A New Model for Thermodynamic Characterization of Hemoglobin



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Thermodynamic Considerations
3. Hemoglobe Model
4. Physical-Mathematics Deductions
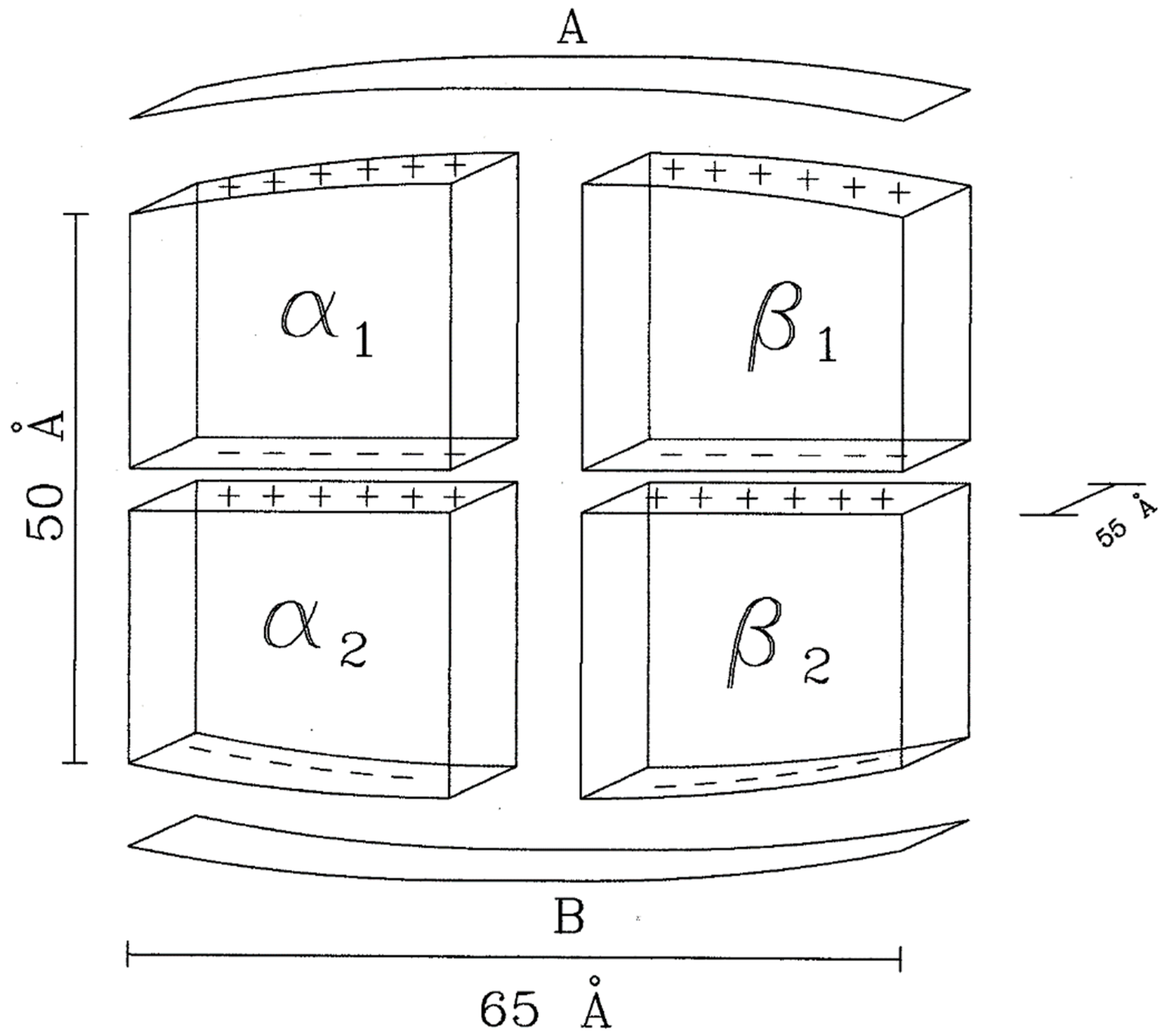
5. Hemoglobin as a Capacitor
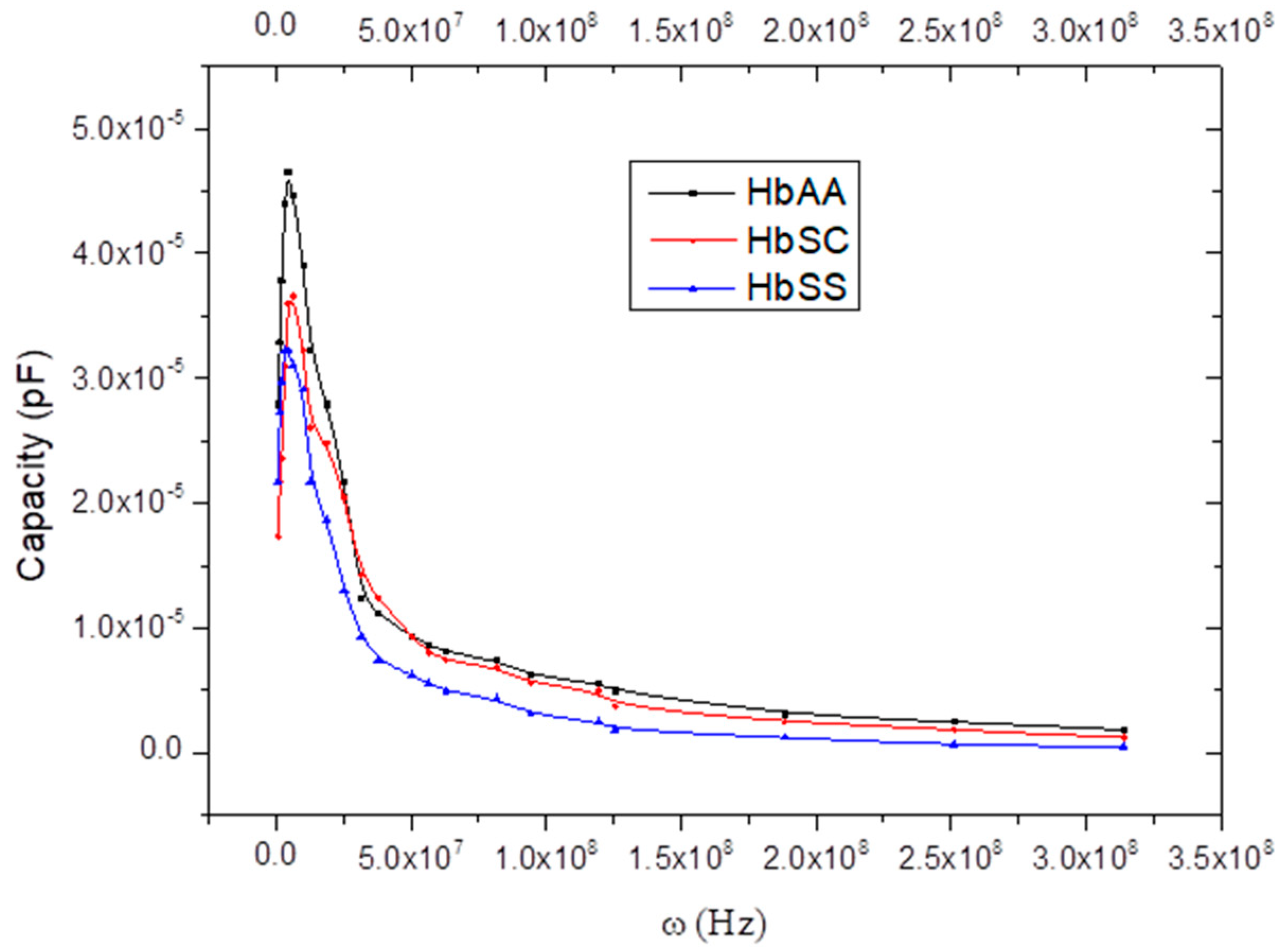
5.1. Observations
5.2. Capacitor Model
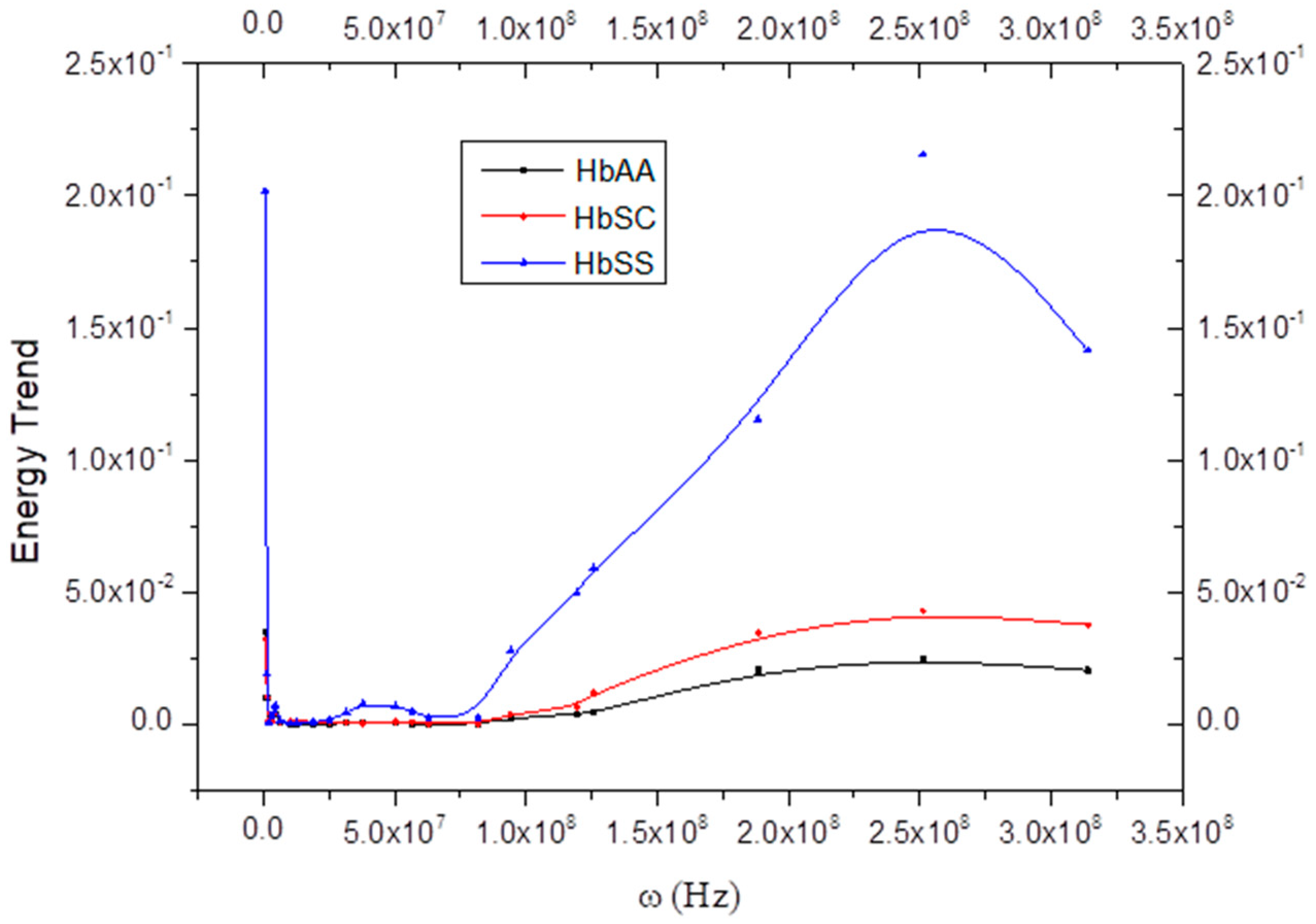
5.3. Hemoglobe Energy In Vitro
5.4. Hemoglobe Energy In Vivo
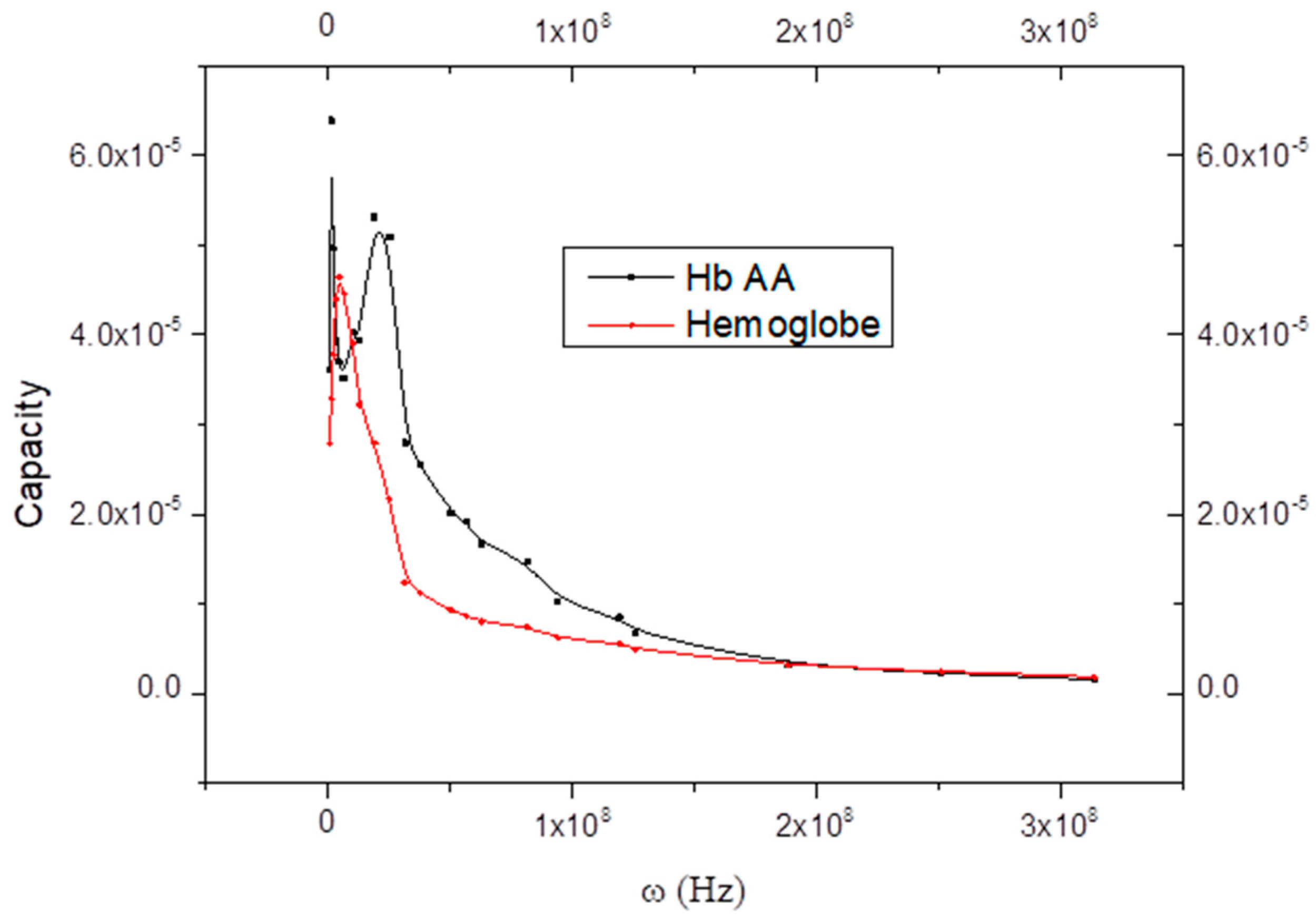
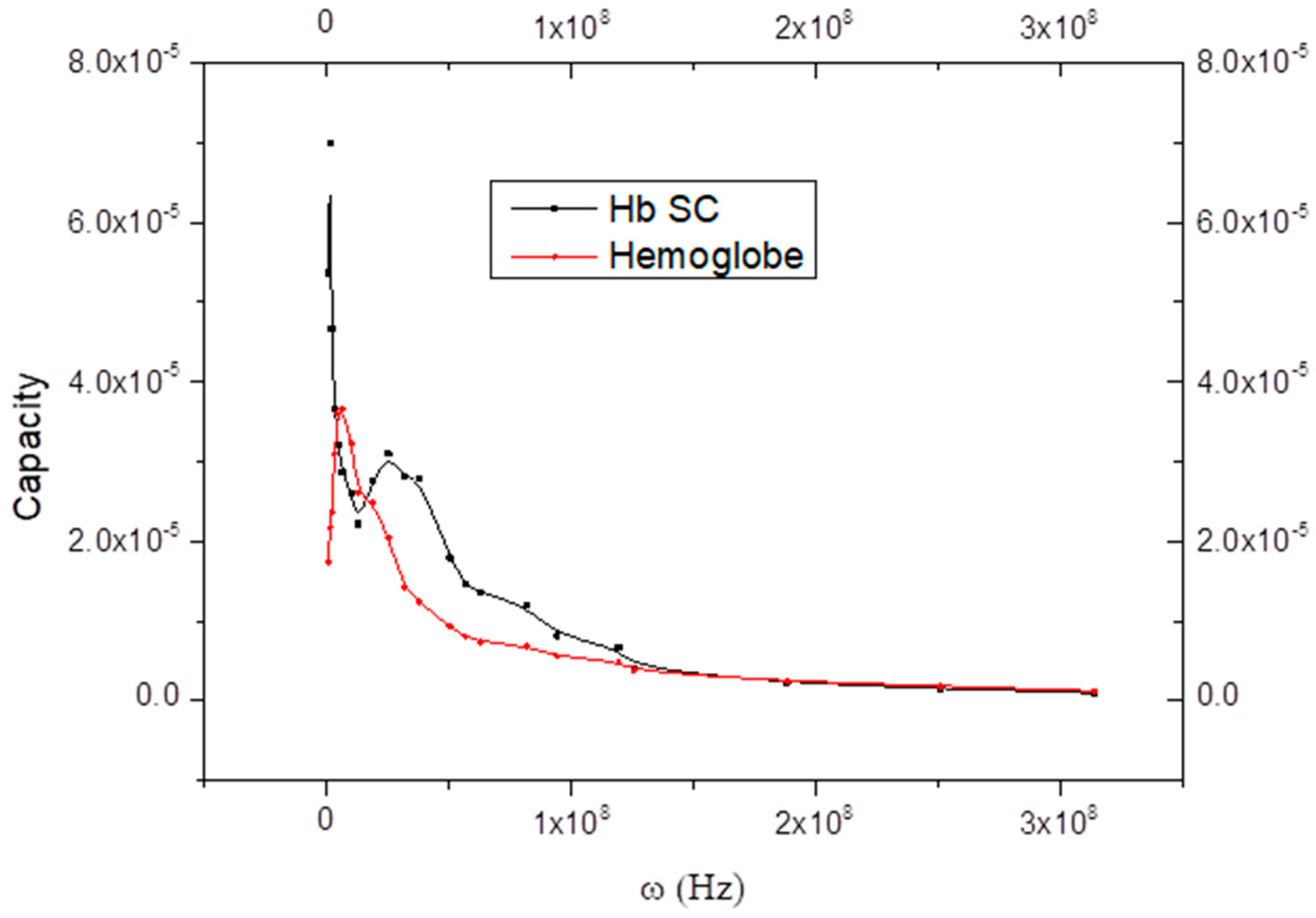
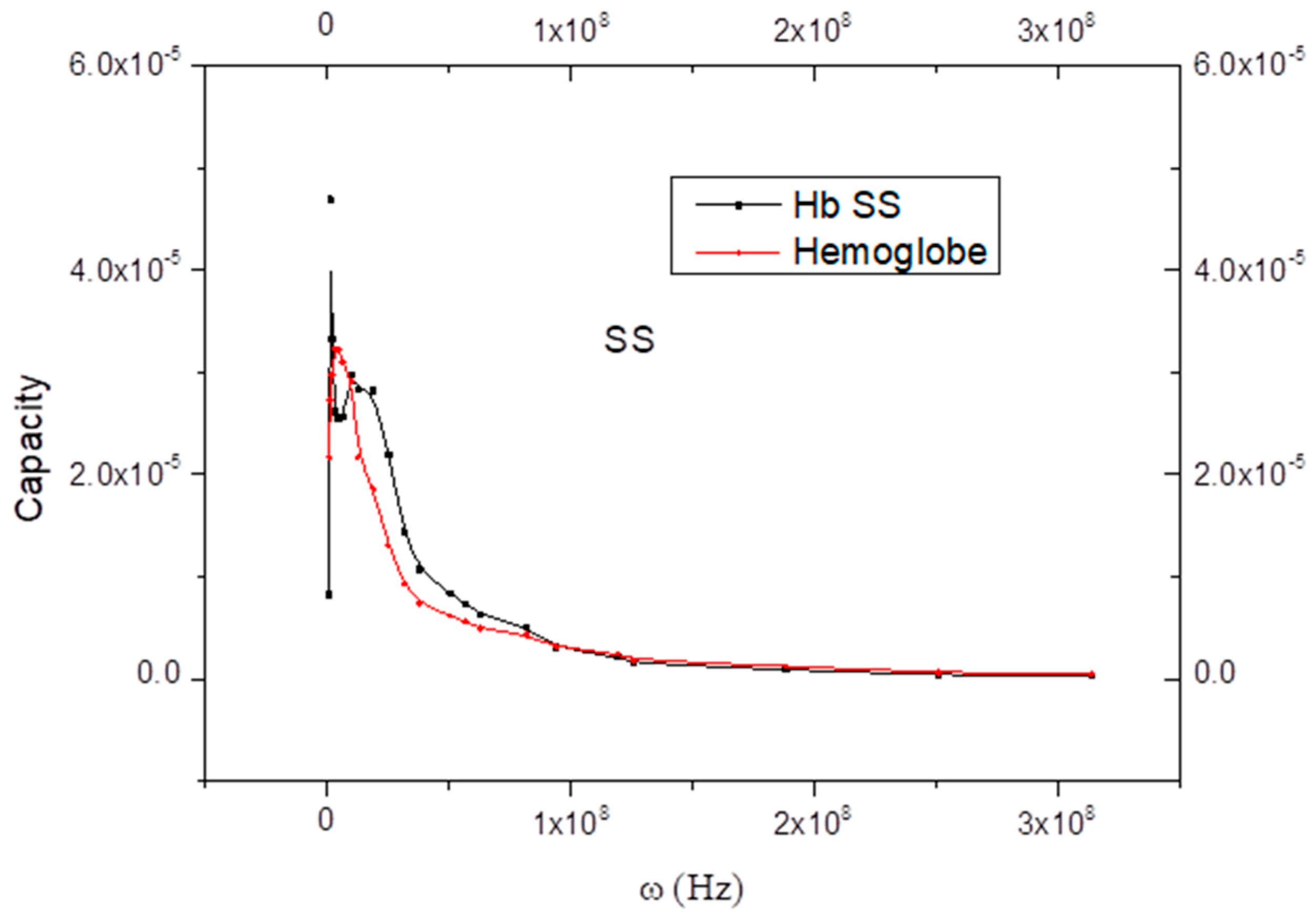
6. Curve Description
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Appleby, C.A. The origin and function of haemoglobin in plants. Sci. Prog. 1994, 76, 365–398. [Google Scholar]
- Van-Holde, K.K.E.; Miller, K.I.; Decker, H. Hemocyanins and invertebrate evolution. J. Biol. Chem. 2001, 276, 15563–15566. [Google Scholar] [CrossRef] [PubMed]
- Perutz, M.F. Haemoglobin as a model of an allosteric protein. Symp. Soc. Exp. Biol. 1973, 27, 1–3. [Google Scholar] [PubMed]
- Farsaci, F.; Tellone, E.; Galtieri, A.; Ficarra, S. A new model with internal variables for theoretical thermodynamic characterization of hemoglobin: Entropy determination and comparative study. J. Mol. Liq. 2019, 279, 632–639. [Google Scholar] [CrossRef]
- Farsaci, F.; Tellone, E.; Galtieri, A.; Ficarra, S. Is a dangerous blood clot formation a reversible process? Introduction of new characteristic parameter for thermodynamic clot blood characterization: Possible molecular mechanisms and pathophysiologic applications. J. Mol. Liq. 2018, 262, 345–353. [Google Scholar] [CrossRef]
- De Groot, S.R.; Mazur, P. Non Equilibrium Thermodynamics; Dover Publication: New York, NY, USA, 1984. [Google Scholar]
- Kluitenberg, G.A. On dielectric and magnetic relaxation phenomena and non-equilibrium thermodynamics. Physica 1973, 68, 75–82. [Google Scholar] [CrossRef]
- Kluitenberg, G.A. On dielectric and magnetic relaxation phenomena and vectorial internal degrees. Physica 1977, 87, 302–330. [Google Scholar] [CrossRef]
- Kluitenberg, G.A. On vectorial internal variables and dielectric and magnetic relaxation phenomena. Physica 1981, 109, 91–122. [Google Scholar] [CrossRef]
- Farsaci, F.; Tellone, E.; Galtieri, A.; Ficarra, S. Molecular characterization of a peculiar blood clot fluidification by theoretical thermodynamic models and entropy production study. J. Mol. Liq. 2018, 26, 457–462. [Google Scholar] [CrossRef]
- Farsaci, F.; Tellone, E.; Cavallaro, M.; Russo, A.; Ficarra, S. Low frequency dielectric characteristics of human blood: A non-equilibrium thermodynamic approach. J. Mol. Liq. 2013, 188, 113–119. [Google Scholar] [CrossRef]
- Farsaci, F.; Ficarra, S.; Russo, A.; Galtieri, A.; Tellone, E. Dielectric properties of human diabetic blood: Thermodynamic characterization and new prospective for alternative diagnostic techniques. J. Adv. Dielectr. 2015, 5, 3. [Google Scholar] [CrossRef]
- Farsaci, F.; Russo, A.; Ficarra, S.; Tellone, E. Dielectric properties of human normal and malignant liver tissue: A non-equilibrium thermodynamics approach. Open Access Libr. J. 2015, 2, e1395. [Google Scholar] [CrossRef]
- Farsaci, F.; Ficarra, S.; Russo, A.; Galtieri, A.; Tellone, E. On evaluation of electric conductivity by mean of non equilibriumthermodynamic approach with internal variables. An application to human erythrocyte suspension for metabolic characterizations. J. Mol. Liq. 2016, 224, 1181–1188. [Google Scholar] [CrossRef]
- Farsaci, F.; Tellone, E.; Galtieri, A.; Ficarra, S. Expanding the repertoire of dielectric fractional models: A comprehensive development and functional applications to predict metabolic alterations in experimentally-inaccessible cells or tissues. Fluids 2018, 3, 9. [Google Scholar] [CrossRef]
- Farsaci, F.; Tellone, E.; Galtieri, A.; Russo, A.; Ficarra, S. Evaluation of the human blood entropy production: A new thermodynamic approach. J. Ultrasound 2016, 19, 265–273. [Google Scholar] [CrossRef]
- Farsaci, F.; Tellone, E.; Russo, A.; Galtieri, A.; Ficarra, S. Rheological properties of human blood in the network of non-equilibrium thermodynamic with internal variables by means of ultrasound wave perturbation. J. Mol. Liq. 2017, 231, 206–212. [Google Scholar] [CrossRef]
- Farsaci, F.; Ficarra, S.; Galtieri, A.; Tellone, E. New non-equilibrium thermodynamic fractional visco-inelastic model to predict experimentally inaccessible processes and investigate pathophysiological cellular structures. Fluids 2017, 2, 59. [Google Scholar] [CrossRef]
- Laogun, A.A.; Agba, E.H.; Ajayi, N.O. A comparison of the dielectric behaviour of human haemoglobin SC with SS and AA in solution. Phys. Med. Biol. 1997, 42, 707–715. [Google Scholar] [CrossRef] [Green Version]
- Takashima, S. Use of protein database for the computation of the dipole moments of normal and abnormal hemoglobins. J. Biophys. Soc. 1993, 64, 1550–1558. [Google Scholar] [CrossRef] [Green Version]
- Antosiewicz, J.; Porschke, D. Electrostatics of Hemoglobins from Measurements of the Electric Dichroism and Computer Simulations. Biophys. J. 1995, 68, 655–664. [Google Scholar] [CrossRef]
- Huang, Y.X.; Wu, Z.J.; Huang, B.T.; Luo, M. Pathway and Mechanism of pH Dependent Human Hemoglobin Tetramer-Dimer-Monomer Dissociations. PLoS ONE 2013, 8, e81708. [Google Scholar] [CrossRef] [PubMed]
- Sommerfeld, A. Electrodynamics: Lectures on Theoretical Physics; Academic Press: Cambridge, MA, USA, 1956; Volume III. [Google Scholar]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farsaci, F.; Tellone, E.; Galtieri, A.; Ficarra, S. A New Model for Thermodynamic Characterization of Hemoglobin. Fluids 2019, 4, 135. https://doi.org/10.3390/fluids4030135
Farsaci F, Tellone E, Galtieri A, Ficarra S. A New Model for Thermodynamic Characterization of Hemoglobin. Fluids. 2019; 4(3):135. https://doi.org/10.3390/fluids4030135
Chicago/Turabian StyleFarsaci, Francesco, Ester Tellone, Antonio Galtieri, and Silvana Ficarra. 2019. "A New Model for Thermodynamic Characterization of Hemoglobin" Fluids 4, no. 3: 135. https://doi.org/10.3390/fluids4030135
APA StyleFarsaci, F., Tellone, E., Galtieri, A., & Ficarra, S. (2019). A New Model for Thermodynamic Characterization of Hemoglobin. Fluids, 4(3), 135. https://doi.org/10.3390/fluids4030135