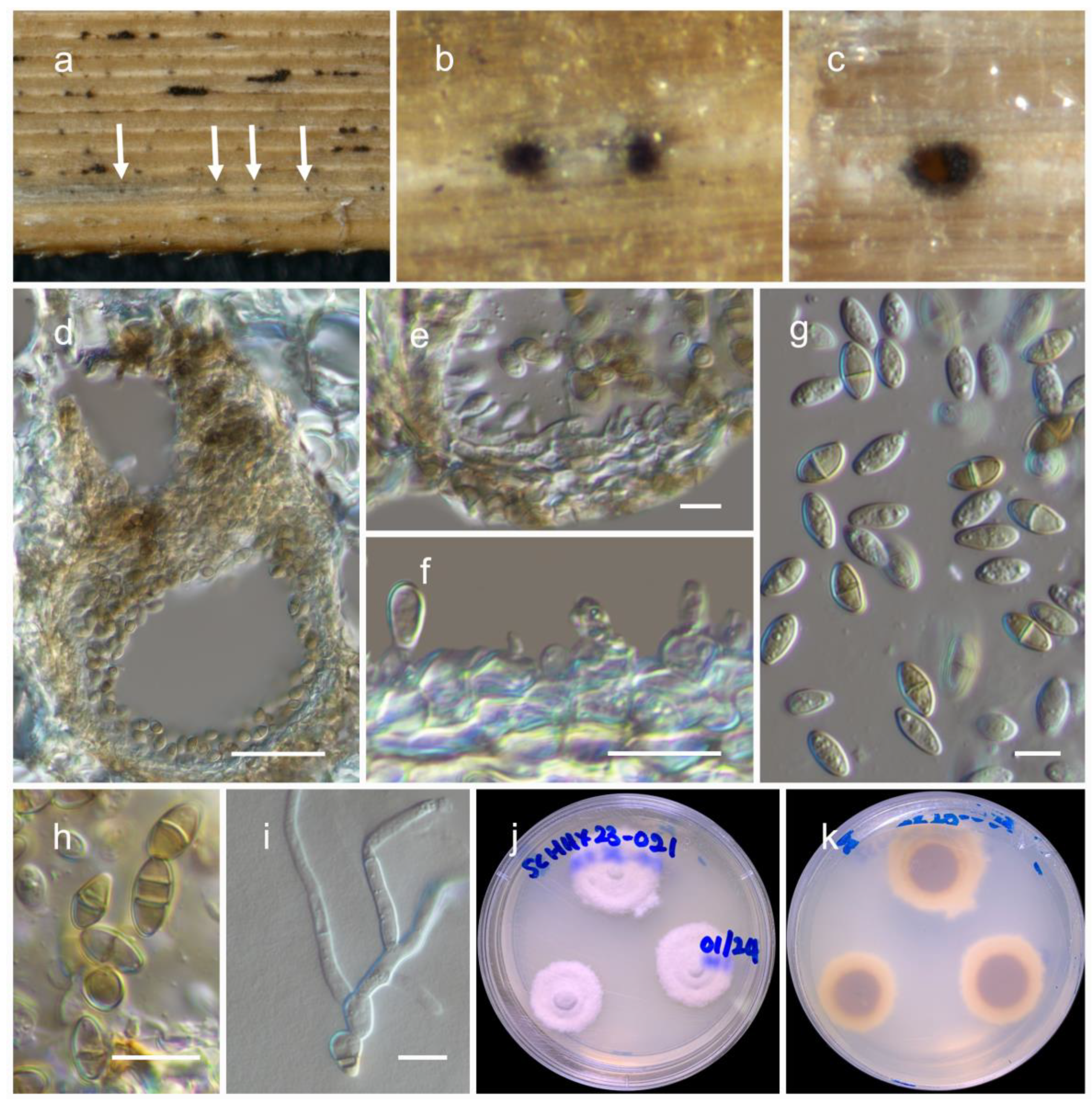
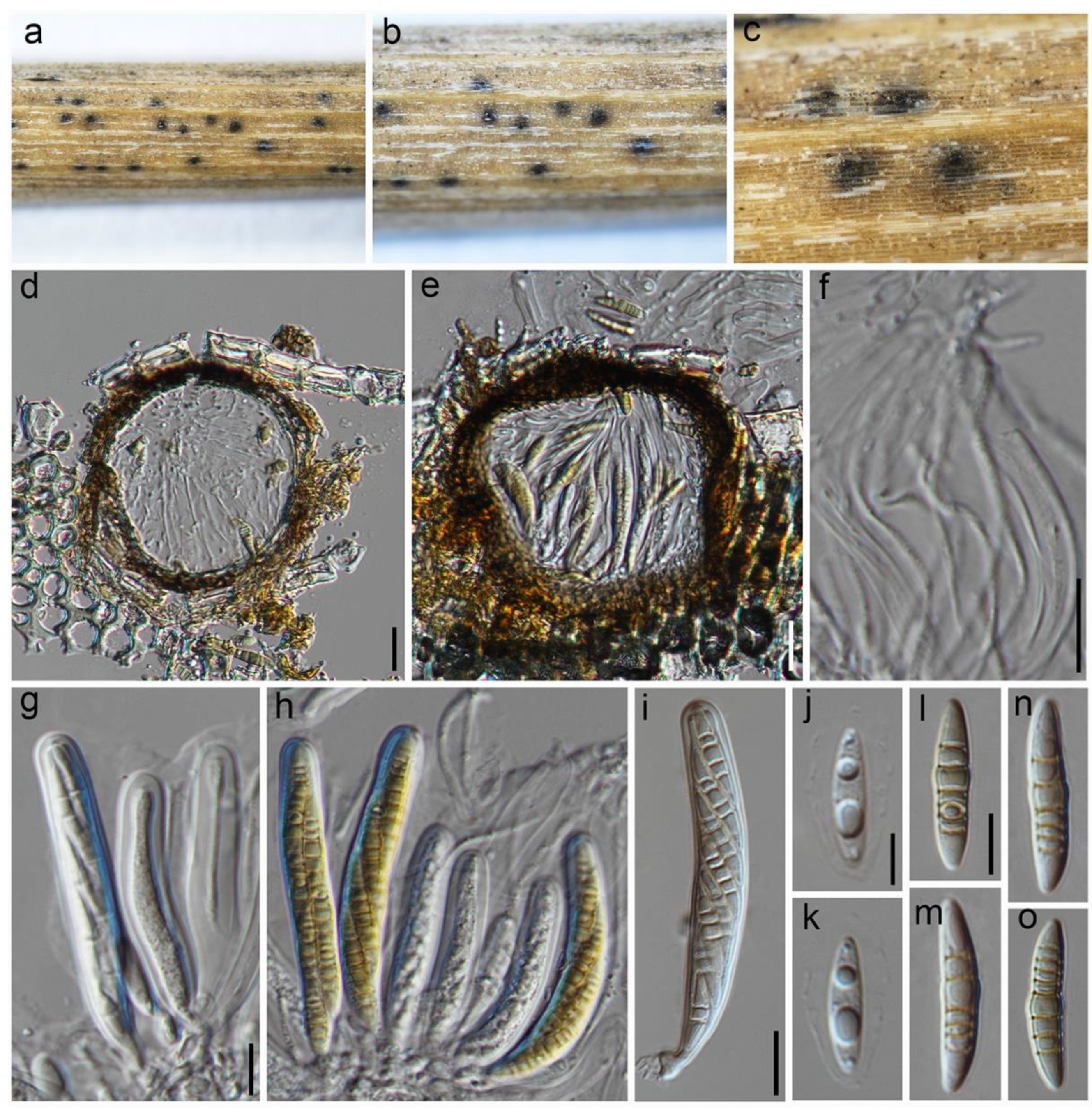
3.2. Taxonomy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Phookamsak, R.; Wanasinghe, D.N.; Hongsanan, S.; Phukhamsakda, C.; Huang, S.K.; Tennakoon, D.S.; Norphanphoun, C.; Camporesi, E.; Bulgakov, T.S.; Promputtha, I.; et al. Towards a natural classification of Ophiobolus and ophiobolus-like taxa; introducing three novel genera Ophiobolopsis, Paraophiobolus and Pseudoophiobolus in Phaeosphaeriaceae (Pleosporales). Fungal Divers. 2017, 87, 299–339. [Google Scholar] [CrossRef]
- Index Fungorum. Available online: http://www.indexfungorum.org/Names/Names.asp (accessed on 28 June 2023).
- Wijayawardene, N.N.; Hyde, K.D.; Dai, D.Q.; Sánchez-García, M.; Goto, B.T.; Saxena, R.K.; Erdoğdu, M.; Selçuk, F.; Rajeshkumar, K.C.; Aptroot, A.; et al. Outline of Fungi and fungus-like taxa—2021. Mycosphere 2022, 13, 53–453. [Google Scholar] [CrossRef]
- Phookamsak, R.; Liu, J.-K.; McKenzie, E.H.C.; Manamgoda, D.S.; Ariyawansa, H.; Thambugala, K.M.; Dai, D.-Q.; Camporesi, E.; Chukeatirote, E.; Wijayawardene, N.N.; et al. Revision of Phaeosphaeriaceae. Fungal Divers. 2014, 68, 159–238. [Google Scholar] [CrossRef]
- Liu, J.-K.; Hyde, K.D.; Jeewon, R.; Phillips, A.; Maharachchikumbura, S.S.N.; Ryberg, M.; Liu, Z.-Y.; Zhao, Q. Ranking higher taxa using divergence times: A case study in Dothideomycetes. Fungal Divers. 2017, 84, 75–99. [Google Scholar] [CrossRef]
- El-Demerdash, A.; El-Demerdash, A. Chemical diversity and biological activities of Phaeosphaeria fungi genus: A systematic review. J. Fungi 2018, 4, 130. [Google Scholar] [CrossRef] [PubMed]
- Norouzi, H.; Sohrabi, M.; Yousefi, M.; Boustie, J. Tridepsides as potential bioactives: A review on their chemistry and the global distribution of their lichenic and non-lichenic natural sources. Front. Fungal Biol. 2023, 4, 1088966. [Google Scholar] [CrossRef]
- Ashrafi, S.; Wennrich, J.P.; Becker, Y.; Maciá-Vicente, J.G.; Brißke-Rode, A.; Daub, M.; Thünen, T.; Dababat, A.A.; Finckh, M.R.; Stadler, M. Polydomus karssenii gen nov. sp. nov. is a dark septate endophyte with a bifunctional lifestyle parasitising eggs of plant parasitic cyst nematodes (Heterodera spp.). IMA Fungus 2023, 14, 6. [Google Scholar]
- Wanasinghe, D.N.; Mortimer, P.E.; Xu, J. Insight into the systematics of microfungi colonizing dead woody twigs of Dodonaea viscosa in Honghe (China). J. Fungi 2021, 7, 180. [Google Scholar] [CrossRef]
- MycoBank. Available online: http://www.mycobank.org (accessed on 28 June 2023).
- Wanasinghe, D.N.; Wijayawardene, N.N.; Xu, J.C.; Cheewangkoon, R.; Mortimer, P.E. Taxonomic novelties in Magnolia-associated pleosporalean fungi in the Kunming Botanical Gardens (Yunnan, China). PLoS ONE 2020, 15, e0235855. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Appl. 1990, 18, 315–322. [Google Scholar]
- Rehner, S.A.; Samuels, G.J. Taxonomy and phylogeny of Gliocladium analysed from nuclear large subunit ribosomal DNA sequences. Mycol. Res. 1994, 98, 625–634. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- Rehner, S.A.; Buckley, E.A. Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes evidence from an RNA polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Sung, G.H.; Sung, J.M.; Hywel-Jones, N.L.; Spatafora, J.W. A multi-gene phylogeny of Clavicipitaceae (Ascomycota, Fungi): Identification of localized incongruence using a combinational bootstrap approach. Mol. Phylogenet. Evol. 2007, 44, 1204–1223. [Google Scholar] [CrossRef]
- Wanasinghe, D.N.; Mortimer, P.E. Taxonomic and phylogenetic insights into novel Ascomycota from forest woody Litter. Biology 2022, 11, 889. [Google Scholar] [CrossRef]
- Hyde, K.D.; Tennakoon, D.S.; Jeewon, R.; Bhat, D.J.; Maharachchikumbura, S.S.N.; Rossi, W.; Leonardi, M.; Lee, H.B.; Mun, H.Y.; Houbraken, J.; et al. Fungal diversity notes 1036–1150: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2019, 96, 1–242. [Google Scholar] [CrossRef]
- Tennakoon, D.S.; Thambugala, K.M.; Wanasinghe, D.N.; Gentekaki, E.; Promputtha, I.; Kuo, C.H.; Hyde, K.D. Additions to Phaeosphaeriaceae (Pleosporales): Elongaticollum gen. nov., Ophiosphaerella taiwanensis sp. nov., Phaeosphaeriopsis beaucarneae sp. nov. and a new host record of Neosetophoma poaceicola from Musaceae. MycoKeys 2020, 70, 59–88. [Google Scholar] [CrossRef]
- Devadatha, B.; Mehta, N.; Wanasinghe, D.N.; Baghela, A.; Sarma, V.V. Vittaliana mangrovei, gen. nov., sp. nov. (Phaeosphaeriaceae) from mangroves near Pondicherry (India), based on morphology and multigene phylogeny. Cryptogam. Mycol. 2019, 40, 117–132. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Ariyawansa, H.A.; Wanasinghe, D.N.; Dayarathne, M.C.; Al-Saady, N.A.; Al-Sadi, A.M. Phylogenetic classification and generic delineation of Hydeomyces desertipleosporoides gen. et sp. nov., (Phaeosphaeriaceae) from Jebel Akhdar Mountain in Oman. Phytotaxa 2019, 391, 28–38. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Phylogenies and the comparative method. Am. Nat. 1985, 125, 1–15. [Google Scholar] [CrossRef]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree Version 1.4.0. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 15 June 2023).
- Crous, P.W.; Wingfield, M.J.; Richardson, D.M. Fungal Planet description sheets: 400–468. Persoonia 2016, 36, 316–458. [Google Scholar] [CrossRef]
- De Gruyter, J.; Aveskamp, M.M.; Woudenberg, J.H.; Verkley, G.J.; Groenewald, J.Z.; Crous, P.W. Molecular phylogeny of Phoma and allied anamorph genera: Towards a reclassification of the Phoma complex. Mycol. Res. 2009, 113, 508–519. [Google Scholar] [CrossRef]
- Leuchtmann, A. Über Phaeosphaeria Miyake und andere bitunicate Ascomyceten mit mehrfach querseptierten Ascosporen. Ph.D. Thesis, ETH Zurich, Zurich, Switzerland, 1984. [Google Scholar]
- Shoemaker, R.A.; Babcock, C.E. Phaeosphaeria. Can. J. Bot. 1989, 67, 1500–1599. [Google Scholar] [CrossRef]
- Magaña-Dueñas, V.; Cano-Lira, J.F.; Stchigel, A.M. New Dothideomycetes from freshwater habitats in Spain. J. Fungi 2021, 7, 1102. [Google Scholar] [CrossRef]
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; McKenzie, E.H.C.; Sarma, V.V.; Boonmee, S.; Lücking, R.; Bhat, D.J.; Liu, N.G. Refined families of Dothideomycetes: Dothideomycetidae and Pleosporomycetidae. Mycosphere 2020, 11, 1533–2107. [Google Scholar] [CrossRef]
- Crous, P.W.; Shivas, R.G.; Quaedvlieg, W.; van der Bank, M.; Zhang, Y.; Summerell, B.A.; Guarro, J.; Wingfield, M.J.; Wood, A.R.; Alfenas, A.C.; et al. Fungal Planet description sheets: 214–280. Persoonia 2014, 32, 184–306. [Google Scholar] [CrossRef] [PubMed]
- Quaedvlieg, W.; Verkley, G.; Shin, H.-D.; Barreto, R.; Alfenas, A.; Swart, W.; Groenewald, J.; Crous, P. Sizing up Septoria. Stud. Mycol. 2013, 75, 307–390. [Google Scholar] [CrossRef] [PubMed]
- Oudemans, C.A.J.A. Contributions à la flore mycologique des Pays-Bas. XIII. Ned. Kruidkd. Arch. Ser. 2 1889, 5, 454–519. [Google Scholar]
- Crous, P.W.; Carris, L.M.; Giraldo, A.; Groenewald, J.Z.; Hawksworth, D.L.; Hemández-Restrepo, M.; Jaklitsch, W.M.; Lebrun, M.H.; Schumacher, R.K.; Stielow, J.B.; et al. The Genera of Fungi-fixing the application of the type species of generic names-G2: Allantophomopsis, Latorua, Macrodiplodiopsis, Macrohilum, Milospium, Protostegia, Pyricularia, Robillarda, Rotula, Septoriella, Torula, and Wojnowicia. IMA Fungus 2015, 6, 163–198. [Google Scholar] [CrossRef] [PubMed]
- Marin-Felix, Y.; Hernández-Restrepo, M.; Iturrieta-González, I.; García, D.; Gené, J.; Groenewald, J.Z.; Cai, L.; Chen, Q.; Quaedvlieg, W.; Schumacher, R.K.; et al. Genera of phytopathogenic fungi: GOPHY 3. Stud. Mycol. 2019, 94, 1–124. [Google Scholar] [CrossRef]
- Li, W.J.; Bhat, D.J.; Camporesi, E.; Tian, Q.; Wijayawardene, N.N.; Dai, D.Q.; Phookamsak, R.; Chomnunti, P.; Bahkali, A.H.; Hyde, K.D. New asexual morph taxa in Phaeosphaeriaceae. Mycosphere 2015, 6, 681–708. [Google Scholar] [CrossRef]
- Thambugala, K.M.; Wanasinghe, D.N.; Phillips, A.J.L.; Camporesi, E.; Bulgakov, T.S.; Phukhamsakda, C.; Ariyawansa, H.A.; Goonasekara, I.D.; Phookamsak, R.; Dissanayake, A.; et al. Mycosphere notes 1–50: Grass (Poaceae) inhabiting Dothideomycetes. Mycosphere 2017, 8, 697–796. [Google Scholar] [CrossRef]
- Hyde, K.D.; Dong, Y.; Phookamsak, R.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Liu, N.G.; Abeywickrama, P.D.; Mapook, A.; Wei, D.P.; et al. Fungal diversity notes 1151–1276: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2020, 100, 5–277. [Google Scholar] [CrossRef]
- Dayarathne, M.C.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Devadatha, B.; Sarma, V.V.; Khongphinitbunjong, K.; Chomnunti, P.; Hyde, K.D. Morpho-molecular characterization of microfungi associated with marine based habitats. Mycosphere 2020, 11, 1–188. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Abreu, V.P.; Bazzicalupo, A.; Chethana, K.W.T.; Clericuzio, M.; Dayarathne, M.C.; Dissanayake, A.J.; Ekanayaka, A.H.; He, M.-Q.; et al. Fungal diversity notes 603–708: Taxonomic and phylogenetic notes on genera and species. Fungal Divers. 2017, 87, 1–235. [Google Scholar] [CrossRef]
- Ariyawansa, H.A.; Hyde, K.D.; Jayasiri, S.C.; Buyck, B.; Chethana, K.W.T.; Dai, D.Q.; Dai, Y.C.; Daranagama, D.A.; Jayawardena, R.S.; Lücking, R.; et al. Fungal diversity notes 111–252—Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2015, 75, 27–274. [Google Scholar] [CrossRef]
- Kohlmeyer, J.; Volkmann-Kohlmeyer, B. Atrotorquata and Loratospora: New ascomycete genera on Juncus roemerianus. Syst. Ascomycetum 1993, 12, 7–22. [Google Scholar]
- Wanasinghe, D.N.; Phukhamsakda, C.; Hyde, K.D.; Jeewon, R.; Lee, H.B.; Jones, G.E.B.; Tibpromma, S.; Tennakoon, D.S.; Dissanayake, A.J.; Jayasiri, S.C.; et al. Fungal diversity notes 709–839: Taxonomic and phylogenetic contributions to fungal taxa with an emphasis on fungi on Rosaceae. Fungal Divers. 2018, 89, 1–236. [Google Scholar] [CrossRef]
- Zhang, J.F.; Liu, J.K.; Jeewon, R.; Wanasinghe, D.N.; Liu, Z.Y. Fungi from Asian Karst formations III. Molecular and morphological characterization reveal new taxa in Phaeosphaeriaceae. Mycosphere 2019, 10, 202–220. [Google Scholar] [CrossRef]
- Tanaka, K.; Hirayama, K.; Yonezawa, H.; Sato, G.; Toriyabe, A.; Kudo, H.; Hashimoto, A.; Matsumura, M.; Harada, Y.; Kurihara, Y.; et al. Revision of the Massarineae (Pleosporales, Dothideomycetes). Stud. Mycol. 2015, 82, 75–136. [Google Scholar] [CrossRef] [PubMed]
- Tennakoon, D.S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; Camporesi, E.; Promputtha, I. Taxonomy and phylogeny of Juncaceicola gen. nov. (Phaeosphaeriaceae, Pleosporinae, Pleosporales). Cryptogam. Mycol. 2016, 37, 135–156. [Google Scholar] [CrossRef]
- Devadatha, B.; Jones, E.B.G.; Wanasinghe, D.N.; Bahkali, A.H.; Hyde, K.D. Characterization of novel estuarine Ascomycota based on taxonomic and phylogenetic evaluation. Bot. Mar. 2023, 66, 281–300. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain | GenBank Accession Numbers | ||||
|---|---|---|---|---|---|---|
| ITS | LSU | SSU | tef1 | rpb2 | ||
| Acericola italica | MFLUCC 13-0609 T | NR_156344 | NG_057143 | NG_063642 | - | - |
| Alloconiothyrium camelliae | NTUCC 17-032-1 | MT112294 | MT071221 | MT071270 | MT232967 | - |
| Alloconiothyrium camelliae | NTUCC 17-032-2 | MT112295 | MT071222 | MT071271 | MT232965 | - |
| Alloneottiosporina thailandica | MFLUCC 15-0576 T | NR_169715 | NG_073831 | NG_070332 | MT454002 | MT432196 |
| Amarenomyces ammophilae | CBS 114595 | KF766146 | GU301859 | GU296185 | GU349035 | GU371724 |
| Arezzomyces cytisi | MFLUCC 15-0649 T | NR_153533 | NG_057083 | NG_063584 | - | - |
| Banksiophoma australiensis | CBS 142163 T | KY979739 | KY979794 | - | - | - |
| Bhagirathimyces himalayensis | NFCCI 4580 T | NR_168816 | NG_068644 | NG_068414 | MT091965 | - |
| Bhatiellae rosae | MFLUCC 17-0664 T | NR_157505 | NG_058718 | NG_061286 | - | - |
| Brunneomurispora lonicerae | KUMCC 18-0157 T | NR_164299 | NG_070462 | NG_067697 | MK359064 | MK359079 |
| Brunneomurispora lonicerae | KUMCC 18-0158 | MK356373 | MK356347 | MK356361 | MK359065 | - |
| Camarosporioides phragmitis | MFLUCC 13-0365 T | NR_153925 | NG_058954 | KX572350 | KX572354 | - |
| Chaetosphaeronema clematidicola | MFLUCC 17-2151 T | MT310619 | MT214574 | NG_070660 | MT394633 | MT394694 |
| Chaetosphaeronema clematidis | MFLUCC 17-2147 | MT310620 | MT214575 | MT226688 | MT394634 | MT394695 |
| Chaetosphaeronema hispidulum | CBS 216.75 | KF251148 | KF251652 | EU754045 | - | GU371777 |
| Dematiopleospora cirsii | MFLUCC 13-0615 T | KX274243 | KX274250 | - | KX284708 | - |
| Dematiopleospora mariae | MFLU 16-0121 | MT310621 | MT214576 | MT226689 | MT394635 | - |
| Dematiopleospora mariae | MFLUCC 13-0612 T | KX274244 | KJ749653 | KJ749652 | KJ749655 | - |
| Dematiopleospora salsolae | MFLUCC 17-0828 T | NR_157514 | NG_059184 | NG_063679 | MG829201 | MG829254 |
| Didymocyrtis consimilis | CBS 129141 | KT383812 | KT383796 | - | - | - |
| Didymocyrtis ramalinae | Ertz 16399 | KT383838 | KT383802 | - | - | - |
| Diederichomyces caloplacae | CBS 129140 | JQ238635 | JQ238637 | - | - | - |
| Diederichomyces caloplacae | CBS 129338 | JQ238641 | JQ238643 | - | - | - |
| Diederichomyces ficuzzae | CBS 128019 | KP170647 | JQ238616 | - | - | - |
| Dlhawksworthia alliariae | MFLUCC 13-0070 T | NR_164244 | NG_069375 | KX494878 | - | KX507261 |
| Dlhawksworthia clematidicola | MFLUCC 14-0910 T | NR_164267 | NG_069556 | NG_063681 | MG829202 | - |
| Dlhawksworthia lonicera | MFLUCC 14-0955 T | NR_164268 | NG_069557 | NG_063682 | MG829203 | - |
| Edenia gomezpompae | CBS 124106 T | NR_156217 | NG_059202 | - | - | - |
| Edenia gomezpompae | LVPEI 3225 | KU578033 | - | - | - | - |
| Elongaticollum hedychii | MFLUCC 18-1638 | MT321796 | MT321810 | MT321803 | MT328753 | - |
| Elongaticollum hedychii | MFLUCC 17-2653 T | MT321797 | MT321811 | MT321804 | MT328754 | - |
| Elongaticollum hedychii | NCYUCC 19-0286 | MT321798 | MT321812 | MT321805 | MT328755 | - |
| Embarria clematidis | MFLUCC 14-0652 T | KT306949 | KT306953 | KT306956 | - | - |
| Embarria clematidis | MFLUCC 14-0976 | MG828871 | MG828987 | MG829099 | MG829194 | - |
| Equiseticola fusispora | MFLUCC 14-0522 T | NR_154010 | NG_059249 | NG_061238 | MG520895 | - |
| Galiicola baoshanensis | HKAS 102234 T | NR_164300 | NG_068613 | NG_067698 | MK359066 | - |
| Galiicola dactylidicola | MFLUCC 13-0738 T | NR_154501 | NG_067531 | NG_063564 | - | - |
| Galiicola pseudophaeosphaeria | MFLU 14-0524 T | KT326692 | KT326693 | - | MG520896 | - |
| Hydeomyces desertipleosporoides | SQUCC 15259 T | NR_164295 | NG_068287 | NG_067692 | MK290848 | - |
| Hydeomyces desertipleosporoides | SQUCC 15260 | MK290842 | MK290840 | MK290844 | MK290849 | - |
| Hydeomyces hydei | SQUCC 15290 T | NR_173311 | MW077157 | NG_074955 | MW075774 | MW276078 |
| Italica achilleae | MFLUCC 14-0959 T | NR_157518 | NG_059862 | NG_063683 | MG829204 | MG829255 |
| Italica heraclei | MFLU 18-1906 T | MT881676 | MT881653 | MT881671 | MT901290 | - |
| Jeremyomyces labinae | CBS 144617 T | NR_163362 | NG_070463 | - | - | MK442665 |
| Juncaceicola alpina | CBS 456.84 T | NR_145172 | NG_069801 | NG_063078 | - | - |
| Juncaceicola luzulae | MFLUCC 13-0780 T | KX449529 | KX449530 | KX449531 | MG520898 | - |
| Juncaceicola padellana | CBS 606.86 T | NR_145138 | NG_069810 | - | - | - |
| Kwanghwaensis miscanthi | FU31017 T | MK503817 | MK503823 | MK503829 | MT009126 | - |
| Leptospora clematidis | MFLUCC 17-2148 T | NR_170811 | MT214577 | MT226690 | MT394636 | MT394696 |
| Leptospora galii | KUMCC 15-0521 T | NR_168158 | NG_068778 | NG_068364 | MG520899 | - |
| Leptospora hydei | GZCC 19-0004 T | MK522507 | MK522497 | MK522503 | MK523387 | - |
| Longispora clematidis | MFLU 15–1277 T | MT310625 | MT214580 | MT226693 | MT394639 | - |
| Loratospora aestuarii | JK 5535B | MH863024 | GU301838 | GU296168 | - | GU371760 |
| Mauginiella scaettae | CBS 239.58 | MH857770 | MH869303 | - | - | - |
| Mauginiella scaettae | CBS 158.26 | MH854874 | MH866370 | - | - | - |
| Megacoelomyces sanchezii | CMHUB 24030 | MN429018 | MN429020 | MN429016 | - | MN546863 |
| Megacoelomyces sanchezii | CMHUB 24055 | MN429019 | MN429021 | MN429017 | - | - |
| Melnikia anthoxanthi | MFLUCC 14-1011 T | - | KU848204 | KU848205 | - | - |
| Murichromolaenicola chiangraiensis | MFLUCC 17-1488 T | MN994582 | MN994559 | MN994605 | MN998163 | - |
| Murichromolaenicola chromolaenae | MFLUCC 17-1489 T | MN994583 | MN994560 | MN994606 | MN998164 | - |
| Muriphaeosphaeria galatellae | MFLUCC 14-0614 T | NR_154189 | NG_059600 | NG_061216 | MG520900 | - |
| Muriphaeosphaeria galatellae | MFLUCC 15-0769 | - | KT438330 | KT438332 | - | - |
| Neoophiobolus chromolaenae | MFLUCC 17-1467 T | NR_168851 | NG_068693 | NG_070144 | MN998166 | - |
| Neoophiobolus chromolaenae | MFLUCC 17-1449 | MN994584 | MN994561 | MN994607 | MN998165 | - |
| Neosetophoma camporesii | MFLUCC 15-0682 T | KU302779 | KU302778 | NG_070320 | MN654114 | - |
| Neosetophoma garethjonesii | MFLUCC 14-0528 T | KY496758 | KY496738 | KY501126 | KY514402 | - |
| Neosetophoma rosae | MFLUCC 17-0844 | MG828926 | MG829035 | MG829141 | MG829219 | - |
| Neosetophoma shoemakeri | MFLUCC 17-2510 T | NR_161044 | NG_069548 | NG_065723 | MG739515 | - |
| Neosphaerellopsis thailandica | CPC 21659 T | NR_137954 | NG_067289 | - | - | - |
| Neostagonospora arrhenatheri | MFLUCC 15-0464 T | KX926417 | KX910091 | KX950402 | MG520901 | - |
| Neostagonospora caricis | CBS 135092 T | NR_156264 | NG_070457 | - | - | KF252171 |
| Neostagonospora elegiae | CBS 135101 T | NR_156265 | NG_070369 | - | - | KF252172 |
| Neostagonospora phragmitis | MFLUCC 16-0493 T | KX926416 | KX910090 | KX950401 | MG520902 | - |
| Neostagonospora sorghi | CBS 145366 T | NR_164452 | NG_070466 | - | - | MK540087 |
| Neostagonospora spinificis | R. Kirschner 3867 | KP676045 | KP676046 | - | - | - |
| Neostagonosporella bambusicola | KUMCC 20-0031 T | OP454331 | OP454335 | OP454058 | - | - |
| Neostagonosporella sichuanensis | MFLUCC 18-1228 T | MH368073 | MH368079 | MH368088 | MK313851 | - |
| Neostagonosporella sichuanensis | MFLU 18-1223 | MH394690 | MH394687 | MK296469 | MK313854 | - |
| Neosulcatispora agaves | CPC 26407 T | NR_154233 | NG_070394 | - | - | - |
| Nodulosphaeria aconiti | MFLUCC 13-0728 T | NR_154236 | KU708844 | KU708840 | KU708852 | KU708856 |
| Nodulosphaeria digitalis | MFLUCC 17-2418 | MG891749 | MG891750 | MH791042 | MH791041 | - |
| Nodulosphaeria guttulatum | MFLUCC 15-0069 | KY496746 | KY496726 | KY501115 | KY514394 | KY514405 |
| Nodulosphaeria scabiosae | MFLUCC 14-1111 T | NR_154237 | KU708846 | NG_063602 | KU708854 | KU708857 |
| Ophiobolopsis italica | MFLUCC 17-1791 T | NR_156678 | NG_059856 | NG_061281 | MG520903 | - |
| Ophiobolus disseminans | MFLUCC 17-1787 | MG520941 | MG520961 | MG520980 | MG520906 | - |
| Ophiobolus hydei | TASM 6143 | MK981301 | MK981305 | MK981303 | MK993651 | - |
| Ophiobolus hydei | TASM 6144 | MK981300 | MK981304 | MK981302 | MK993650 | - |
| Ophiosimulans tanaceti | MFLUCC 14-0525 | KU738890 | KU738891 | KU738892 | MG520910 | - |
| Ophiosphaerella taiwanensis | NCYUCC 19-0152 T | NR_171874 | MT321815 | MT321808 | MT328758 | - |
| Ophiosphaerella taiwanica | NTUCC 17-024 T | NR_171846 | MN082419 | - | MN199124 | - |
| Ophiosphaerella taiwanica | NTUCC 17-025 | MN082418 | MN082420 | - | MN199125 | - |
| Paraleptospora chromolaenae | MFLUCC 17-1481 T | NR_168852 | NG_068694 | NG_070145 | MN998167 | - |
| Paraleptospora chromolaenicola | MFLUCC 17-2670 T | NR_168853 | NG_068695 | NG_070146 | MN998168 | - |
| Paraloratospora breonadiae | CPC 25944 T | NR_155675 | NG_068582 | - | - | - |
| Paraloratospora camporesii | MFLU 18-0915 T | NR_170014 | NG_073784 | MN756635 | MN756633 | - |
| Paraloratospora fructigena | FMR 17808 T | OU612363 | OU612362 | - | OU600609 | OU600607 |
| Paraloratospora gahniae | CPC 32454 T | NR_156675 | NG_059852 | - | - | MG386148 |
| Paraloratospora marina | MFLUCC 19 0691 T | OQ130046 | OQ130110 | OQ130107 | OQ357219 | OQ162221 |
| Paraloratospora sichuanensis | KUNCC 23-14218T | OR206396 | OR206415 | OR206405 | OR195712 | OR195721 |
| Paraloratospora sichuanensis | HKAS 129218 | OR206397 | OR206416 | OR206406 | OR195713 | OR195722 |
| Paraophiobolus arundinis | MFLUCC 17-1789 T | NR_156680 | NG_069539 | NG_061282 | MG520912 | - |
| Paraophiobolus torilicola | MFLU 20-0392 | MT370411 | MT370428 | MT370372 | - | - |
| Paraphoma chrysanthemicola | CBS 522.66 | KF251166 | KF251670 | GQ387521 | - | KF252174 |
| Paraphoma ledniceana | CBS 146533 T | NR_172538 | MT371396 | - | - | - |
| Paraphoma radicina | CBS 111.79 T | NR_156556 | NG_070446 | EU754092 | - | KF252180 |
| Paraphoma rhaphiolepidis | CBS 142524 T | KY979758 | NG_070415 | - | - | KY979851 |
| Paraphoma vinacea | UMPV001 | KU176884 | KU176888 | - | - | - |
| Parastagonospora fusiformis | MFLUCC 13-0215 T | NR_165848 | NG_068235 | NG_068367 | - | KX863711 |
| Parastagonospora nodorum | CBS 259.49 T | KF251185 | KF251688 | - | GU456285 | KF252192 |
| Parastagonospora phragmitis | CPC 32075 T | NR_164454 | NG_066451 | - | - | MK540089 |
| Parastagonospora poaceicola | MFLUCC 15-0471 T | - | NG_068537 | NG_068368 | - | KX880499 |
| Parastagonosporella fallopiae | CBS 135981 T | NR_160062 | NG_067337 | - | - | MH460547 |
| Parastagonosporella fallopiae | CCTU 1151-1 | MH460544 | MH460546 | - | - | - |
| Phaeosphaeria acaciae | MFLU 17-0496 T | NR_160335 | NG_069453 | KY768870 | - | - |
| Phaeosphaeria acaciae | KUMCC 20-0214 | MW078431 | MW078444 | MW078482 | MW082602 | MW192765 |
| Phaeosphaeria ampeli | MFLUCC 18-1641 T | NR_165910 | MK503808 | MK503814 | MK503802 | - |
| Phaeosphaeria ampeli | MFLUCC 19-0150 | MK503798 | MK503809 | MK503815 | MK503803 | - |
| Phaeosphaeria avenaria f. sp. tritici | CBS 289.52 | MH857041 | MH868572 | - | - | KY090671 |
| Phaeosphaeria calamicola | MFLUCC 14-1168 T | KY511429 | KY511423 | KY511426 | - | - |
| Phaeosphaeria caricicola | CBS 603.86 | KF251182 | GQ387590 | GQ387529 | - | KF252189 |
| Phaeosphaeria caricis-sectae | CBS 146823 T | NR_173034 | MZ064470 | - | - | MZ078195 |
| Phaeosphaeria chengduensis | KUNCC 23-13570 | OR206391 | OR206410 | OR206400 | OR195707 | OR195716 |
| Phaeosphaeria chengduensis | KUNCC 23-13571 T | OR206392 | OR206411 | OR206401 | OR195708 | OR195717 |
| Phaeosphaeria chiangraina | MFLUCC 13-0231 T | NR_155643 | NG_069237 | KM434289 | KM434298 | KM434307 |
| Phaeosphaeria chinensis | MFLUCC 19-0217 T | MN173212 | MN173208 | MN173216 | - | - |
| Phaeosphaeria chinensis | KUMCC 19-0161 | MN173213 | MN173210 | MN173217 | - | - |
| Phaeosphaeria cycadis | KUMCC 18-0161 T | NR_164445 | NG_070078 | NG_067700 | MK359069 | - |
| Phaeosphaeria cycadis | KUMCC 18-0162 | MK356379 | MK356353 | MK356367 | MK359070 | - |
| Phaeosphaeria elongata | CBS 120250 | MH863080 | GU456327 | GU456306 | GU456261 | GU456345 |
| Phaeosphaeria eustoma | CBS 724.92 | MH862385 | JX681112 | - | - | - |
| Phaeosphaeria eustoma | CBS 307.71 | MH860138 | JX681111 | - | - | - |
| Phaeosphaeria glyceriae-plicatae | CBS 101261 | MH862724 | MH874330 | - | - | - |
| Phaeosphaeria juncophila | CBS 575.86 | AF439488 | GU456328 | GU456307 | GU456283 | - |
| Phaeosphaeria luctuosa | CBS 308.79 | MH861209 | GU301861 | - | GU349004 | KY090678 |
| Phaeosphaeria musae | MFLUCC 17-2648 | MK503796 | MK503807 | MK503813 | - | - |
| Phaeosphaeria musae | CBS 120026 | DQ885894 | GU301862 | GU296186 | GU349037 | GU357748 |
| Phaeosphaeria musae | MFLUCC 11-0151 | KM434268 | KM434278 | KM434288 | KM434297 | - |
| Phaeosphaeria musae | MFLUCC 11-0133 | KM434267 | KM434277 | KM434287 | KM434296 | - |
| Phaeosphaeria nodulispora | URM 7220 | KR092904 | KR092903 | - | - | - |
| Phaeosphaeria norfolcia | CBS 593.86 | MH861997 | MH873686 | - | - | - |
| Phaeosphaeria oryzae | CBS 110110 T | NR_156557 | NG_069025 | NG_061080 | - | KF252193 |
| Phaeosphaeria oryzae | MFLUCC 11-0170 | KM434269 | KM434279 | - | - | KM434306 |
| Phaeosphaeria papayae | S528 | KF251187 | KF251690 | - | - | - |
| Phaeosphaeria papayae | CBS 135416 | MH866082 | MH877574 | - | - | - |
| Phaeosphaeria parvula | CBS 605.86 | MH862001 | MH873689 | - | - | - |
| Phaeosphaeria parvula | CBS 260.49 | MH856516 | MH868046 | - | - | - |
| Phaeosphaeria penniseti | FU31020 | MK503819 | MK503825 | MK503831 | - | - |
| Phaeosphaeria phoenicicola | CPC 28711 T | NR_156608 | - | - | - | - |
| Phaeosphaeria poagena | CBS 136771 T | NR_168146 | NG_068518 | - | - | - |
| Phaeosphaeria poagena | KUNCC 23-13572 | OR206393 | OR206412 | OR206402 | OR195709 | OR195718 |
| Phaeosphaeria poagena | KUNCC 23-13573 | OR206394 | OR206413 | OR206403 | OR195710 | OR195719 |
| Phaeosphaeria podocarpi | CBS 138903 T | NR_137933 | NG_070060 | - | - | - |
| Phaeosphaeria sichuanensis | KUNCC 23-13568 | OR206389 | OR206408 | OR206398 | OR195705 | OR195714 |
| Phaeosphaeria sichuanensis | KUNCC 23-13569 T | OR206390 | OR206409 | OR206399 | OR195706 | OR195715 |
| Phaeosphaeria sinensis | MFLUCC 18-1552 T | NR_163350 | NG_070076 | NG_065788 | MK360072 | - |
| Phaeosphaeria sinensis | KUMCC 17-0195 | OM212456 | OL813496 | OL824792 | ON203111 | - |
| Phaeosphaeria sinensis | NCYUCC 19-0369 | MN937237 | MN937219 | - | - | - |
| Phaeosphaeria thysanolaenicola | MFLUCC 10-0563 T | NR_155642 | NG_069236 | KM434286 | KM434295 | KM434303 |
| Phaeosphaeriopsis aloes | HKAS 115783 | MZ493305 | MZ493319 | MZ493291 | MZ508414 | - |
| Phaeosphaeriopsis beaucarneae | MFLU 18-2586 T | NR_170822 | MT321813 | MT321806 | MT328756 | - |
| Phaeosphaeriopsis beaucarneae | MFLU 18-2587 | MT321800 | MT321814 | MT321807 | MT328757 | - |
| Phaeosphaeriopsis glaucopunctata | MFLUCC 13-0265 | KJ522473 | KJ522477 | KJ522481 | MG520918 | - |
| Phaeosphaeriopsis triseptata | MFLUCC 13-0271 | KJ522475 | KJ522479 | KJ522484 | MG520919 | KJ522485 |
| Phaeosphaeriopsis yuccae | MFLUCC 16-0558 | KY554482 | KY554481 | KY554480 | MG520920 | - |
| Piniphoma wesendahlina | CBS 145032 T | NR_163375 | NG_070464 | - | - | MK442676 |
| Populocrescentia ammophilae | MFLUCC 17-0665 T | NR_157535 | NG_059875 | NG_063690 | MG829231 | - |
| Populocrescentia forlicesenensis | MFLUCC 14-0651 T | NR_154326 | KT306952 | KT306955 | MG520925 | - |
| Populocrescentia rosae | TASM 6125 T | - | MG829060 | NG_062440 | MG829232 | - |
| Pseudoophiobolus galii | MFLUCC 17-2257 T | MG520947 | NG_069541 | NG_063667 | MG520926 | - |
| Pseudoophiobolus italicus | MFLUCC 17-2255 T | NR_156683 | NG_059858 | NG_063668 | MG520927 | - |
| Pseudoophiobolus mathieui | MFLUCC 17-1785 | MG520951 | MG520971 | MG520992 | MG520929 | - |
| Pseudoophiobolus urticicola | KUMCC 17-0168 T | NR_156686 | NG_069543 | NG_065140 | MG520933 | - |
| Pseudoophiosphaerella huishuiensis | HS13 | MK522509 | MK522499 | MK522505 | MK523389 | - |
| Pseudoophiosphaerella huishuiensis | MFLUCC 17-1453 | MN994590 | MN994567 | MN994613 | MN998171 | - |
| Pseudoophiosphaerella huishuiensis | MFLUCC 17-1463 | MN994591 | MN994568 | MN994614 | MN998172 | - |
| Pseudophaeosphaeria rubi | MFLUCC 14-0259 T | NR_154351 | NG_067543 | NG_061255 | MG520934 | - |
| Pseudostaurosphaeria chromolaenae | MFLUCC 17-1490 T | NR_168854 | NG_068696 | NG_070147 | MN998174 | - |
| Pseudostaurosphaeria chromolaenicola | MFLUCC 17-1491 | MN994594 | MN994571 | MN994617 | MN998175 | - |
| Pseudostaurosphaeria chromolaenicola | MFLUCC 17-1497 T | NR_168855 | NG_068697 | NG_070148 | MN998176 | - |
| Quixadomyces hongheensis | KUMCC 20-0215 T | NR_172441 | MW264194 | NG_074964 | MW256816 | MW269529 |
| Quixadomyces hongheensis | HKAS112346 | MW541826 | MW541822 | MW541833 | MW556134 | MW556136 |
| Sclerostagonospora cycadis | CBS 123538 T | NR_160231 | FJ372410 | - | - | - |
| Sclerostagonospora elegiae | CBS 118142 T | NR_176098 | NG_081272 | - | - | - |
| Sclerostagonospora ericae | CBS 141318 T | NR_145199 | NG_070625 | - | - | - |
| Sclerostagonospora fusiformis | CBS 118152 | JX517283 | JX517292 | - | - | - |
| Sclerostagonospora lathyri | MFLUCC 14-0958 T | NR_158956 | NG_069566 | NG_063692 | MG829235 | - |
| Sclerostagonospora pinguis | CBS 118146 T | NR_176097 | NG_081271 | - | - | - |
| Sclerostagonospora rosae | MFLU 18-0115 T | NR_157541 | NG_069567 | NG_065151 | MG829236 | - |
| Sclerostagonospora rosicola | MFLUCC 15-0129 T | MG828957 | MG829068 | NG_063693 | MG829237 | - |
| Scolicosporium minkeviciusii | MFLUCC 12-0089 | - | KF366382 | KF366383 | - | - |
| Septoriella agrostina | MFLU 18-0113 T | NR_157533 | NG_069561 | NG_062198 | MG829227 | - |
| Septoriella allojunci | MFLU 15-0701 T | KU058718 | KU058728 | NG_065141 | MG520935 | - |
| Septoriella ammophilicola | MFLU 17-2571 T | MN047087 | NG_070468 | NG_068403 | MN077065 | - |
| Septoriella ammophilicola | MFLU 17-2568 | MN047088 | MN017848 | MN017914 | MN077066 | - |
| Septoriella artemisiae | MFLUCC 17-0693 T | MG828929 | MG829038 | NG_063688 | - | - |
| Septoriella arundinicola | MFLU 16-0225 T | MG828946 | MG829056 | NG_062199 | MG829228 | MG829261 |
| Septoriella arundinis | MFLUCC 15-0702 T | KU058716 | KU058726 | NG_061283 | MG520921 | - |
| Septoriella asparagicola | MFLUCC 16-0379 T | NR_165908 | NG_070081 | NG_067708 | MK443385 | MK443387 |
| Septoriella bromi | MFLUCC 13-0739 T | KU058717 | KU058727 | - | - | - |
| Septoriella callistemonis | CBS 146822 T | NR_173033 | MZ064469 | - | - | MZ078194 |
| Septoriella camporesii | KUMCC 16-0113 T | NR_168233 | MN648201 | NG_068422 | - | - |
| Septoriella chlamydospora | MFLUCC 15-0177 T | NR_154508 | KU163654 | NG_063595 | - | - |
| Septoriella dactylidicola | MFLUCC 14-0002 T | - | KY657264 | KY657265 | - | - |
| Septoriella dactylidis | MFLU 15-2720 T | NR_154507 | KU163656 | - | - | - |
| Septoriella elongata | HA 4444 | KM491546 | KM491548 | KM491549 | - | - |
| Septoriella festucae | MFLUCC 17-0056 T | KY824766 | KY824767 | KY824769 | - | KY824768 |
| Septoriella forlicesenica | MFLUCC 15-0470 | KX926422 | KX910095 | KX950406 | MG520922 | KY131966 |
| Septoriella garethjonesii | MFLUCC 15-0469 | KX926425 | KX954390 | KY205717 | MG520923 | KX898363 |
| Septoriella garethjonesii | MFLUCC 13-0275 | KX926420 | KX910093 | KX950405 | - | KX880500 |
| Septoriella germanica | CBS 145372 T | NR_164459 | MK540035 | - | - | MK540096 |
| Septoriella hibernica | CBS 145371 T | NR_164460 | MK540036 | - | - | MK540097 |
| Septoriella hirta | CBS 536.77 T | NR_145192 | KR873278 | - | - | KR873324 |
| Septoriella hollandica | CBS 145374 T | NR_164461 | MK540037 | - | - | MK540098 |
| Septoriella hubertusii | CBS 338.86 T | NR_155786 | KF251733 | - | - | KF252235 |
| Septoriella italica | MFLUCC 13-0267 | KX926421 | KX910094 | KX950409 | MG520924 | KX891169 |
| Septoriella leuchtmannii | CBS 459.84 T | NR_163526 | NG_057999 | KY090700 | - | KF252195 |
| Septoriella luzulae | MFLUCC 14-0826 T | NR_154121 | NG_069310 | NG_063585 | - | - |
| Septoriella muriformis | MFLUCC 13-0277 | KX926415 | KX910089 | KX950400 | - | KX863710 |
| Septoriella neoarundinis | MFLUCC 15-0046 | KY706140 | KY706130 | KY706135 | KY706144 | KY706147 |
| Septoriella neoarundinis | MFLUCC 15-0027 T | NR_154541 | KY706129 | NG_063636 | MG520936 | - |
| Septoriella neodactylidis | MFLUCC 14-0966 T | NR_157511 | NG_069554 | NG_061288 | MG829199 | MG829253 |
| Septoriella neodactylidis | MFLUCC 13-0618 | KP744432 | KP744473 | KP753946 | - | - |
| Septoriella neomuriformis | MFLUCC 17-0372 T | MF611637 | MF611638 | MF611639 | - | - |
| Septoriella oudemansii | CBS 138012 T | KR873250 | KJ869224 | - | - | - |
| Septoriella paradactylidis | KUMCC 18-0154 | MK356371 | MK356345 | MK356359 | - | - |
| Septoriella paradactylidis | MFLU 17-0498 T | NR_164251 | NG_070411 | - | - | - |
| Septoriella paradactylidis | KUNCC 23-14219 | OR206395 | OR206414 | OR206404 | OR195711 | OR195720 |
| Septoriella phragmitis | CPC 24118 T | NR_132926 | NG_069285 | - | - | - |
| Septoriella poae | CBS 136766 T | NR_155793 | NG_067494 | - | - | - |
| Septoriella pseudophragmitis | CBS 145417 T | NR_164468 | MK560160 | - | - | MK559450 |
| Septoriella rosae | MFLU 18-0114 T | NR_157534 | NG_069562 | NG_065150 | MG829230 | - |
| Septoriella shoemakeri | MFLUCC 14-0963 T | NR_157512 | MG829003 | NG_063678 | MG829200 | - |
| Septoriella subcylindrospora | MFLUCC 13-0380 T | KT314184 | KT314183 | KT314185 | - | - |
| Septoriella tridentina | MFLUCC 15-0474 T | NR_165849 | NG_069389 | NG_068369 | - | KX891170 |
| Septoriella vagans | CBS 604.86 | KF251193 | KF251696 | - | - | KF252200 |
| Septoriella verrucispora | MFLUCC 19-0163 T | MK522508 | MK522498 | MK522504 | MK523388 | - |
| Setomelanomma holmii | CBS 110217 | KT389542 | GU301871 | GU296196 | GU349028 | GU371800 |
| Setophoma caverna | R150 | MK511944 | MK511965 | - | - | - |
| Setophoma sacchari | CBS 333.39 T | NR_145173 | NG_057837 | NG_062779 | - | KF252250 |
| Setophoma sacchari | MFLUCC 12-0241 | KJ476145 | KJ476147 | KJ476149 | KJ461318 | - |
| Setophoma vernoniae | CBS 137988 T | NR_168153 | KJ869198 | - | - | - |
| Stagonospora foliicola | CBS 110111 | KF251256 | KF251759 | EU754118 | - | - |
| Sulcispora pleurospora | CBS 460.84 | AF439498 | - | - | - | - |
| Sulcispora supratumida | MFLUCC 14-0995 T | NR_160325 | NG_067530 | NG_065627 | MH665366 | - |
| Tintelnotia destructans | CBS 127737 T | NR_147684 | NG_058274 | NG_063077 | - | KY090683 |
| Tintelnotia opuntiae | CBS 376.91 T | NR_147683 | NG_067267 | NG_062790 | - | KY090680 |
| Vittaliana mangrovei | NFCCI 4251 T | NR_165870 | NG_067809 | NG_067664 | MG767314 | MG767315 |
| Vrystaatia aloeicola | CBS 135107 | KF251278 | KF251781 | - | - | KF252283 |
| Wingfieldomyces cyperi | CBS 141450 T | NR_155805 | NG_059684 | - | - | MK540101 |
| Wojnowiciella cissampeli | CPC 27455 T | NR_155972 | NG_069358 | - | LT990616 | - |
| Wojnowiciella clematidis | MFLUCC 17-2159 T | NR_170812 | MT214582 | MT226695 | MT394641 | MT394698 |
| Wojnowiciella eucalypti | CPC 25024 T | NR_137996 | NG_070629 | - | LT990617 | - |
| Wojnowiciella kunmingensis | KUMCC 18-0159 T | NR_164446 | NG_070079 | NG_067701 | MK359071 | MK359078 |
| Wojnowiciella leptocarpi | CBS 115684 T | NR_155973 | KX306800 | - | LT990615 | LT990646 |
| Wojnowiciella spartii | MFLUCC 13-0402 T | KU058719 | KU058729 | NG_063670 | MG520937 | - |
| Xenophaeosphaeria grewiae | CBS 138867 T | NR_137944 | NG_058124 | - | - | - |
| Xenophoma puncteliae | CBS 128022 | JQ238617 | JQ238619 | - | - | - |
| Xenoseptoria neosaccardoi | CBS 120.43 | KF251280 | KF251783 | - | - | KF252285 |
| Xenoseptoria neosaccardoi | CBS 128665 | KF251281 | KF251784 | - | - | KF252286 |
| Yunnanensis chromolaenae | MFLUCC 17-1486 | MN994596 | MN994573 | MN994619 | MN998177 | - |
| Yunnanensis chromolaenae | MFLUCC 17-1487 T | NR_168856 | NG_068698 | NG_070149 | MN998178 | - |
| Yunnanensis phragmitis | MFLUCC 17-0315 T | MF684862 | MF684863 | MF684867 | MF683624 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wanasinghe, D.N.; Maharachchikumbura, S.S.N. Exploring the Diversity and Systematics of Phaeosphaeriaceae: Taxonomic Novelties from Ecologically Diverse Habitats and Their Phylogenetic Resolution. J. Fungi 2023, 9, 853. https://doi.org/10.3390/jof9080853
Wanasinghe DN, Maharachchikumbura SSN. Exploring the Diversity and Systematics of Phaeosphaeriaceae: Taxonomic Novelties from Ecologically Diverse Habitats and Their Phylogenetic Resolution. Journal of Fungi. 2023; 9(8):853. https://doi.org/10.3390/jof9080853
Chicago/Turabian StyleWanasinghe, Dhanushka N., and Sajeewa S. N. Maharachchikumbura. 2023. "Exploring the Diversity and Systematics of Phaeosphaeriaceae: Taxonomic Novelties from Ecologically Diverse Habitats and Their Phylogenetic Resolution" Journal of Fungi 9, no. 8: 853. https://doi.org/10.3390/jof9080853
APA StyleWanasinghe, D. N., & Maharachchikumbura, S. S. N. (2023). Exploring the Diversity and Systematics of Phaeosphaeriaceae: Taxonomic Novelties from Ecologically Diverse Habitats and Their Phylogenetic Resolution. Journal of Fungi, 9(8), 853. https://doi.org/10.3390/jof9080853