
A Mathematical Model of Blood Loss during Renal Resection



Abstract
:1. Introduction
2. Materials and Methods
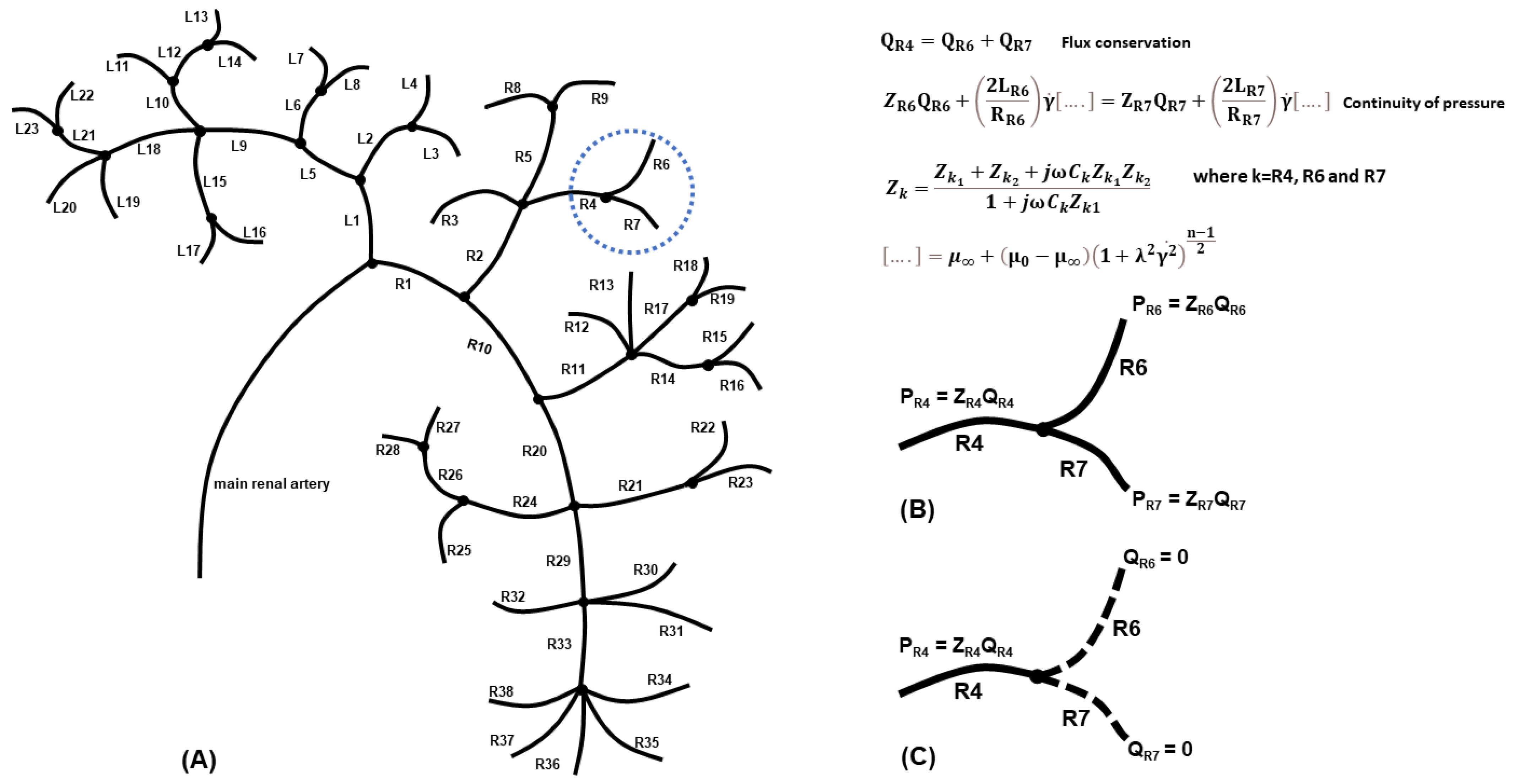
2.1. Generating the Renal Vasculature
2.2. Numerical Approximations
2.3. Effect of the Fluid Model Assumption on a Single-Node Asymmetric Bifurcation
2.4. Modelling Vessel Cuts
2.5. Model Validation and Verification
3. Results
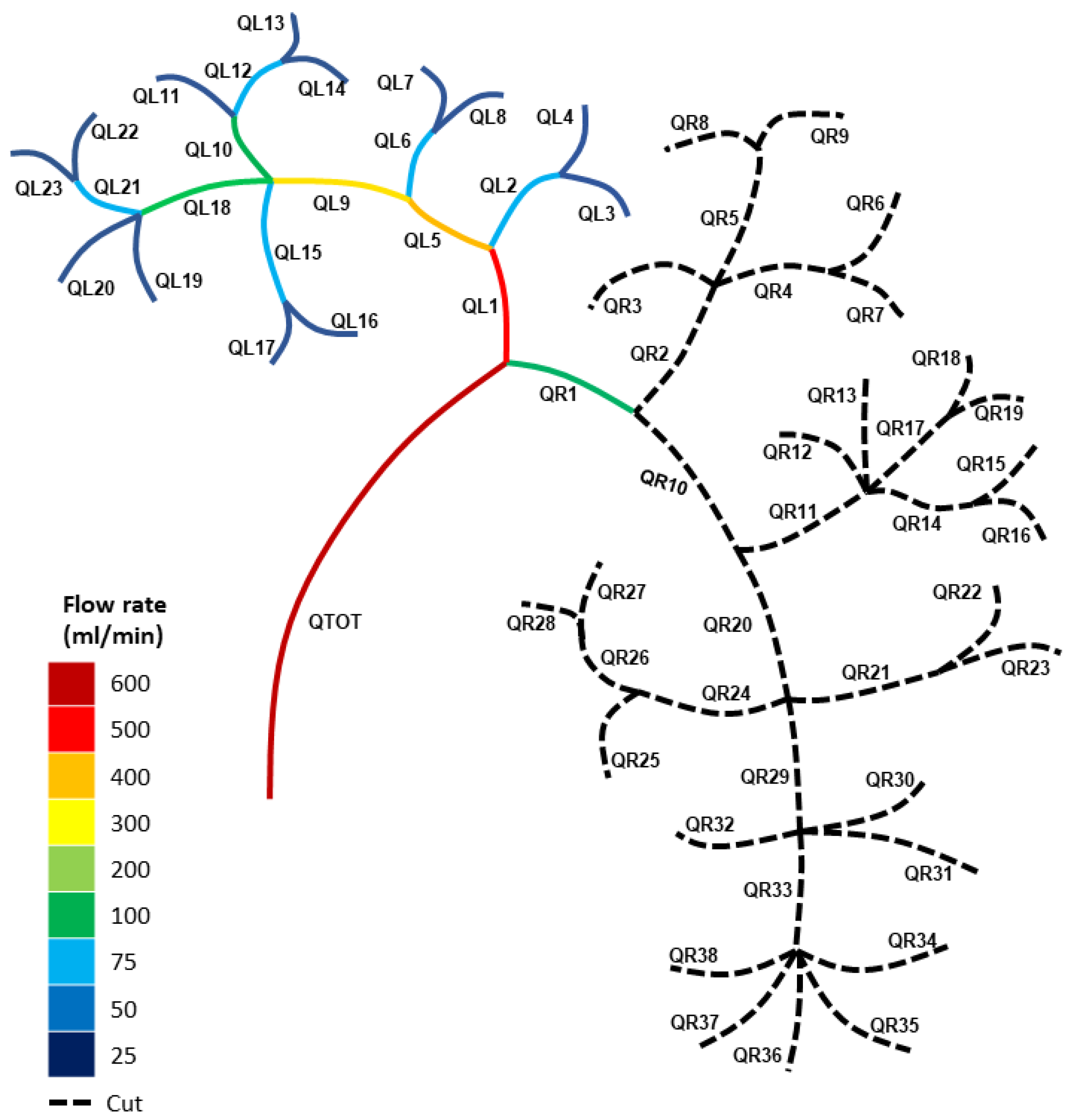
3.1. Modelling the Healthy (Uncut) Kidney Vascular Network
3.2. Modelling the Blood Loss for Single Cuts in the Kidney Vasculature
3.3. Modelling the Blood Loss for Double Cuts in the Kidney Vasculature
3.4. Blood Pressures for Multiple Cuts in the Kidney Vascular Network
4. Discussion
4.1. Clinical Relevance
4.2. Limitations
4.3. Future Work
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Stewart, G.D.; Klatte, T.; Cosmai, L.; Bex, A.; Lamb, B.W.; Moch, H.; Sala, E.; Siva, S.; Porta, C.; Gallieni, M. The multispeciality approach to the management of localised kidney cancer. Lancet 2022, 400, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Royal College of Surgeons of England. Surgery and the NHS in Numbers. Available online: https://www.rcseng.ac.uk/news-and-events/media-centre/media-background-briefings-and-statistics/surgery-and-the-nhs-in-numbers/ (accessed on 20 March 2023).
- NHS; Providers. NHS-Activity and Performance. Available online: http://nhsproviders.org (accessed on 20 March 2023).
- Dobbs, T.D.; Gibson, J.A.G.; Fowler, A.J.; Abbott, T.E.; Shahid, T.; Torabi, F.; Griffiths, R.; Lyons, R.A.; Pearse, R.M.; Whitaker, I.S. Surgical activity in England and Wales during the COVID-19 pandemic: A nationwide observational cohort study. Br. J. Anaesth. 2021, 127, 196–204. [Google Scholar] [CrossRef]
- NDRS. COVID-19 Rapid Cancer Registration and Treatment Data. 2022. Available online: https://digital.nhs.uk/ndrs/data (accessed on 20 March 2023).
- Abbott, T.E.F.; Fowler, A.J.; Dobbs, T.D.; Harrison, E.M.; Gillies, M.A.; Pearse, R.M. Frequency of surgical treatment and related hospital procedures in the UK: A national ecological study using hospital episode statistics. Br. J. Anaesth. 2017, 119, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Garnett, S.; Ahmed, S.; Sawyer, S. Laparoscopic Nephrectomoy; Department of Urology, Hailsham Urology Ward Eastbourne District General Hospital (UHS), Ed.; NHS: Sussex, UK, 2023. [Google Scholar]
- Hassouna, H.A.; Manikandan, R. Hemostasis in laparoscopic renal surgery. Indian J. Urol. 2012, 28, 3–8. [Google Scholar]
- Jaramillo, S.; Muntane, M.M.; Gambus, P.L.; Capitan, D.; Navarro-Ripoll, R.; Blasi, A. Perioperative blood loss: Estimation of blood volume loss or haemoglobin mass loss? Blood Transfus. 2020, 18, 20–29. [Google Scholar] [PubMed]
- Rosiello, G.; Larcher, A.; Fallara, G.; Basile, G.; Cignoli, D.; Colandrea, G.; Re, C.; Trevisani, F.; Karakiewicz, P.I.; Salonia, A.; et al. The impact of intraoperative bleeding on the risk of chronic kidney disease after nephron-sparing surgery. World J. Urol. 2021, 39, 2553–2558. [Google Scholar] [CrossRef]
- Puelles, V.G.; Combes, A.N.; Bertram, J.F. Clearly imaging and quantifying the kidney in 3D. Kidney Int. 2021, 100, 780–786. [Google Scholar] [CrossRef]
- Schutter, R.; Lantinga, V.A.; Hamelink, T.L.; Pool, M.B.F.; Varsseveld, O.C.v.; Potze, J.H.; Hillebrands, J.L.; Heuvel, M.C.v.d.; Dierckx, R.A.J.O.; Leuvenink, H.G.D.; et al. Magnetic resonance imaging assessment of renal flow distribution patterns during ex vivo normothermic machine perfusion in porcine and human kidneys. Transpl. Int. 2021, 34, 1643–1655. [Google Scholar] [CrossRef]
- Nordsletten, D.A.; Blackett, S.; Bentley, M.D.; Ritman, E.L.; Smith, N.P. Structural morphology of renal vasculature. Am. Physiol. Soc. 2006, 291, 296–309. [Google Scholar] [CrossRef]
- Cowley, J.; McGinty, S. A mathematical model of sonoporation using a liquid-crystalline shelled microbubble. Ultrasonics 2019, 96, 214–219. [Google Scholar] [CrossRef]
- Cowley, J.; Mulholland, A.J.; Gachagan, A. The Rayleigh-Plesset equation for a liquid-crystalline shelled microbubble. Int. J. Mod. Eng. Res. IJMER 2020, 10, 25–35. [Google Scholar]
- Lok, C.E.; Huber, T.S.; Lee, T.; Shenoy, S.; Yevzlin, A.S.; Abreo, K.; Allon, M.; Asif, A.; Astor, B.C.; Glickman, M.H.; et al. Kdoqi clinical practice guideline for vascular access: 2019 update. Am. J. Kidney Dis. 2020, 75, S1–S164. [Google Scholar] [CrossRef]
- Bivet, F. Nonuse of RIFLE classification urine output criteria: Biases for acute kidney injury biomarkers performance assessment? Crit. Care Med. 2012, 40, 1692–1693. [Google Scholar] [CrossRef]
- Cupples, W.A.; Braam, B. Assessment of renal autoregulation. Am. J. Physiol. Ren. Physiol. 2007, 292, 1105–1123. [Google Scholar] [CrossRef]
- Kanji, H.D.; Schulze, C.J.; Hervas-Malo, M. Difference between pre-operative and cardiopulmonary bypass mean arterial pressure is independently associated with early cardiac surgery-associated acute kidney injury. J. Cardiothorac. Surg. 2010, 5, 71. [Google Scholar] [CrossRef]
- Robert, A.M.; Kramer, R.S.; Dacey, L.I. Cardiac surgery-associated acute kidney injury: A comparison of two consensus criteria. Ann. Thorac Surg. 2010, 90, 1939–1943. [Google Scholar] [CrossRef]
- Weir, M.R.; Aronson, S.; Avery, E.G. Acute kidney injury following cardiac surgery: Role of perioperative blood pressure control. Am. J. Nephrol. 2011, 33, 438–452. [Google Scholar] [CrossRef]
- Cupples, W.; Loutzenhiser, R. Dynamic autoregulation in the in vitro perfused hydronephrotic rat kidney. Am. J. Physiol.—Ren. Physiol. 1998, 275, 126–130. [Google Scholar] [CrossRef]
- Holstein-Rathlou, N.; Wagner, A.; Marsh, D. Tubloglomerular feedback dynamics and renal blood flow autoregulation in rats. Am. J. Physiol.—Ren. Physiol. 1991, 260, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Just, A. Mechanisms of renal blood flow autoregulation: Dynamics and contributions. Am. J. Physiol.—Regul. Integr. Comp. Physiol. 2007, 292, R1–R17. [Google Scholar] [CrossRef]
- Just, A.; Arendshorst, W.J. Dynamics and contribution of mechanisms mediating renal blood flow autoregulation. Am. J. Physiol.—Regul. Integr. Comp. Physiol. 2003, 285, 619–631. [Google Scholar] [CrossRef]
- Loutzenhiser, R.; Bidani, A.; Chilton, L. Renal myogenic response: Kinetic attributes and physiological role. Circ. Res. 2002, 90, 1316–1324. [Google Scholar] [CrossRef]
- Loutzenhiser, R.; Griffin, K.; Williamson, G.; Bidani, A. Renal autoregulation: New perspectives regarding the protective and regulatory roles of the underlying mechanisms. Am. J. Physiol.—Regul. Integr. Comput. Physiol. 2006, 290, 1156–1167. [Google Scholar] [CrossRef]
- Lush, D.J.; Fray, J.C. Steady-state autoregulation of renal blood flow: A myogenic model. Am. J. Physiol.—Regul. Integr. Comput. Physiol. 1984, 247, 89–99. [Google Scholar] [CrossRef]
- Marsh, D.; Sosnovtseva, O.; Chon, K.; Holstein-Rathlou, N. Nonlinear interactions in renal blood blow regulation. Am. J. Physiol.—Regul. Integr. Comp. Physiol. 2005, 288, 1143–1159. [Google Scholar] [CrossRef]
- Marsh, D.; Sosnovtseva, O.; Mosekilde, E.; Holstein-Rathlou, N. Vascular coupling induces synchronization, quasiperiodicity, and chaos in a nephron tree. Chaos 2007, 17, 015114. [Google Scholar] [CrossRef]
- Marsh, D.; Sosnovtseva, O.; Pavlov, A.; Yip, K.; Holstein-Rathlou, N. Frequency encoding in renal blood blow regulation. Am. J. Physiol.—Regul. Integr. Comp. Physiol. 2005, 288, 1160–1167. [Google Scholar] [CrossRef]
- Moore, L. Tubuloglomerular feedback and SNGFR autoregulation in the rat. Am. J. Physiol.—Ren. Physiol. 1984, 247, 267–276. [Google Scholar] [CrossRef]
- Oien, A.H.; Aukland, K. A mathematical-analysis of the myogenic hypothesis with special reference to auto-regulation of renal blood-flow. Circ. Res. 1983, 52, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Persson, P. Renal blood flow autoregulation in blood pressure control. Curr. Opin. Nephrol. Hypertens. 2002, 11, 67–72. [Google Scholar] [CrossRef]
- Pires, S.L.S.; Julien, C.; Chapuis, B.; Sassard, J.; Barres, C. Spontaneous renal blood flow autoregulation curves in conscious sinoaortic baroreceptor-denervated rats. Am. J. Physiol.—Ren. Physiol. 2002, 282, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Racasan, S.; Joles, J.; Boer, P.; Koomans, H.; Braam, B. NO dependency of RBF and autoregulation in the spontaneously hypertensive rat. Am. J. Physiol.—Ren. Physiol. 2003, 285, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Turkstra, E.; Braam, B.; Koomans, H. Impaired renal blood flow autoregulation in twokidney, one-clip hypertensive rats is caused by enhanced activity of nitric oxide. J. Am. Soc. Nephrol. 2000, 11, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Sgouralis, I.; Layton, A.T. Mathematical modeling of renal hemodynamics in physiology and pathophysiology. Math. Biosci. 2015, 264, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Postnov, D.D.; Marsh, D.J.; Postnov, D.E.; Braunstein, T.H.; Holstein-Rathlou, N.H.; Martens, E.A.; Sosnovtseva, O. Modeling of Kidney Hemodynamics: Probability-Based Topology of an Arterial Network. PLoS Comput. Biol. 2016, 12, e1004922. [Google Scholar] [CrossRef] [PubMed]
- Cury, L.F.M.; Talou, G.M.; Younes-Ibrahim, M.; Blanco, P.J. Parallel generation of extensive vascular networks with application to an archetypal human kidney model. R. Soc. Open Sci. 2021, 8, 210973. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Tsubota, K.I. Numerical simulation of the vascular structure dependence of blood flow in the kidney. Med. Eng. Phys. 2022, 104, 103809. [Google Scholar] [CrossRef]
- Deng, W.; Tsubota, K.I. Numerical Modeling and Simulation of Blood Flow in a Rat Kidney: Coupling of the Myogenic Response and the Vascular Structure. Processes 2022, 10, 1005. [Google Scholar] [CrossRef]
- Holstein-Rathlon, N.H.; Marsh, D.J. A dynamic model of renal blood flow autoregulation. Bull. Math. Biol. 1994, 56, 411–429. [Google Scholar] [CrossRef]
- Sgouralis, I.; Evans, R.G.; Layton, A.T. Renal medullary and urinary oxygen tension during cardiopulmonary bypass in the rat. Math. Med. Biol. 2017, 34, 313–333. [Google Scholar] [CrossRef]
- Hao, W.; Rovin, B.H.; Friedman, A. Mathematical model of renal interstitial fibrosis. Proc. Natl. Acad. Sci. USA 2014, 111, 14193–14198. [Google Scholar] [CrossRef] [PubMed]
- Basri, A.A.; Khader, S.M.A.; Johny, C.; Raghuvir, P.B.; Zuber, M.; Ahmad, Z.; Ahmad, K.A. Effect of Single and Double Stenosed on Renal Arteries of Abdominal Aorta: A Computational Fluid Dynamics. CFD Lett. 2020, 12, 87–97. [Google Scholar]
- Black, S.M.; Maclean, C.; Hall-Barrientos, P.; Ritos, K.; Kazakidi, A. Reconstruction and Validation of Arterial Geometries from 4D Flow-MRI Images: A Novel Approach. Cardiovasc. Eng. Technol. 2023, 14, 655–676. [Google Scholar] [CrossRef] [PubMed]
- Boumpouli, M.; Sauvage, E.L.; Capelli, C.; Schievano, S.; Kazakidi, A. Characterization of Flow Dynamics in the Pulmonary Bifurcation of Patients with Repaired Tetralogy of Fallot: A Computational Approach. Front. Cardiovasc. Med. 2021, 8, 703717. [Google Scholar] [CrossRef] [PubMed]
- Grinberg, L.; Cheever, E.; Anor, T.; Madsen, J.R.; Karniadakis, G.E. Modeling blood flow circulation in intracranial arterial networks: A comparative 3D/1D simulation study. Ann. Biomed. Eng. 2011, 39, 297–309. [Google Scholar] [CrossRef]
- Grinberg, L.; Karniadakis, G.E. Outflow boundary conditions for arterial networks with multiple outlets. Ann. Biomech. Eng. 2008, 36, 1496–1514. [Google Scholar] [CrossRef] [PubMed]
- Hyde-Linaker, G.; Barientos, P.H.; Stoumpos, S.; Kingsmore, D.B.; Kazakidi, A. Patient-specific computational haemodynamics associated with the surgical creation of an arteriovenous fistula. Med. Eng. Phys. 2022, 105, 103814. [Google Scholar] [CrossRef]
- Johnston, L.; Allen, R.; Hall-Barrientos, P.; Mason, A.; Kazakidi, A. Hemodynamic Abnormalities in the Aorta of Turner Syndrome Girls. Front. Cardiovasc. Med. 2021, 8, 670841. [Google Scholar] [CrossRef]
- Kamiya, A.; Togawa, T. Optimal branching structure of the vascular tree. Bull. Math. Biophys. 1972, 34, 431–438. [Google Scholar] [CrossRef]
- Kazakidi, A.; Sherwin, S.J.; Weinberg, P.D. Effect of Reynolds number and flow division on patterns of haemodynamic wall shear stress near branch points in the descending thoracic aorta. J. R. Soc. Interface 2009, 6, 539–548. [Google Scholar] [CrossRef]
- Watanabe, S.M.; Blanco, P.J.; Feijóo, R.A. Mathematica Model of Blood Flow in an Anatomically Detailed Arterial Network of the Arm. ESAIM Math. Model. Numer. Anal. 2013, 47, 961–985. [Google Scholar] [CrossRef]
- Boumpouli, M.; Danton, M.H.D.; Gourlay, T.; Kazakidi, A. Blood flow simulations in the pulmonary bifurcation in relation to adult patients with repaired tetralogy of Fallot. Med. Eng. Phys. 2020, 85, 123–138. [Google Scholar] [CrossRef]
- Van Doormaal, M.A.; Kazakidi, A.; Wylezinska, M.; Hunt, A.; Tremoleda, J.L.; Protti, A.; Bohraus, Y.; Gsell, W.; Weinberg, P.D.; Ethier, C.R. Haemodynamics in the mouse aortic arch computed from MRI-derived velocities at the aortic root. J. R. Soc. Interface 2012, 9, 2834–2844. [Google Scholar] [CrossRef]
- Johnston, L.; Boumpouli, M.; Kazakidi, A. Hemodynamics in the Aorta and Pulmonary Arteries of Congenital Heart Disease Patients: A Mini Review. J. Cardiol. Cardiovasc. Sci. 2021, 5, 1–5. [Google Scholar] [CrossRef]
- Kazakidi, A.; Plata, A.M.; Sherwin, S.J.; Weinberg, P.D. Effect of reverse flow on the pattern of wall shear stress near arterial branches. J. R. Soc. Interface 2011, 8, 1594–1603. [Google Scholar] [CrossRef]
- Pedley, T.J.; Schroter, R.C.; Sudlow, M.F. Flow and pressure drop in systems of repeatedly branching tubes. J. Fluid Mech. 1971, 46, 365–383. [Google Scholar] [CrossRef]
- Shi, Y.; Lawford, P.; Hose, R. Review of Zero-D and 1-D models of blood flow in the cardiovascular system. BioMedical Eng. Online 2011, 10, 219. [Google Scholar] [CrossRef]
- Black, S.M.; Maclean, C.; Barrientos, P.H.; Ritos, K.; McQueen, A.; Kazakidi, A. Calibration of patient-specific boundary conditions for coupled CFD models of the aorta derived from 4D Flow-MRI. Front. Bioeng. Biotechnol. 2023, 11, 1178483. [Google Scholar] [CrossRef]
- Johnston, L.; Allen, R.; Mason, A.; Kazakidi, A. Morphological characterisation of pediatric Turner syndrome aortae: Insights from a small cohort study. Med. Eng. Phys. 2023, 120, 104045. [Google Scholar] [CrossRef]
- Sherman, T.F. On connecting large vessels to small. The meaning of Murray’s law. J. Gen. Physiol. 1981, 78, 431–453. [Google Scholar] [CrossRef]
- Chnafa, C.; Brina, O.; Pereira, V.M.; Steinman, D.A. Better Than Nothing: A Rational Approach for Minimizing the Impact of Outflow Strategy on Cerebrovascular Simulations. AJNR. Am. J. Neuroradiol. 2018, 39, 337–343. [Google Scholar] [CrossRef]
- Yang, J.; Pak, Y.E.; Lee, T.-R. Predicting bifurcation angle effect on blood flow in the microvasculature. Microvasc. Res. 2016, 108, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Bessonov, N.; Sequeira, A.; Simakov, S.; Vassilevskii, Y.; Volpert, V. Methods of blood flow modelling. Math. Model. Nat. Phenom. 2016, 11, 1–25. [Google Scholar] [CrossRef]
- Aroesty, J.; Gross, J.F. The mathematics of pulsatile flow in small vessels I. Casson theory. Microvasc. Res. 1972, 4, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Stark, H.; Schuster, S. Comparison of various approaches to calculating the hematocrit in vertebrates. J. Appl. Physiol. 2012, 113, 355–367. [Google Scholar] [CrossRef]
- Sochi, T. Analytical solutions for the flow of Carreau and Cross fluids in circular pipes and thin slits. Rheol. Acta 2015, 54, 745–756. [Google Scholar] [CrossRef]
- Abu-Naser, M.; Williamson, G.A.; Bidani, A.K.; Griffin, K.A. Vascular resistance estimation in real hemodynamics using a time-varying Windkessel model. In Proceedings of the IEEE International Conference on Acoustics, Speech, and Signal Processing, Philadelphia, PA, USA, 23 March 2005; Volume 5, pp. 641–644. [Google Scholar]
- Collard, D.; Brussel, P.M.v.; Velde, L.v.d.; Wijntjens, G.W.M.; Westerhof, B.E.; Karemaker, J.M.; Piek, J.J.; Reekers, J.A.; Vogt, L.; Winter, R.J.D.; et al. Estimation of Intraglomerular Pressure Using Invasive Renal Arterial Pressure and Flow Velocity Measurements in Humans. J. Am. Soc. Nephrol. JASN 2020, 31, 1905–1914. [Google Scholar] [CrossRef]
- Hsu, T.L.; Hsiu, H.; Chao, P.T.; Li, S.P.; Wang, W.K.; Wang, Y.Y.L. Three-block electrical model of renal impedance. IOP Physiol. Meas. 2005, 26, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Mathematica, Version 13.0; Wolfram Research Inc.: Champaign, IL, USA, 2022.
- Zhao, Y.; Lieber, B. Steady inspiratory flow in a model symmetric bifurcation. J. Biomech. Eng. 1994, 116, 488–496. [Google Scholar] [CrossRef]
- Schroter, R.C.; Sudlow, M.F. Flow patterns in models of the human bronchial airways. Respir. Physiol. 1969, 7, 341–355. [Google Scholar] [CrossRef]
- López-Picado, A.; Albinarrate, A.; Barrachina, B. Determination of perioperative blood loss: Accuracy or approximation? Anesth. Analg. 2017, 125, 280–286. [Google Scholar] [CrossRef] [PubMed]
- Piccioli, F.; Bertaglia, G.; Valiani, A.; Caleffi, V. Modeling blood flow in networks of viscoelastic vessels with the 1-D augmented fluid–structure interaction system. J. Comput. Phys. 2022, 464, 111364. [Google Scholar] [CrossRef]
- Kalantarinia, K.; Belcik, J.T.; Patrie, J.T.; Wei, K. Real-time measurement of renal blood flow in healthy subjects using contrast-enhanced ultrasound. Am. J. Physiol. Ren. Physiol. 2009, 297, 1129–1134. [Google Scholar] [CrossRef]
- Young, L.S.; Regan, M.C.; Sweeney, P.; Barry, K.M.; Ryan, M.P.; Fitzpatrick, J.M. Changes in regional renal blood flow after unilateral nephrectomy using the techniques of autoradiography and microautoradiography. J. Urol. 1998, 160, 926–931. [Google Scholar] [CrossRef]
- Warren, D.R.; Partridge, M. The role of necrosis, acute hypoxia and chronic hypoxia in F18–FMISO PET image contrast: A computational modelling study. Phys. Med. Biol. 2016, 61, 8596–8624. [Google Scholar] [CrossRef] [PubMed]
- Miyata, H.; Abe, T.; Hotta, K.; Higuchi, M.; Osawa, T.; Matsumoto, R.; Kikuchi, H.; Kurashima, Y.; Murai, S.; Shinohara, N. Validity assessment of the laparoscopic radical nephrectomy module of the LapVision virtual reality simulator. Surg. Open Sci. 2020, 2, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Salotto, A.G.; Muscarella, L.F.; Melbin, J.; Li, J.K.J.; Noordergraaf, A. Pressure pulse transmission into vasculare beds. Microvasc. Res. 1986, 32, 152–163. [Google Scholar] [CrossRef]
- Wang, X. 1D Modelling of Blood Flow in Networks. Ph.D. Thesis, University of Pierre and Marie Curie, Paris, France, 2014. [Google Scholar]
- Andreeva, A.A.; Anand, M.; Lobanov, A.I.; Nikolaev, A.V.; Panteleev, M.A.; Susree, M. Mathematical modelling of platelet rich plasma clotting. Pointwise unified model. Russ. J. Numer. Anal. Math. Model. 2018, 33, 265–276. [Google Scholar] [CrossRef]
- Galochkina, T.; Marion, M.; Volpert, V. Initiation of reaction-diffusion waves of blood coagulation. Phys. D 2018, 376, 160–170. [Google Scholar] [CrossRef]
- Ratto, N.; Bouchnita, A.; Chelle, P.; Marion, M.; Panteleev, M.; Nechipurenko, D.; Tardy-Poncet, B.; Volpert, V. Patient-specific modelling of blood coagulation. Bull. Math. Biol. 2021, 83, 2243. [Google Scholar] [CrossRef]
- Hund, S.J.; Kameneva, M.V.; Antaki, J.F. A quasi-mechanistic mathematical representation for blood viscosity. Fluids 2017, 2, 10. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physical Parameter | Value |
|---|---|
| RTOT = RL1 = RR1 | 2.7 × 10−3 m |
| RL5 = RL9 = RL18 | 2.5 × 10−3 m |
| RR10 = RR20 = RR29 = RR33 | 2.5 × 10−3 m |
| The remaining radii, R | 1.6 × 10−3 m |
| All lengths, L | 4 × 10−2 m |
| 600 s−1 | |
| λ | 3.313 s |
| 0.056 Pas | |
| 0.00345 Pas | |
| n | 0.3568 |
| f | 10 Hz |
| ZTOT = ZL1 = ZL5 = ZL9 = ZL18 | 1.649 × 109 kgm−4s−1 |
| ZR1 = ZR10 = ZR20 = ZR29 = ZR33 | 1.649 × 109 kgm−4s−1 |
| All remaining impedances, Z | 3.185 × 109 kgm−4s−1 |
| QTOT | 600 mLmin−1 (10−5 m3s−1) |
| Branch | Z1 (kgm−4s−1) | Z2 (kgm−4s−1) | C (kg−1m4s2) | |Z| (kgm−4s−1) |
|---|---|---|---|---|
| QTOT = QL1 = QL5 = QL9 = QL18 | 1.180 × 109 | 4.830 × 108 | 1.840 × 10−12 | 1.649 × 109 |
| QR1 = QR10 = QR20 = QR29 = QR33 | 1.180 × 109 | 4.830 × 108 | 1.840 × 10−12 | 1.649 × 109 |
| All remaining vessels | 6.136 × 109 | 2.512 × 109 | 9.568 × 10−12 | 3.316 × 109 |
| Branch. | Blood Flux (mL/min) | Blood Pressure (mmHg) | Branch | Blood Flux (mL/min) | Blood Pressure (mmHg) |
|---|---|---|---|---|---|
| QL1 | 212.68 | 44.42 | QL2 | 37.86 | 15.27 |
| QL3 | 18.93 | 7.64 | QL4 | 18.93 | 7.64 |
| QL5 | 174.82 | 36.52 | QL6 | 35.08 | 14.15 |
| QL7 | 17.54 | 7.08 | QL8 | 17.54 | 7.08 |
| QL9 | 139.75 | 29.19 | QL10 | 44.08 | 17.78 |
| QL11 | 16.15 | 6.51 | QL12 | 27.93 | 11.27 |
| QL13 | 13.97 | 5.63 | QL14 | 13.97 | 5.63 |
| QL15 | 32.29 | 13.03 | QL16 | 16.15 | 6.51 |
| QL17 | 16.15 | 6.51 | QL18 | 63.38 | 13.24 |
| QL19 | 16.94 | 6.83 | QL20 | 16.94 | 6.83 |
| QL21 | 29.51 | 11.90 | QL22 | 14.75 | 5.95 |
| QL23 | 14.75 | 5.95 | |||
| QR1 | 387.32 | 80.90 | QR2 | 85.93 | 34.67 |
| QR3 | 18.93 | 7.64 | QR4 | 33.50 | 13.51 |
| QR5 | 33.50 | 13.51 | QR6 | 16.75 | 6.76 |
| QR7 | 16.75 | 6.76 | QR8 | 16.75 | 6.76 |
| QR9 | 16.75 | 6.76 | QR10 | 301.40 | 62.95 |
| QR11 | 96.50 | 38.93 | QR12 | 17.54 | 7.08 |
| QR13 | 17.54 | 7.08 | QR14 | 30.71 | 12.39 |
| QR15 | 15.36 | 6.20 | QR16 | 15.36 | 6.20 |
| QR17 | 30.71 | 12.39 | QR18 | 15.36 | 6.20 |
| QR19 | 15.36 | 6.20 | QR20 | 204.90 | 42.80 |
| QR21 | 32.29 | 13.03 | QR22 | 16.15 | 6.51 |
| QR23 | 16.15 | 6.51 | QR24 | 44.08 | 17.78 |
| QR25 | 16.15 | 6.51 | QR26 | 27.93 | 11.27 |
| QR27 | 13.97 | 5.63 | QR28 | 13.97 | 5.63 |
| QR29 | 128.53 | 26.85 | QR30 | 16.94 | 6.83 |
| QR31 | 16.94 | 6.83 | QR32 | 16.94 | 6.83 |
| QR33 | 77.72 | 16.23 | QR34 | 15.54 | 6.27 |
| QR35 | 15.54 | 6.27 | QR36 | 15.54 | 6.27 |
| QR37 | 15.54 | 6.27 | QR38 | 15.54 | 6.27 |
| Branch Cut | Blood Loss (mL/min) | % Blood Loss for the Human Body |
|---|---|---|
| QL1 | 57.48 | 1.15 |
| QL5 | 51.46 | 1.03 |
| QL9 | 46.07 | 0.92 |
| QL18 | 38.38 | 0.77 |
| QR1 | 89.27 | 1.79 |
| QR10 | 67.41 | 1.35 |
| QR20 | 51.92 | 1.04 |
| QR29 | 42.64 | 0.85 |
| QL3 | 18.93 | 0.38 |
| QL7 | 17.54 | 0.35 |
| QL13 | 13.97 | 0.28 |
| QL23 | 14.75 | 0.30 |
| QR9 | 16.75 | 0.34 |
| QR28 | 13.97 | 0.28 |
| QR32 | 16.94 | 0.34 |
| QR38 | 15.54 | 0.31 |
| Branch Cuts | Blood Loss (mL/min) | % Blood Loss | Blood Loss (mL/min) | % Blood Loss | % Total Blood Loss |
|---|---|---|---|---|---|
| QL2 and QL5 | QL2 = 26.65 | QL2 = 0.53 | QL5 = 53.00 | QL5 = 1.06 | QL2 & QL5 = 1.59 |
| QL10 and QL18 | QL10 = 19.91 | QL10 = 0.40 | QL18 = 39.99 | QL18 = 0.80 | QL10 & QL18 = 1.20 |
| QL15 and QL21 | QL15 = 19.09 | QL15 = 0.38 | QL21 = 17.69 | QL21 = 0.35 | QL15 & QL21 = 0.73 |
| QR2 and QR10 | QR2 = 41.44 | QR2 = 0.83 | QR10 = 81.57 | QR10 = 1.63 | QR2 & QR10 = 2.46 |
| QR11 and QR20 | QR11 = 31.28 | QR11 = 0.63 | QR20 = 61.95 | QR20 = 1.24 | QR11 & QR20 = 1.87 |
| QR17 and QR29 | QR17 = 21.04 | QR17 = 0.42 | QR29 = 43.68 | QR29 = 0.87 | QR17 & QR29 = 1.29 |
| QR24 and QR29 | QR24 = 22.38 | QR24 = 0.45 | QR29 = 44.76 | QR29 = 0.90 | QR24 & QR29 = 1.35 |
| QL18 and QR33 | QL18 = 41.16 | QL18 = 0.82 | QR33 = 38.48 | QR33 = 0.77 | QL18 & QR33 = 1.59 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cowley, J.; Luo, X.; Stewart, G.D.; Shu, W.; Kazakidi, A. A Mathematical Model of Blood Loss during Renal Resection. Fluids 2023, 8, 316. https://doi.org/10.3390/fluids8120316
Cowley J, Luo X, Stewart GD, Shu W, Kazakidi A. A Mathematical Model of Blood Loss during Renal Resection. Fluids. 2023; 8(12):316. https://doi.org/10.3390/fluids8120316
Chicago/Turabian StyleCowley, James, Xichun Luo, Grant D. Stewart, Wenmiao Shu, and Asimina Kazakidi. 2023. "A Mathematical Model of Blood Loss during Renal Resection" Fluids 8, no. 12: 316. https://doi.org/10.3390/fluids8120316
APA StyleCowley, J., Luo, X., Stewart, G. D., Shu, W., & Kazakidi, A. (2023). A Mathematical Model of Blood Loss during Renal Resection. Fluids, 8(12), 316. https://doi.org/10.3390/fluids8120316