
Epstein-Barr Virus EBER Transcripts Affect miRNA-Mediated Regulation of Specific Targets and Are Processed to Small RNA Species

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Cell Culture
2.2. RNA Sequencing
2.3. Dual Luciferase Assays
2.4. Plasmids
2.5. Primer Extension
2.6. RNA Isolation from Cell Lines/Tissues and Northern Blotting
2.7. La Immunoprecipitation and Detection of Associated RNA
2.8. Western Blotting
2.9. ELISA
3. Results
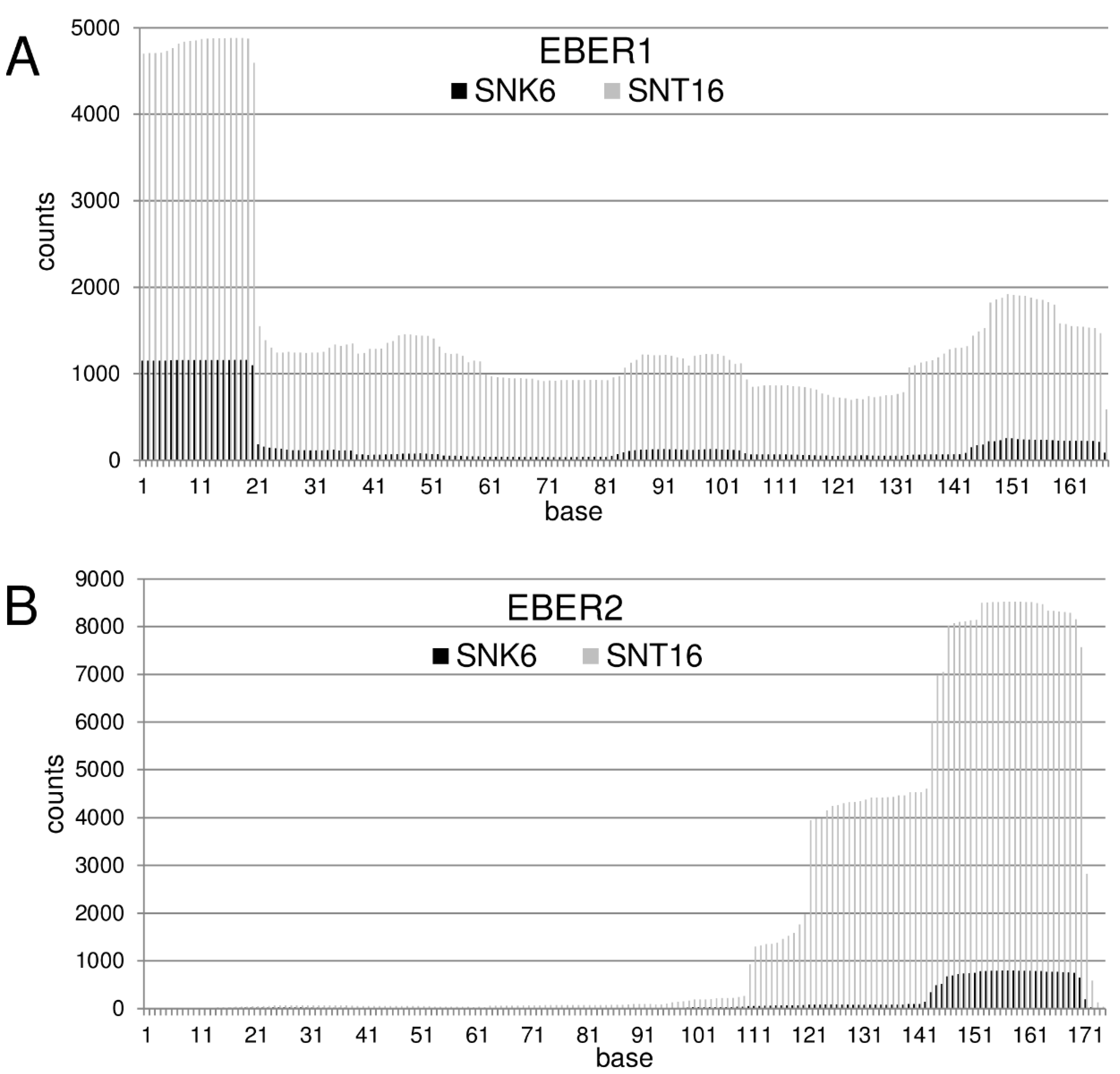
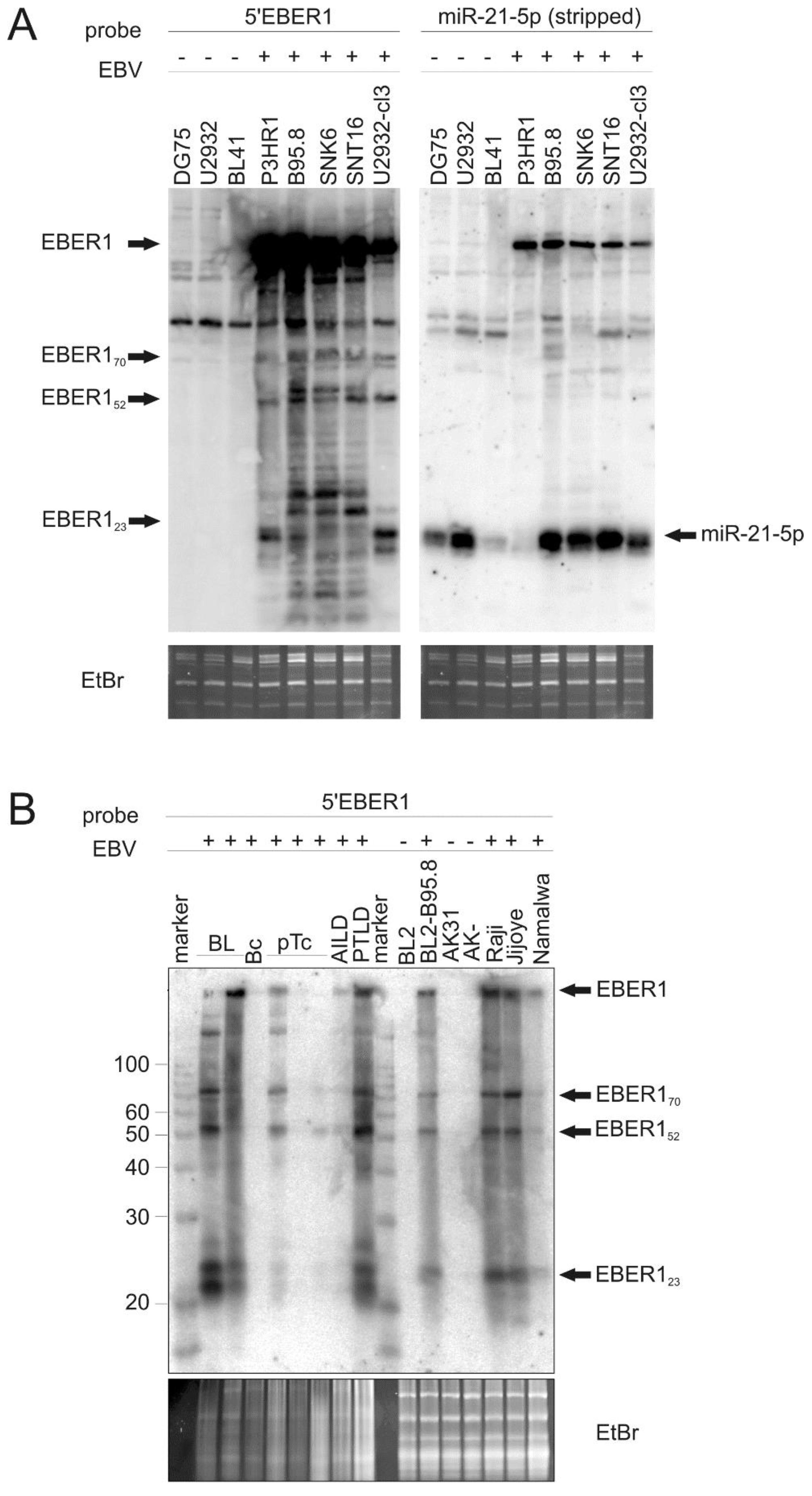
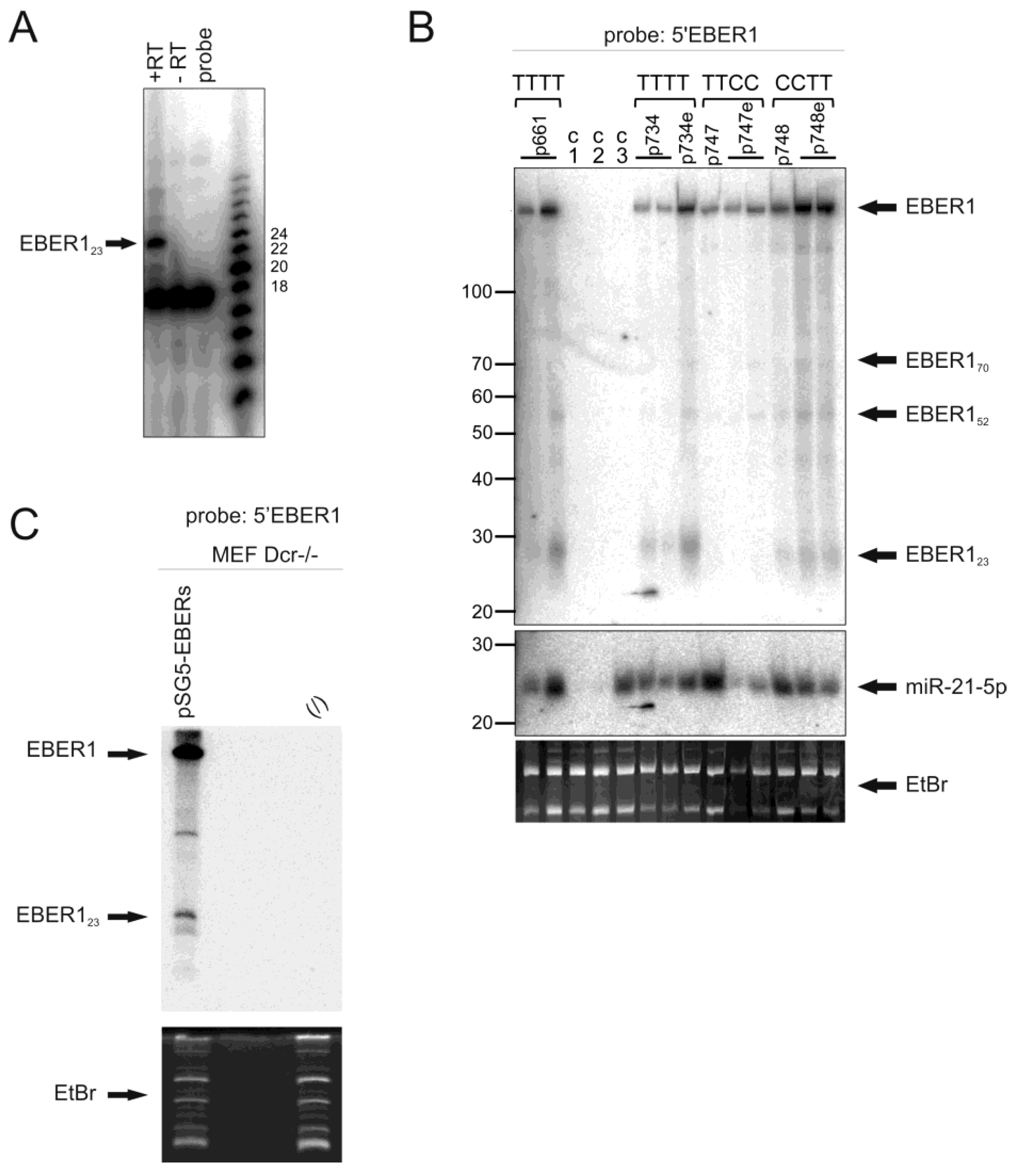
3.1. EBER RNAs Are Processed to Small Fragments



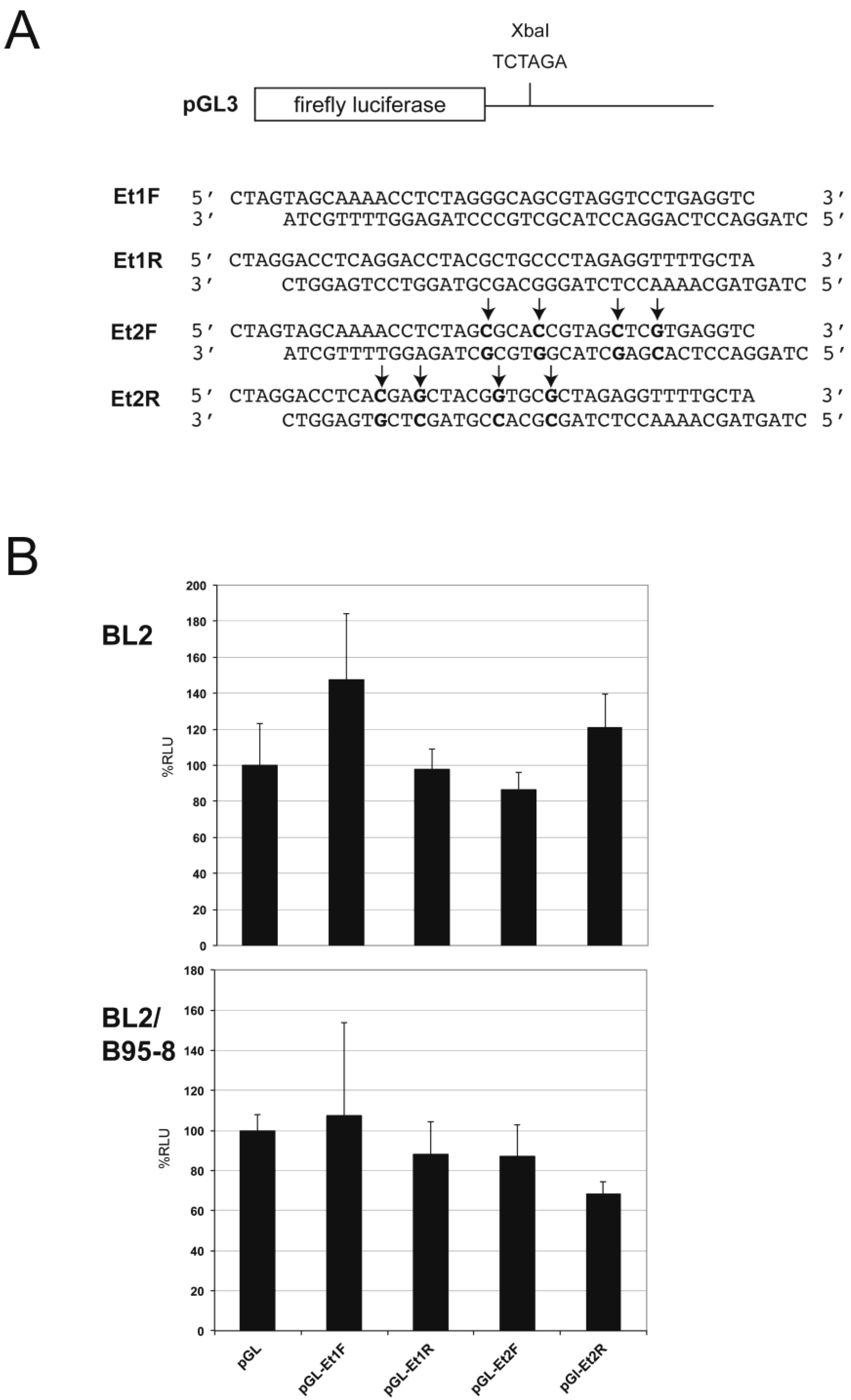
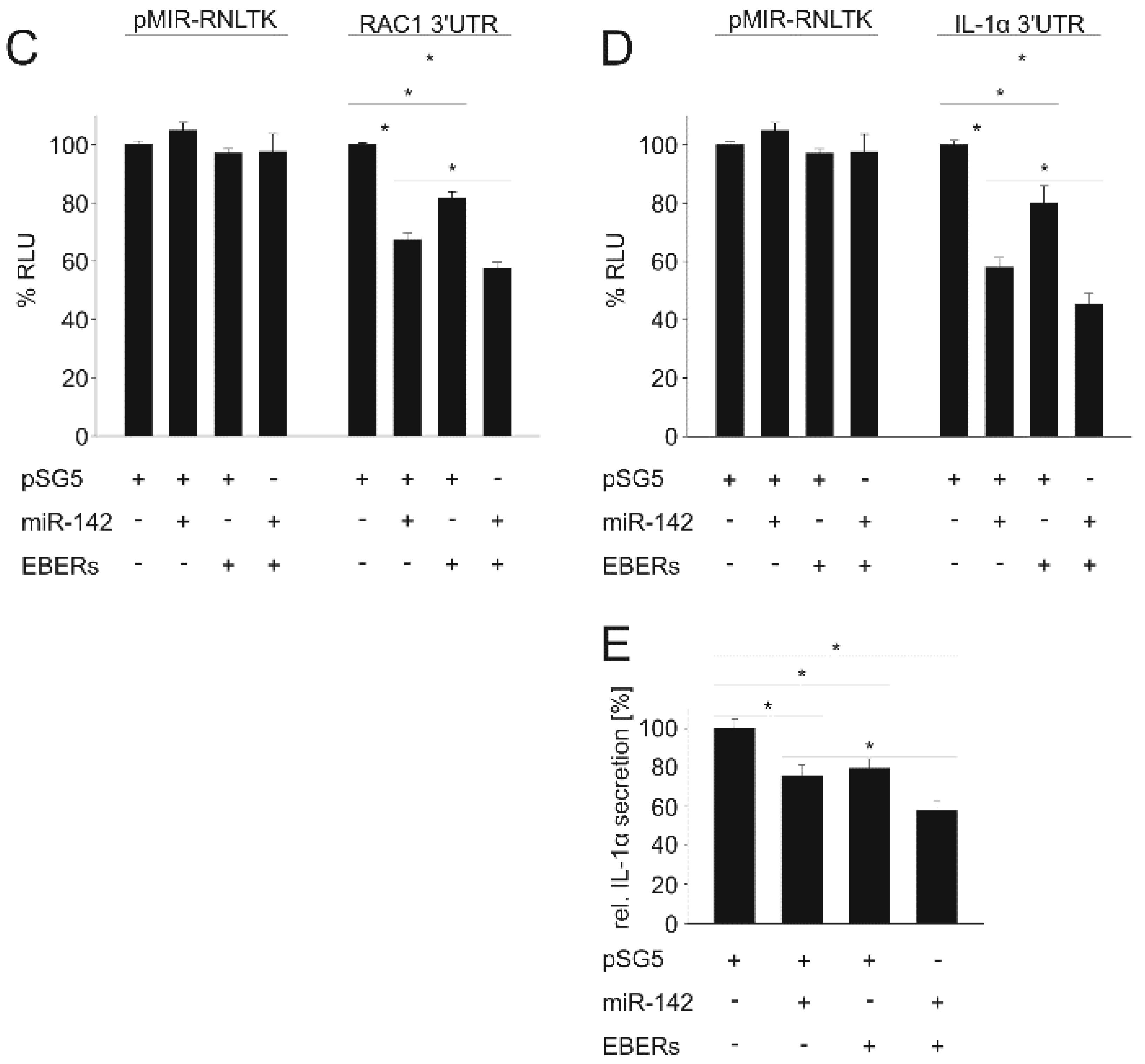
3.2. EBER1/2 Affects Cellular and Viral miRNA-Guided Repression of Specific mRNAs



3.3. The Small EBER123 Fragment Is Bound to the Lupus Antigen (La)

4. Discussion
Supplementary Files
Supplementary File 1Acknowledgments
Author Contribution
Conflicts of Interest
References
- Kieff, E.; Rickinson, A.E. Epstein-barr virus and its replication. In Fields Virology, 5th ed.; Knipe, D., Griffin, D.E., Lamb, R.A., Strauss, S.E., Howley, P.M., Martin, M.A., Roizman, B., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; Volume 2, pp. 2603–2654. [Google Scholar]
- Delecluse, H.J.; Feederle, R.; O’Sullivan, B.; Taniere, P. Epstein barr virus-associated tumors: An update for the attention of the working pathologist. J. Clin. Pathol. 2007, 60, 1358–1364. [Google Scholar] [CrossRef] [PubMed]
- Rickinson, A.B.; Kieff, E. Epstein-barr virus. In Fields Virology; Knipe, D., Griffin, D.E., Lamb, R.A., Strauss, S.E., Howley, P.M., Martin, M.A., Roizman, B., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; Volume 2, pp. 2655–2700. [Google Scholar]
- Pfeffer, S.; Zavolan, M.; Grasser, F.A.; Chien, M.; Russo, J.J.; Ju, J.; John, B.; Enright, A.J.; Marks, D.; Sander, C.; et al. Identification of virus-encoded micrornas. Science 2004, 304, 734–736. [Google Scholar] [CrossRef] [PubMed]
- Feederle, R.; Linnstaedt, S.D.; Bannert, H.; Lips, H.; Bencun, M.; Cullen, B.R.; Delecluse, H.J. A viral microrna cluster strongly potentiates the transforming properties of a human herpesvirus. PLoS Pathog. 2011, 7, e1001294. [Google Scholar] [CrossRef] [PubMed]
- Seto, E.; Moosmann, A.; Gromminger, S.; Walz, N.; Grundhoff, A.; Hammerschmidt, W. Micro rnas of epstein-barr virus promote cell cycle progression and prevent apoptosis of primary human b cells. PLoS Pathog. 2010, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barth, S.; Meister, G.; Grasser, F.A. Ebv-encoded mirnas. Biochim. Biophys. Acta 2011, 1809, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Skalsky, R.L.; Cullen, B.R. Viruses, micrornas, and host interactions. Annu. Rev. Microbiol. 2010, 64, 123–141. [Google Scholar] [CrossRef] [PubMed]
- Meister, G. Argonaute proteins: Functional insights and emerging roles. Nat. Rev. Genet. 2013, 14, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Braun, J.E.; Huntzinger, E.; Izaurralde, E. The role of gw182 proteins in mirna-mediated gene silencing. Adv. Exp. Med. Biol. 2013, 768, 147–163. [Google Scholar] [PubMed]
- Pfaff, J.; Meister, G. Argonaute and gw182 proteins: An effective alliance in gene silencing. Biochem. Soc. Trans. 2013, 41, 855–860. [Google Scholar] [CrossRef] [PubMed]
- Peters, L.; Meister, G. Argonaute proteins: Mediators of rna silencing. Mol. Cell 2007, 26, 611–623. [Google Scholar] [CrossRef] [PubMed]
- Koppers-Lalic, D.; Hackenberg, M.; Bijnsdorp, I.V.; van Eijndhoven, M.A.; Sadek, P.; Sie, D.; Zini, N.; Middeldorp, J.M.; Ylstra, B.; de Menezes, R.X.; et al. Nontemplated nucleotide additions distinguish the small rna composition in cells from exosomes. Cell Rep. 2014, 8, 1649–1658. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Pressman, S.; Andress, A.P.; Kim, K.; White, J.L.; Cassidy, J.J.; Li, X.; Lubell, K.; Lim do, H.; Cho, I.S.; et al. Silencing by small rnas is linked to endosomal trafficking. Nat. Cell Biol. 2009, 11, 1150–1156. [Google Scholar] [CrossRef] [PubMed]
- Gibbings, D.J.; Ciaudo, C.; Erhardt, M.; Voinnet, O. Multivesicular bodies associate with components of mirna effector complexes and modulate mirna activity. Nat. Cell Biol. 2009, 11, 1143–1149. [Google Scholar] [CrossRef] [PubMed]
- Yao, B.; La, L.B.; Chen, Y.C.; Chang, L.J.; Chan, E.K. Defining a new role of gw182 in maintaining mirna stability. EMBO Rep. 2012, 13, 1102–1108. [Google Scholar] [CrossRef] [PubMed]
- Lerner, M.R.; Andrews, N.C.; Miller, G.; Steitz, J.A. Two small rnas encoded by epstein-barr virus and complexed with protein are precipitated by antibodies from patients with systemic lupus erythematosus. Proc. Natl. Acad. Sci. USA 1981, 78, 805–809. [Google Scholar] [CrossRef] [PubMed]
- Howe, J.G.; Shu, M.D. Isolation and characterization of the genes for two small rnas of herpesvirus papio and their comparison with epstein-barr virus-encoded eber rnas. J. Virol. 1988, 62, 2790–2798. [Google Scholar] [PubMed]
- Iwakiri, D. Epstein-barr virus-encoded rnas: Key molecules in viral pathogenesis. Cancers (Basel) 2014, 6, 1615–1630. [Google Scholar] [CrossRef] [PubMed]
- Schwemmle, M.; Clemens, M.J.; Hilse, K.; Pfeifer, K.; Troster, H.; Muller, W.E.; Bachmann, M. Localization of epstein-barr virus-encoded rnas eber-1 and eber-2 in interphase and mitotic burkitt lymphoma cells. Proc. Natl. Acad. Sci. USA 1992, 89, 10292–10296. [Google Scholar] [CrossRef] [PubMed]
- Rymo, L. Identification of transcribed regions of epstein-barr virus DNA in burkitt lymphoma-derived cells. J. Virol. 1979, 32, 8–18. [Google Scholar] [PubMed]
- Iwakiri, D.; Zhou, L.; Samanta, M.; Matsumoto, M.; Ebihara, T.; Seya, T.; Imai, S.; Fujieda, M.; Kawa, K.; Takada, K. Epstein-barr virus (ebv)-encoded small rna is released from ebv-infected cells and activates signaling from toll-like receptor 3. J. Exp. Med. 2009, 206, 2091–2099. [Google Scholar] [CrossRef] [PubMed]
- Rosa, M.D.; Gottlieb, E.; Lerner, M.R.; Steitz, J.A. Striking similarities are exhibited by two small epstein-barr virus-encoded ribonucleic acids and the adenovirus-associated ribonucleic acids vai and vaii. Mol. Cell. Biol. 1981, 1, 785–796. [Google Scholar] [CrossRef] [PubMed]
- Bhat, R.A.; Thimmappaya, B. Two small rnas encoded by epstein-barr virus can functionally substitute for the virus-associated rnas in the lytic growth of adenovirus 5. Proc. Natl. Acad. Sci. USA 1983, 80, 4789–4793. [Google Scholar] [CrossRef] [PubMed]
- Clarke, P.A.; Schwemmle, M.; Schickinger, J.; Hilse, K.; Clemens, M.J. Binding of epstein-barr virus small rna eber-1 to the double-stranded rna-activated protein kinase dai. Nucl. Acids Res. 1991, 19, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Toczyski, D.P.; Matera, A.G.; Ward, D.C.; Steitz, J.A. The epstein-barr virus (ebv) small rna eber1 binds and relocalizes ribosomal protein l22 in ebv-infected human b lymphocytes. Proc. Natl. Acad. Sci. USA 1994, 91, 3463–3467. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Pimienta, G.; Steitz, J.A. Auf1/hnrnp d is a novel protein partner of the eber1 noncoding rna of epstein-barr virus. Rna 2012, 18, 2073–2082. [Google Scholar] [CrossRef] [PubMed]
- Samanta, M.; Iwakiri, D.; Takada, K. Epstein-barr virus-encoded small rna induces il-10 through rig-i-mediated irf-3 signaling. Oncogene 2008, 27, 4150–4160. [Google Scholar] [CrossRef] [PubMed]
- Ablasser, A.; Bauernfeind, F.; Hartmann, G.; Latz, E.; Fitzgerald, K.A.; Hornung, V. Rig-i-dependent sensing of poly(da:Dt) through the induction of an rna polymerase iii-transcribed rna intermediate. Nat. Immunol. 2009, 10, 1065–1072. [Google Scholar] [CrossRef] [PubMed]
- Swaminathan, S.; Tomkinson, B.; Kieff, E. Recombinant epstein-barr virus with small rna (eber) genes deleted transforms lymphocytes and replicates in vitro. Proc. Natl. Acad. Sci. USA 1991, 88, 1546–1550. [Google Scholar] [CrossRef] [PubMed]
- Yajima, M.; Kanda, T.; Takada, K. Critical role of epstein-barr virus (ebv)-encoded rna in efficient ebv-induced b-lymphocyte growth transformation. J. Virol. 2005, 79, 4298–4307. [Google Scholar] [CrossRef] [PubMed]
- Takada, K. Role of eber and barf1 in nasopharyngeal carcinoma (npc) tumorigenesis. Semin. Cancer Biol. 2012, 22, 162–165. [Google Scholar] [CrossRef] [PubMed]
- Iwakiri, D.; Eizuru, Y.; Tokunaga, M.; Takada, K. Autocrine growth of epstein-barr virus-positive gastric carcinoma cells mediated by an epstein-barr virus-encoded small rna. Cancer Res. 2003, 63, 7062–7067. [Google Scholar] [PubMed]
- Iwakiri, D.; Sheen, T.S.; Chen, J.Y.; Huang, D.P.; Takada, K. Epstein-barr virus-encoded small rna induces insulin-like growth factor 1 and supports growth of nasopharyngeal carcinoma-derived cell lines. Oncogene 2005, 24, 1767–1773. [Google Scholar] [CrossRef] [PubMed]
- Wong, H.L.; Wang, X.; Chang, R.C.; Jin, D.Y.; Feng, H.; Wang, Q.; Lo, K.W.; Huang, D.P.; Yuen, P.W.; Takada, K.; et al. Stable expression of ebers in immortalized nasopharyngeal epithelial cells confers resistance to apoptotic stress. Mol. Carcinog. 2005, 44, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Repellin, C.E.; Tsimbouri, P.M.; Philbey, A.W.; Wilson, J.B. Lymphoid hyperplasia and lymphoma in transgenic mice expressing the small non-coding rna, eber1 of epstein-barr virus. PLoS ONE 2010, 5, e9092. [Google Scholar] [CrossRef] [PubMed]
- Pimienta, G.; Fok, V.; Haslip, M.; Nagy, M.; Takyar, S.; Steitz, J.A. Proteomics and transcriptomics of bjab cells expressing the epstein-barr virus noncoding rnas eber1 and eber2. PLoS ONE 2015, 10, e0124638. [Google Scholar] [CrossRef] [PubMed]
- Dolken, L.; Malterer, G.; Erhard, F.; Kothe, S.; Friedel, C.C.; Suffert, G.; Marcinowski, L.; Motsch, N.; Barth, S.; Beitzinger, M.; et al. Systematic analysis of viral and cellular microrna targets in cells latently infected with human gamma-herpesviruses by risc immunoprecipitation assay. Cell Host Microbe 2010, 7, 324–334. [Google Scholar] [CrossRef] [PubMed]
- Motsch, N.; Alles, J.; Imig, J.; Zhu, J.; Barth, S.; Reineke, T.; Tinguely, M.; Cogliatti, S.; Dueck, A.; Meister, G.; et al. Microrna profiling of epstein-barr virus-associated nk/t-cell lymphomas by deep sequencing. PLoS ONE 2012, 7, e42193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motsch, N.; Pfuhl, T.; Mrazek, J.; Barth, S.; Grasser, F.A. Epstein-barr virus-encoded latent membrane protein 1 (lmp1) induces the expression of the cellular microrna mir-146a. RNA Biol. 2007, 4, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Frohn, A.; Eberl, H.C.; Stohr, J.; Glasmacher, E.; Rudel, S.; Heissmeyer, V.; Mann, M.; Meister, G. Dicer-dependent and -independent argonaute2 protein interaction networks in mammalian cells. Mol. Cell Proteom. 2012, 11, 1442–1456. [Google Scholar] [CrossRef] [PubMed]
- Imig, J.; Motsch, N.; Zhu, J.Y.; Barth, S.; Okoniewski, M.; Reineke, T.; Tinguely, M.; Faggioni, A.; Trivedi, P.; Meister, G.; et al. Microrna profiling in epstein-barr virus-associated b-cell lymphoma. Nucl. Acids Res. 2011, 39, 1880–1893. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.Y.; Pfuhl, T.; Motsch, N.; Barth, S.; Nicholls, J.; Grasser, F.; Meister, G. Identification of novel epstein-barr virus microrna genes from nasopharyngeal carcinomas. J. Virol. 2009, 83, 3333–3341. [Google Scholar] [CrossRef] [PubMed]
- Shannon-Lowe, C.; Adland, E.; Bell, A.I.; Delecluse, H.J.; Rickinson, A.B.; Rowe, M. Features distinguishing epstein-barr virus infections of epithelial cells and b cells: Viral genome expression, genome maintenance, and genome amplification. J. Virol. 2009, 83, 7749–7760. [Google Scholar] [CrossRef] [PubMed]
- Pall, G.S.; Codony-Servat, C.; Byrne, J.; Ritchie, L.; Hamilton, A. Carbodiimide-mediated cross-linking of rna to nylon membranes improves the detection of sirna, mirna and pirna by northern blot. Nucl. Acids Res. 2007, 35, e60. [Google Scholar] [CrossRef] [PubMed]
- Gross, H.; Barth, S.; Palermo, R.D.; Mamiani, A.; Hennard, C.; Zimber-Strobl, U.; West, M.J.; Kremmer, E.; Grasser, F.A. Asymmetric arginine dimethylation of epstein-barr virus nuclear antigen 2 promotes DNA targeting. Virology 2010, 397, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Grasser, F.A.; Murray, P.G.; Kremmer, E.; Klein, K.; Remberger, K.; Feiden, W.; Reynolds, G.; Niedobitek, G.; Young, L.S.; Mueller Lantzsch, N. Monoclonal antibodies directed against the epstein-barr virus-encoded nuclear antigen 1 (ebna1): Immunohistologic detection of ebna1 in the malignant cells of hodgkin’s disease. Blood 1994, 84, 3792–3798. [Google Scholar] [PubMed]
- Andersson, M.G.; Haasnoot, P.C.; Xu, N.; Berenjian, S.; Berkhout, B.; Akusjarvi, G. Suppression of rna interference by adenovirus virus-associated rna. J. Virol. 2005, 79, 9556–9565. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tan, H.; Tian, H.; Liang, C.; Chen, S.; Liu, Q. Autoantigen la promotes efficient rnai, antiviral response, and transposon silencing by facilitating multiple-turnover risc catalysis. Mol. Cell 2011, 44, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Hock, J.; Weinmann, L.; Ender, C.; Rudel, S.; Kremmer, E.; Raabe, M.; Urlaub, H.; Meister, G. Proteomic and functional analysis of argonaute-containing mrna-protein complexes in human cells. EMBO Rep. 2007, 12, 12. [Google Scholar] [CrossRef] [PubMed]
- Kwanhian, W.; Lenze, D.; Alles, J.; Motsch, N.; Barth, S.; Doll, C.; Imig, J.; Hummel, M.; Tinguely, M.; Trivedi, P.; et al. Microrna-142 is mutated in about 20% of diffuse large b-cell lymphoma. Cancer Med. 2012, 1, 141–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szczyrba, J.; Nolte, E.; Hart, M.; Doll, C.; Wach, S.; Taubert, H.; Keck, B.; Kremmer, E.; Stohr, R.; Hartmann, A.; et al. Identification of znf217, hnrnp-k, vegf-a and ipo7 as targets for micrornas that are downregulated in prostate carcinoma. Int. J. Cancer 2013, 132, 775–784. [Google Scholar] [CrossRef] [PubMed]
- Fok, V.; Friend, K.; Steitz, J.A. Epstein-barr virus noncoding rnas are confined to the nucleus, whereas their partner, the human la protein, undergoes nucleocytoplasmic shuttling. J. Cell Biol. 2006, 173, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Brenet, F.; Socci, N.D.; Sonenberg, N.; Holland, E.C. Akt phosphorylation of la regulates specific mrna translation in glial progenitors. Oncogene 2009, 28, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Vargas, K.J.; Sarmiento, J.M.; Ehrenfeld, P.; Anazco, C.C.; Villanueva, C.I.; Carmona, P.L.; Brenet, M.; Navarro, J.; Muller-Esterl, W.; Gonzalez, C.B. Postnatal expression of v2 vasopressin receptor splice variants in the rat cerebellum. Differentiation 2009, 77, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Bayfield, M.A.; Yang, R.; Maraia, R.J. Conserved and divergent features of the structure and function of la and la-related proteins (larps). Biochim. Biophys. Acta 2010, 1799, 365–378. [Google Scholar] [CrossRef] [PubMed]
- Gaidamakov, S.; Maximova, O.A.; Chon, H.; Blewett, N.H.; Wang, H.; Crawford, A.K.; Day, A.; Tulchin, N.; Crouch, R.J.; Morse, H.C., 3rd; et al. Targeted deletion of the gene encoding the la autoantigen (sjogren’s syndrome antigen b) in b cells or the frontal brain causes extensive tissue loss. Mol. Cell. Biol. 2014, 34, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Maruo, S.; Yajima, M.; Kanda, T.; Takada, K. Epstein-barr virus (ebv)-encoded rna 2 (eber2) but not eber1 plays a critical role in ebv-induced b-cell growth transformation. J. Virol. 2007, 81, 11236–11245. [Google Scholar] [CrossRef] [PubMed]
- Inada, M.; Guthrie, C. Identification of lhp1p-associated rnas by microarray analysis in saccharomyces cerevisiae reveals association with coding and noncoding rnas. Proc. Natl. Acad. Sci. USA 2004, 101, 434–439. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alles, J.; Hasler, D.; Kazmi, S.M.A.; Tesson, M.; Hamilton, A.; Schlegel, L.; Marx, S.; Eichner, N.; Reinhardt, R.; Meister, G.; et al. Epstein-Barr Virus EBER Transcripts Affect miRNA-Mediated Regulation of Specific Targets and Are Processed to Small RNA Species. Non-Coding RNA 2015, 1, 170-191. https://doi.org/10.3390/ncrna1030170
Alles J, Hasler D, Kazmi SMA, Tesson M, Hamilton A, Schlegel L, Marx S, Eichner N, Reinhardt R, Meister G, et al. Epstein-Barr Virus EBER Transcripts Affect miRNA-Mediated Regulation of Specific Targets and Are Processed to Small RNA Species. Non-Coding RNA. 2015; 1(3):170-191. https://doi.org/10.3390/ncrna1030170
Chicago/Turabian StyleAlles, Julia, Daniele Hasler, Syed Mohammad Ali Kazmi, Mathias Tesson, Andrew Hamilton, Linda Schlegel, Stefanie Marx, Norbert Eichner, Richard Reinhardt, Gunter Meister, and et al. 2015. "Epstein-Barr Virus EBER Transcripts Affect miRNA-Mediated Regulation of Specific Targets and Are Processed to Small RNA Species" Non-Coding RNA 1, no. 3: 170-191. https://doi.org/10.3390/ncrna1030170
APA StyleAlles, J., Hasler, D., Kazmi, S. M. A., Tesson, M., Hamilton, A., Schlegel, L., Marx, S., Eichner, N., Reinhardt, R., Meister, G., Wilson, J. B., & Grässer, F. A. (2015). Epstein-Barr Virus EBER Transcripts Affect miRNA-Mediated Regulation of Specific Targets and Are Processed to Small RNA Species. Non-Coding RNA, 1(3), 170-191. https://doi.org/10.3390/ncrna1030170