
Yeast Bioflavoring in Beer: Complexity Decoded and Built up Again



Abstract
:1. Introduction
2. Flavor-Active Compounds Derived from Yeast Metabolism
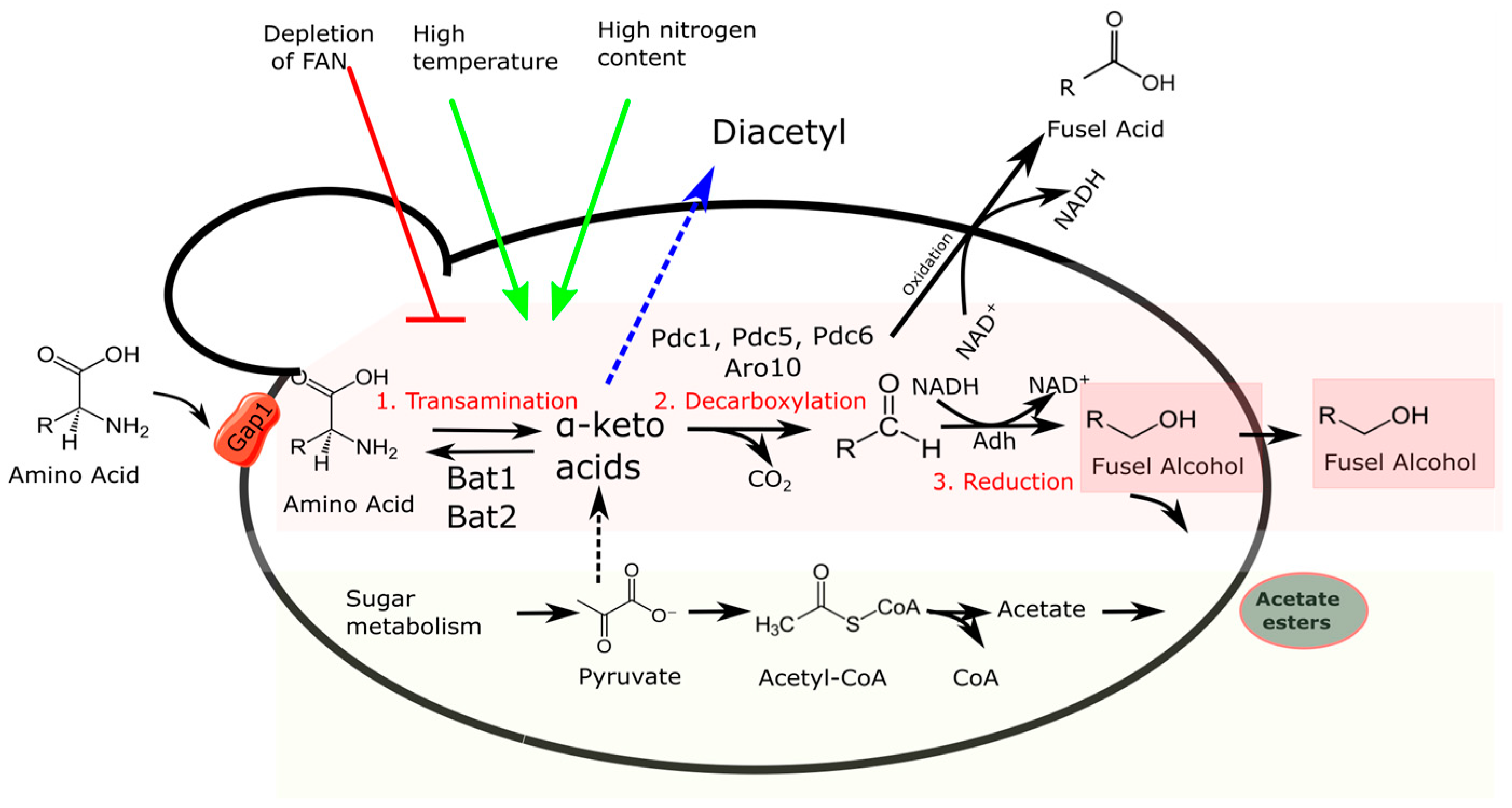
2.1. Higher Alcohols
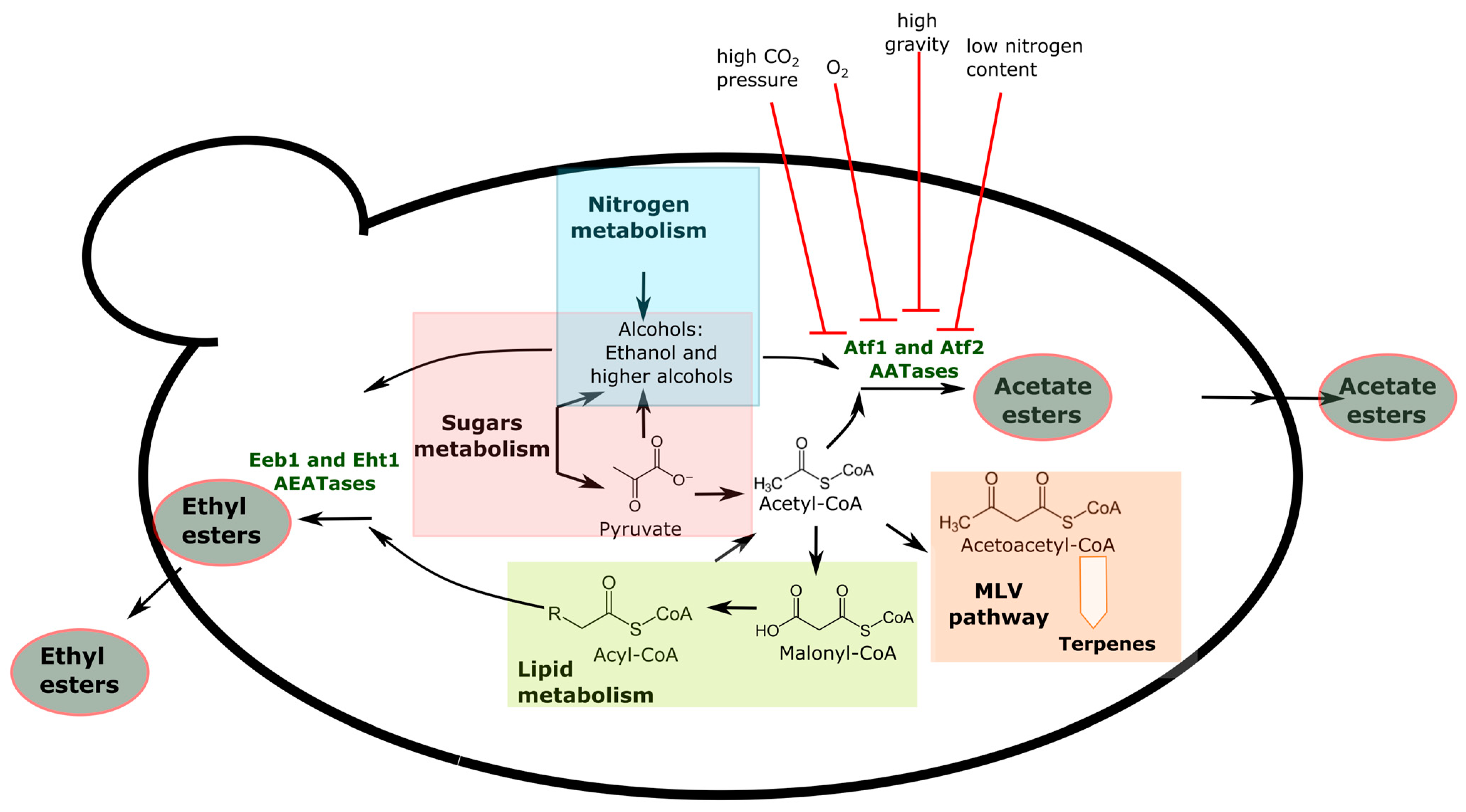
2.2. Esters and Fatty Acids
3. Flavor-Active Compounds Originated from Beer Precursors by Yeast Enzymes
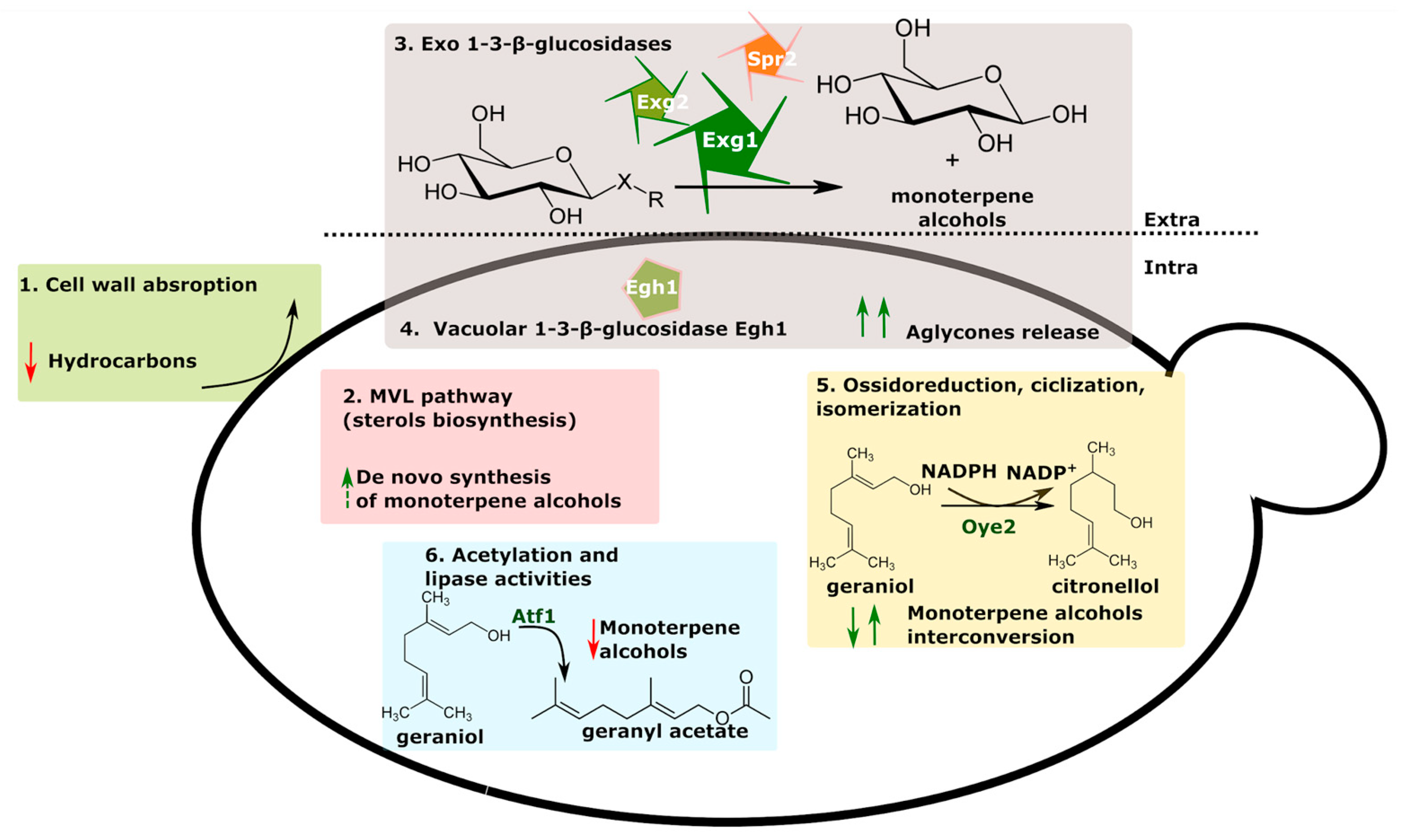
3.1. Glucosidase Activity and Monoterpene Alcohols
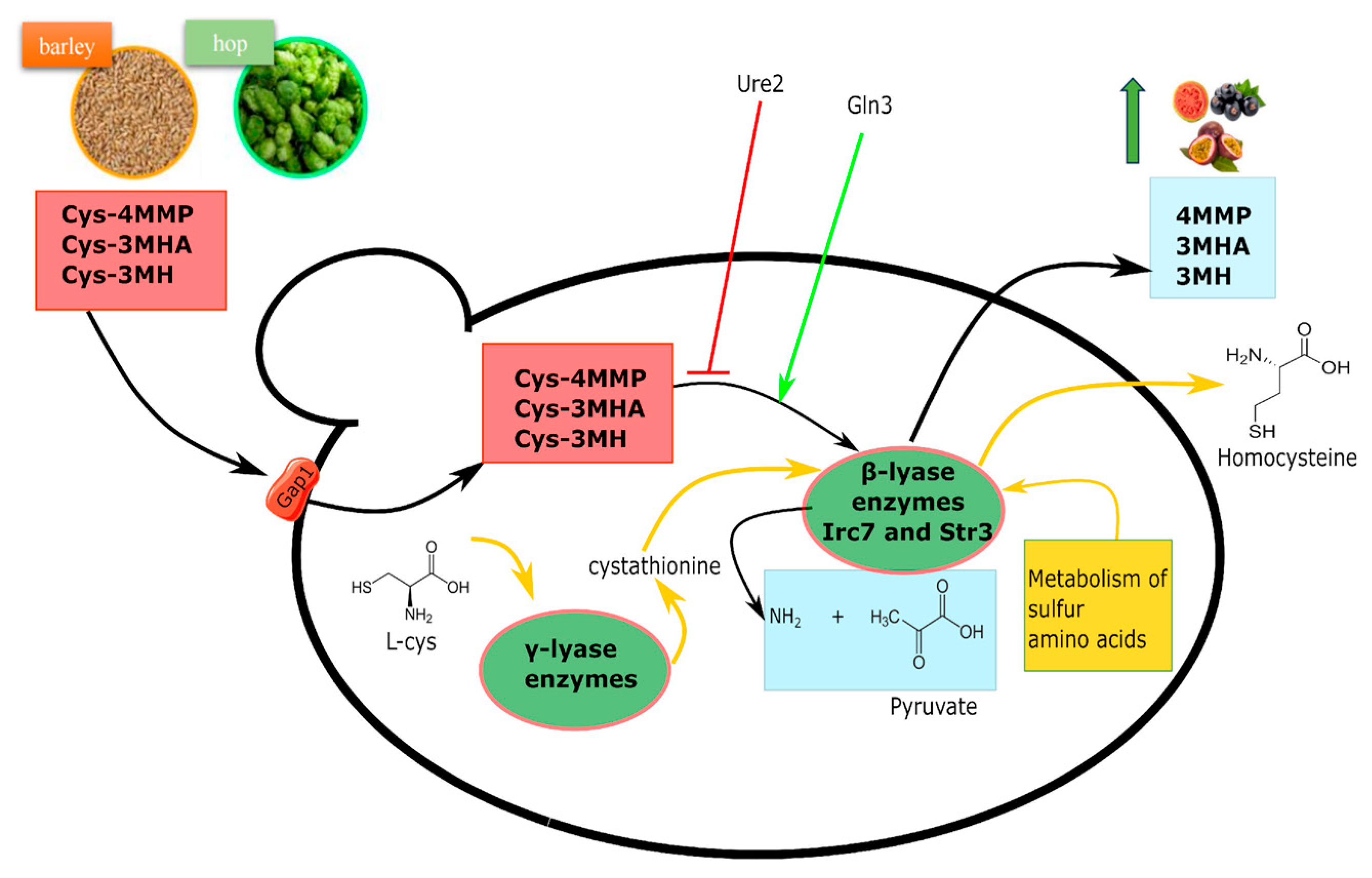
3.2. β-Lyase Activity and Thiols
3.3. Ferulic Acid Decarboxylase and Phenolic Off-Flavour Compounds
4. New Insights on Strategies to Enhance Yeast Aroma Profile
4.1. S. cerevisiae vs. Nonconventional Yeasts and the Rising of Synthetic Consortia
4.2. Bioprospecting vs. Metabolic Engineering
5. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hittinger, C.T.; Steele, J.L.; Ryder, D.S. Diverse yeasts for diverse fermented beverages and foods. Curr. Opin. Biotechnol. 2018, 49, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Libkind, D.; Hittinger, C.T.; Valério, E.; Gonçalves, C.; Dover, J.; Johnston, M.; Gonçalves, P.; Sampaio, J.P. Microbe domestication and the identification of the wild genetic stock of lager-brewing yeast. Proc. Natl. Acad. Sci. USA 2011, 108, 14539–14544. [Google Scholar] [CrossRef] [PubMed]
- Gallone, B.; Steensels, J.; Prahl, T.; Soriaga, L.; Saels, V.; Herrera-Malaver, B.; Merlevede, A.; Roncoroni, M.; Voordeckers, K.; Miraglia, L.; et al. Domestication and Divergence of Saccharomyces cerevisiae Beer Yeasts. Cell 2016, 166, 1397–1410.e16. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, M.; Pontes, A.; Almeida, P.; Barbosa, R.; Serra, M.; Libkind, D.; Hutzler, M.; Gonçalves, P.; Sampaio, J.P. Distinct domestication trajectories in top-fermenting beer yeasts and wine yeasts. Curr. Biol. 2016, 26, 2750–2761. [Google Scholar] [CrossRef]
- Meilgaard, M. Flavour chemistry of beer. Part 2 Flavour and hreshold of beer volatiles. Tech. Q. Master Brew. Assoc. Am. 1975, 12, 161–168. [Google Scholar]
- Alvarez, P.; Malcorps, P.; Sa Almeida, A.; Ferreira, A.; Meyer, A.M.; Dufour, J.P. Analysis of free fatty acids, fusel alcohols, and esters in beer: An alternative to CS2 extraction. J. Am. Soc. Brew. Chem. 1994, 52, 127–134. [Google Scholar] [CrossRef]
- Meilgaard, M.C. Prediction of flavor differences between beers from their chemical composition. J. Agric. Food Chem. 1982, 30, 1009–1017. [Google Scholar] [CrossRef]
- Haslbeck, K.; Bub, S.; von Kamp, K.; Michel, M.; Zarnkow, M.; Hutzler, M.; Coelhan, M. The influence of brewing yeast strains on monoterpene alcohols and esters contributing to the citrus flavour of beer. J. Inst. Brew. 2018, 124, 403–415. [Google Scholar] [CrossRef]
- Waterhouse, A.L.; Sacks, G.L.; Jeffery, D.W. Higher Alcohols. In Understanding Wine Chemistry, 1st ed.; Waterhouse, A.L., Sacks, G.L., Jeffery, D.W., Eds.; John Wiley & Sons: New York, NY, USA, 2016; pp. 51–56. [Google Scholar]
- Michel, M.; Haslbeck, K.; Ampenberger, F.; Meier-Dörnberg, T.; Stretz, D.; Hutzler, M.; Coelhan, M.; Jacob, F.; Liu, Y. Screening of brewing yeast β-lyase activity and release of hop volatile thiols from precursors during fermentation. Brew. Sci. 2019, 72, 179–186. [Google Scholar]
- Holt, S.; Miks, M.H.; Trindade de Carvalho, B.; Foulquié-Moreno, M.R.; Thevelein, J.M. The molecular biology of fruity and floral aromas in beer and other alcoholic beverages. FEMS Microbiol. Rev. 2019, 43, 193–222. [Google Scholar] [CrossRef]
- Buiatti, S.; Tat, L.; Natolino, A.; Passaghe, P. Biotransformations performed by yeasts on aromatic compounds provided by hop—A Review. Fermentation 2023, 9, 327. [Google Scholar] [CrossRef]
- Kind, C.; Kaiser, T. Heat, hops, Hallertau: Exploring implications of climate change for the German beer sector. In The Geography of Beer: Culture and Economics; Springer: Berlin/Heidelberg, Germany, 2020; pp. 103–111. [Google Scholar]
- Denby, C.M.; Li, R.A.; Vu, V.T.; Costello, Z.; Lin, W.; Chan, L.; Williams, J.; Donaldson, B.; Bamforth, C.W.; Petzold, C.J.; et al. Industrial brewing yeast engineered for the production of primary flavor determinants in hopped beer. Nat. Commun. 2018, 9, 965. [Google Scholar] [CrossRef] [PubMed]
- Rojas, C.; Peterson, E.B. Demand for differentiated products: Price and advertising evidence from the US beer market. Int. J. Ind. Organ. 2008, 26, 288–307. [Google Scholar] [CrossRef]
- Garavaglia, C.; Swinnen, J. The Craft Beer Revolution: An International Perspective. Choices 2017, 32, 1–8. [Google Scholar]
- Capece, A.; Romaniello, R.; Siesto, G.; Romano, P. Conventional and non-conventional yeasts in beer production. Fermentation 2018, 4, 38. [Google Scholar] [CrossRef]
- Iattici, F.; Catallo, M.; Solieri, L. Designing new yeasts for craft brewing: When natural biodiversity meets biotechnology. Beverages 2020, 6, 3. [Google Scholar] [CrossRef]
- Lengeler, K.B.; Stovicek, V.; Fennessy, R.T.; Katz, M.; Förster, J. Never Change a Brewing Yeast? Why Not, There Are Plenty to Choose From. Front. Genet. 2020, 11, 582789. [Google Scholar] [CrossRef] [PubMed]
- Cubillos, F.A.; Gibson, B.; Grijalva-Vallejos, N.; Krogerus, K.; Nikulin, J. Bioprospecting for brewers: Exploiting natural diversity for naturally diverse beers. Yeast 2019, 36, 383–398. [Google Scholar] [CrossRef]
- Dunn, B.; Kvitek, D.J.; Sherlock, G. Genetic manipulation of brewing yeasts: Challenges and opportunities. In Brewing Microbiology; Caister Academic Press: Poole, UK, 2017; p. 119. [Google Scholar]
- Gibson, B.; Geertman, J.-M.; Hittinger, C.; Krogerus, K.; Libkind, D.; Louis, E.; Magalhães, F.; Sampaio, J. New Yeasts-New Brews: Modern Approaches to Brewing Yeast Design and Development. FEMS Yeast Res. 2017, 17, fox038. [Google Scholar] [CrossRef]
- Klimczak, K.; Cioch-Skoneczny, M. Changes in beer bitterness level during the beer production process. Eur. Food Res. Technol. 2023, 249, 13–22. [Google Scholar] [CrossRef]
- Gamero, A.; Ferreira, V.; Pretorius, I.S.; Querol, A. Wine, Beer and Cider: Unravelling the Aroma Profile. In Molecular Mechanisms in Yeast Carbon Metabolism; Springer: Berlin/Heidelberg, Germany, 2014; pp. 261–297. [Google Scholar]
- Pires, E.J.; Teixeira, J.A.; Brányik, T.; Vicente, A.A. Yeast: The soul of beer’s aroma—A review of flavour-active esters and higher alcohols produced by the brewing yeast. Appl. Microbiol. Biotechnol. 2014, 98, 1937–1949. [Google Scholar] [CrossRef] [PubMed]
- Cordente, A.G.; Schmidt, S.; Beltran, G.; Torija, M.J.; Curtin, C.D. Harnessing Yeast Metabolism of Aromatic Amino Acids for Fermented Beverage Bioflavouring and Bioproduction. Appl. Microbiol. Biotechnol. 2019, 103, 4325–4336. [Google Scholar] [CrossRef]
- Svedlund, N.; Evering, S.; Gibson, B.; Krogerus, K. Fruits of their labour: Biotransformation reactions of yeasts during brewery fermentation. Appl. Microbiol. Biotechnol. 2022, 106, 4929–4944. [Google Scholar] [CrossRef]
- Briggs, D.E.; Boulton, C.A.; Brookes, P.A.; Stevens, R. Brewing: Science and Practice; Woodhead Publishing Ltd.: Abington, PA, USA, 2004. [Google Scholar]
- Procopio, S.; Qian, F.; Becker, T. Function and regulation of yeast genes involved in higher alcohol and ester metabolism during beverage fermentation. Eur. Food Res. Technol. 2011, 233, 721–729. [Google Scholar] [CrossRef]
- Humia, B.V.; Santos, K.S.; Barbosa, A.M.; Sawata, M.; da Costa Mendonça, M.; Padilha, F.F. Beer Molecules and Its Sensory and Biological Properties: A Review. Molecules 2019, 24, 1568. [Google Scholar] [CrossRef]
- Kobayashi, M.; Nagahisa, K.; Shimizu, H.; Shioya, S. Simultaneous control of apparent extract and volatile compounds concentrations in low-malt beer fermentation. Appl. Microbiol. Biotechnol. 2006, 73, 549–558. [Google Scholar] [CrossRef]
- Cordente, A.G.; Curtin, C.D.; Varela, C.; Pretorius, I.S. Flavour-active wine yeasts. Appl. Microbiol. Biotechnol. 2012, 96, 601–618. [Google Scholar] [CrossRef] [PubMed]
- Hazelwood, L.A.; Daran, J.M.; Maris, A.; Pronk, J.T.; Dickinson, J.R. The Ehrlich Pathway for Fusel Alcohol Production: A Century of Research on Saccharomyces cerevisiae Metabolism. Appl. Environ. Microbiol. 2008, 74, 2259–2266. [Google Scholar] [CrossRef]
- Eden, A.; Van Nedervelde, L.; Drukker, M.; Benvenisty, N.; Debourg, A. Involvement of branched-chain amino acid aminotransferases in the production of fusel alcohols during fermentation in yeast. Appl. Microbiol. Biotechnol. 2001, 55, 296–300. [Google Scholar] [CrossRef]
- Romagnoli, G.; Luttik, M.A.; Kotter, P.; Pronk, J.T.; Daran, J.M. Substrate specificity of thiamine pyrophosphate-dependent 2-oxo-acid decarboxylases in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2012, 78, 7538–7548. [Google Scholar] [CrossRef]
- Koonthongkaew, J.; Ploysongsri, N.; Toyokawa, Y.; Ruangpornvisuti, V.; Takagi, H. Improvement of fusel alcohol production by engineering of the yeast branched-chain amino acid aminotransaminase. Appl. Environ. Microbiol. 2022, 88, e00557-22. [Google Scholar] [CrossRef]
- Bolat, I.; Romagnoli, G.; Zhu, F.; Pronk, J.T.; Daran, J.M. Functional analysis and transcriptional regulation of two orthologs of ARO10, encoding broad-substrate-specificity 2-oxo-acid decarboxylases, in the brewing yeast Saccharomyces pastorianus CBS1483. FEMS Yeast Res. 2013, 13, 505–517. [Google Scholar] [CrossRef]
- Sun, Z.G.; Wang, M.Q.; Wang, Y.P.; Xing, S.; Hong, K.Q.; Chen, Y.F.; Guo, X.W.; Xiao, D.G. Identification by comparative transcriptomics of core regulatory genes for higher alcohol production in a top-fermenting yeast at different temperatures in beer fermentation. Appl. Microbiol. Biotechnol. 2019, 103, 4917–4929. [Google Scholar] [CrossRef]
- Cui, D.; Liu, L.; Zhang, X.; Lin, L.; Li, X.; Cheng, T.; Wei, C.; Zhang, Y.; Zhou, Z.; Li, W.; et al. Using transcriptomics to reveal the molecular mechanism of higher alcohol metabolism in Saccharomyces cerevisiae. Food Biosci. 2023, 51, 102227. [Google Scholar] [CrossRef]
- Gonzalez, J.; Lopez, G.; Argueta, S.; Escalera-Fanjul, X.; El Hafidi, M.; Campero-Basaldua, C.; Strauss, J.; Riego-Ruiz, L.; Gonzalez, A. Diversification of transcriptional regulation determines subfunctionalization of paralogous branched chain aminotransferases in the yeast Saccharomyces cerevisiae. Genetics 2017, 207, 975–991. [Google Scholar] [CrossRef]
- Ma, L.; Huang, S.; Du, L.; Tang, P.; Xiao, D. Reduced production of higher alcohols by Saccharomyces cerevisiae in red wine fermentation by simultaneously overexpressing BAT1 and deleting BAT2. J. Agric. Food Chem. 2017, 65, 6936–6942. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, G.; Engqvist, M.K.M.; Krivoruchko, A.; Hallström, B.M.; Chen, Y.; Siewers, V.; Nielsen, J. Adaptive mutations in sugar metabolism restore growth on glucose in a pyruvate decarboxylase negative yeast strain. Microb. Cell Factories 2015, 14, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Feng, R.; Wen, Z.; Zhang, A. Overexpression of ARO10 in pdc5 Δmutant resulted in higher isobutanol titers in Saccharomyces cerevisiae. Biotechnol. Bioprocess Eng. 2017, 22, 382–389. [Google Scholar] [CrossRef]
- Shen, L.; Nishimura, Y.; Matsuda, F.; Ishi, J.; Kondo, A. Overexpressing enzymes of the ehrlich pathway and deleting genes of the competing pathway in Saccharomyces cerevisiae for increasing 2-phenylethanol production from glucose. J. Biosci. Bioeng. 2016, 122, 34–39. [Google Scholar] [CrossRef]
- De Smidt, O.; du Preez, J.C.; Albertyn, J. Molecular and physiological aspects of alcohol dehydrogenases in the ethanol metabolism of Saccharomyces cerevisiae. FEMS Yeast Res. 2012, 12, 33–47. [Google Scholar] [CrossRef]
- Tapia, S.M.; Pérez-Torrado, R.; Adam, A.C.; Macías, L.G.; Barrio, E.; Querol, A. Adaptive evolution in the Saccharomyces kudriavzevii Aro4p promoted a reduced production of higher alcohols. Microb. Biotechnol. 2022, 15, 2958–2969. [Google Scholar] [CrossRef] [PubMed]
- Verstrepen, K.J.; Derdelinckx, G.; Dufour, J.-P.; Winderickx, J.; Thevelein, J.M.; Pretorius, I.S.; Delvaux, F.R. Flavor-active esters: Adding fruitiness to beer. J. Biosci. Bioeng. 2003, 96, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Krogerus, K.; Gibson, B.R. Influence of valine and other amino acids on total diacetyl and 2,3-pentanedione levels during fermentation of brewer’s wort. Appl. Microbiol. Biotechnol. 2013, 97, 6919–6930. [Google Scholar] [CrossRef] [PubMed]
- van Bergen, B.; Cyr, N.; Strasser, R.; Blanchette, M.; Sheppard, J.D.; Jardim, A. α, β-Dicarbonyl reduction is mediated by the Saccharomyces Old Yellow Enzyme. FEMS Yeast Res. 2016, 16, fow059. [Google Scholar] [CrossRef]
- Shi, T.T.; Guo, X.W.; Li, P.; Zhou, Z.; Xiao, D.G. Diacetyl content reduction in industrial brewer’s yeast through ILV2 disruption and BDH1 expression. Eur. Food Res. Technol. 2016, 242, 919–926. [Google Scholar] [CrossRef]
- Yoshimoto, H.; Bogaki, T. Mechanisms of production and control of acetate esters in yeasts. J. Biosci. Bioeng. 2023, 136, 261–269. [Google Scholar] [CrossRef]
- Pizarro, F.J.; Jewett, M.C.; Nielsen, J.; Agosin, E. Growth temperature exerts differential physiological and transcriptional responses in laboratory and wine strains of Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2008, 74, 6358–6368. [Google Scholar] [CrossRef]
- Saerens, S.M.G.; Verbelen, P.J.; Vanbeneden, N.; Thevelein, J.M.; Delvaux, F.R. Monitoring the influence of high-gravity brewing and fermentation temperature on flavour formation by analysis of gene expression levels in brewing yeast. Appl. Microbiol. Biotechnol. 2008, 80, 1039–1051. [Google Scholar] [CrossRef]
- Kucharczyk, K.; Tuszyński, T. The effect of wort aeration on fermentation, maturation and volatile components of beer produced on an industrial scale. J. Inst. Brew. 2017, 123, 31–38. [Google Scholar] [CrossRef]
- Lekkas, C.; Stewart, G.G.; Hill, A.E.; Taidi, B.; Hodgson, J. Elucidation of the role of nitrogenous wort components in yeast fermentation. J. Inst. Brew. 2007, 113, 3–8. [Google Scholar] [CrossRef]
- Vanderhaegen, B.; Neven, H.; Coghe, S.; Verstrepen, K.J.; Verachtert, H.; Derdelinckx, G. Evolution of chemical and sensory properties during aging of top-fermented beer. J. Agric. Food Chem. 2003, 51, 6782–6790. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, F.; van’t Klooster, J.S.; Ruiz, S.J.; Poolman, B. Regulation of amino acid transport in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2019, 83, e00024-19. [Google Scholar] [CrossRef] [PubMed]
- Donaton, M.C.; Holsbeeks, I.; Lagatie, O.; Van Zeebroeck, G.; Crauwels, M.; Winderickx, J.; Thevelein, J.M. The Gap1 general amino acid permease acts as an amino acid sensor for activation of protein kinase A targets in the yeast Saccharomyces cerevisiae. Mol. Microbiol. 2003, 50, 911–929. [Google Scholar] [CrossRef] [PubMed]
- James, N.; Stahl, U. Amino acid permeases and their influence on flavour compounds in beer. Brew. Sci. 2014, 67, 120–127. [Google Scholar]
- Wang, Y.P.; Liu, L.; Wang, X.S.; Hong, K.Q.; Zhang, L.H.; Sun, Z.G.; Xiao, D.G. GAT1 Gene, the GATA Transcription Activator, Regulates the Production of Higher Alcohol during Wheat Beer Fermentation by Saccharomyces cerevisiae. Bioengineering 2021, 8, 61. [Google Scholar] [CrossRef]
- Lei, H.; Li, H.; Mo, F.; Zheng, L.; Zhao, H.; Zhao, M. Effects of Lys and His supplementations on the regulation of nitrogen metabolism in lager yeast. Appl. Microbiol. Biotechnol. 2013, 97, 8913–8921. [Google Scholar] [CrossRef] [PubMed]
- Scott, W.T., Jr.; Van Mastrigt, O.; Block, D.E.; Notebaart, R.A.; Smid, E.J. Nitrogenous compound utilization and production of volatile organic compounds among commercial wine yeasts highlight strain-specific metabolic diversity. Microbiol. Spectr. 2021, 31, 10–128. [Google Scholar] [CrossRef] [PubMed]
- Mason, A.B.; Dufour, J. Alcohol acetyltransferases and the significance of ester synthesis in yeast. Yeast 2000, 16, 1287–1298. [Google Scholar] [CrossRef]
- Saerens, S.M.; Delvaux, F.R.; Verstrepen, K.J.; Thevelein, J.M. Production and biological function of volatile esters in Saccharomyces cerevisiae. Microb. Biotechnol. 2010, 3, 165–177. [Google Scholar] [CrossRef]
- Verstrepen, K.J.; Van Laere, S.D.; Vanderhaegen, B.M.; Derdelinckx, G.; Dufour, J.P.; Pretorius, I.S.; Winderickx, J.; Thevelein, J.M.; Delvaux, F.R. Expression levels of the yeast alcohol acetyltransferase genes ATF1, Lg-ATF1, and ATF2 control the formation of a broad range of volatile esters. Appl. Environ. Microbiol. 2003, 69, 5228–5237. [Google Scholar] [CrossRef]
- Vögtle, F.-N.; Prinz, C.; Kellermann, J.; Lottspeich, F.; Pfanner, N.; Meisinger, C. Mitochondrial Protein Turnover: Role of the Precursor Intermediate Peptidase Oct1 in Protein Stabilization. Mol. Biol. Cell 2011, 22, 2135–2143. [Google Scholar] [CrossRef] [PubMed]
- Kruis, A.J.; Levisson, M.; Mars, A.E.; van der Ploeg, M.; Garcés Daza, F.; Ellena, V.; Kengen, S.W.M.; van der Oost, J.; Weusthuis, R.A. Ethyl acetate production by the elusive alcohol acetyltransferase from yeast. Metab. Eng. 2017, 41, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Holt, S.; de Carvalho, B.T.; Foulquié-Moreno, M.R.; Thevelein, J.M. Polygenic analysis in absence of major effector ATF1 unveils novel components in yeast flavor ester biosynthesis. mBio 2018, 9, e01279-18. [Google Scholar] [CrossRef] [PubMed]
- Kruis, A.J.; Gallone, B.; Jonker, T.; Mars, A.E.; van Rijswijck, I.M.H.; Wolkers-Rooijackers, J.C.M.; Smid, E.J.; Steensels, J.; Verstrepen, K.J.; Kengen, S.W.; et al. Contribution of Eat1 and other alcohol acyltransferases to ester production in Saccharomyces cerevisiae. Front. Microbiol. 2018, 9, 3202. [Google Scholar] [CrossRef] [PubMed]
- Rollero, S.; Bloem, A.; Camarasa, C.; Sanchez, I.; Ortiz-Julien, A.; Sablayrolles, J.M.; Dequin, S.; Mouret, J.R. Combined effects of nutrients and temperature on the production of fermentative aromas by Saccharomyces cerevisiae during wine fermentation. Appl. Microbiol. Biotechnol. 2015, 99, 2291–2304. [Google Scholar] [CrossRef] [PubMed]
- Seguinot, P.; Bloem, A.; Brial, P.; Meudec, E.; Ortiz-Julien, A.; Camarasa, C. Analysing the impact of the nature of the nitrogen source on the formation of volatile compounds to unravel the aroma metabolism of two non-Saccharomyces strains. Int. J. Food Microbiol. 2020, 316, 108441. [Google Scholar] [CrossRef] [PubMed]
- Souffriau, B.; Holt, S.; Hagman, A.; De Graeve, S.; Malcorps, P.; Foulquié-Moreno, M.R.; Thevelein, J.M. Polygenic analysis of tolerance to carbon dioxide inhibition of isoamyl acetate “banana” flavor production in yeast reveals MDS3 as major causative gene. Appl. Environ. Microbiol. 2022, 88, e00814-22. [Google Scholar] [CrossRef] [PubMed]
- Saerens, S.M.G.; Verstrepen, K.; Van Laere, S.D.M.; Voet, A.; Van Dijck, P.; Delvaux, F.R.; Thevelein, J. The Saccharomyces cerevisiae EHT1 and EEB1 genes encode novel enzymes with medium-chain fatty acid ethyl ester synthesis and hydrolysis capacity. J. Biol. Chem. 2006, 281, 4446–4456. [Google Scholar] [CrossRef] [PubMed]
- Lilly, M.; Bauer, F.F.; Lambrechts, M.G.; Swiegers, J.H.; Cozzolino, D.; Pretorius, I.S. The effect of increased yeast alcohol acetyltransferase and esterase activity on the flavour profiles of wine and distillates. Yeast 2006, 23, 641–659. [Google Scholar] [CrossRef] [PubMed]
- Sharp, D.C.; Steensels, J.; Shellhammer, T.H. The effect of hopping regime, cultivar and β-glucosidase activity on monoterpene alcohol concentrations in wort and beer. J. Inst. Brew. 2017, 123, 185–191. [Google Scholar] [CrossRef]
- Sharpe, F.R.; Laws, D.R.J. The essential oil of hops a review. J. Inst. Brew. 1981, 87, 96–107. [Google Scholar] [CrossRef]
- Carrau, M.F.; Medina, K.; Boido, E.; Farina, L.; Gaggero, C.; Dellacassa, E.; Versini, G.; Henschke, A.P. De novo synthesis of monoterpenes by Saccharomyces cerevisiae wine yeasts. FEMS Microbiol. Lett. 2005, 243, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Takoi, K.; Itoga, Y.; Takayanagi, J.; Kosugi, T.; Shioi, T.; Nakamura, T.; Watari, J. Screening of geraniol-rich flavor hop and interesting behavior of β-citronellol during fermentation under various hop-addition timings. J. Am. Soc. Brew. Chem. 2014, 72, 22–29. [Google Scholar] [CrossRef]
- King, A.J.; Dickinson, J.R. Biotransformation of monoterpene alcohols by Saccharomyces cerevisiae, Torulaspora delbrueckii and Kluyveromyces lactis. Yeast 2000, 16, 499–506. [Google Scholar] [CrossRef]
- King, A.J.; Dickinson, J.R. Biotransformation of hop aroma terpenoids by ale and lager yeasts. FEMS Yeast Res. 2003, 3, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Steyer, D.; Erny, C.; Claudel, P.; Riveill, G.; Karst, F.; Legras, J.-L. Genetic analysis of geraniol metabolism during fermentation. Food Microbiol. 2013, 33, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Warburton, A.; Silcock, P.; Bremer, P.J.; Eyres, G.T. Yeast Strain Influences the Hop-Derived Sensory Properties and Volatile Composition of Beer. Foods 2023, 12, 1064. [Google Scholar] [CrossRef] [PubMed]
- Daenen, L.; Saison, D.; Sterckx, F.; Delvaux, F.R.; Verachtert, H.; Derdelinckx, G. Screening and evaluation of the glucoside hydrolase activity in Saccharomyces and Brettanomyces brewing yeasts. J. Appl. Microbiol. 2008, 104, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.; Rainieri, S.; Witte, S.; Matern, U.; Martens, S. Identification of a Saccharomyces cerevisiae Glucosidase That Hydrolyzes Flavonoid Glucosides. Appl. Environ. Microbiol. 2011, 77, 1751–1757. [Google Scholar] [CrossRef]
- Swiegers, J.H.; Pretorius, I.S. Modulation of volatile sulfur compounds by wine yeast. Appl. Microbiol. Biotechnol. 2007, 74, 954–960. [Google Scholar] [CrossRef]
- Subileau, M.; Schneider, R.; Salmon, J.-M.; Degryse, E. Nitrogen catabolite repression modulates the production of aromatic thiols characteristic of Sauvignon Blanc at the level of precursor transport. FEMS Yeast Res. 2008, 8, 771–780. [Google Scholar] [CrossRef] [PubMed]
- Vega, E.A. Variation in Irc7p Activity Amongst Brewing Strains of Saccharomyces cerevisiae. Bachelor’s Thesis, Oregon State University, Corvallis, OR, USA, 2021. [Google Scholar]
- Belda, I.; Ruiz, J.; Navascués, E.; Marquina, D.; Santos, A. Improvement of aromatic thiol release through the selection of yeasts with increased beta-lyase activity. Int. J. Food Microbiol. 2016, 225, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Roncoroni, M.; Santiago, M.; Hooks, D.O.; Moroney, S.; Harsch, M.J.; Lee, S.A.; Richards, K.D.; Nicolau, L.; Gardner, R.C. The yeast IRC7 gene encodes a β-lyase responsible for production of the varietal thiol 4-mercapto-4-methylpentan-2-one in wine. Food Microbiol. 2011, 28, 926–935. [Google Scholar] [CrossRef] [PubMed]
- Santiago, M.; Gardner, R.C. The IRC7 gene encodes cysteine desulphydrase activity and confers on yeast the ability to grow on cysteine as a nitrogen source. Yeast 2015, 32, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, J.; de Celis, M.; Martín-Santamaría, M.; Benito-Vázquez, I.; Pontes, A.; Lanza, V.F.; Sampaio, J.P.; Santos, A.; Belda, I. Global distribution of IRC7 alleles in Saccharomyces cerevisiae populations: A genomic and phenotypic survey within the wine clade. Environ. Microbiol. 2021, 23, 3182–3195. [Google Scholar] [CrossRef] [PubMed]
- Thibon, C.; Marullo, P.; Claisse, O.; Cullin, C.; Dubourdieu, D.; Tominaga, T. Nitrogen catabolic repression controls the release of volatile thiols by Saccharomyces cerevisiae during wine fermentation. FEMS Yeast Res. 2008, 8, 1076–1086. [Google Scholar] [CrossRef]
- Deed, N.K.; van Vuuren, H.J.J.; Gardner, R.C. Effects of nitrogen catabolite repression and di-ammonium phosphate addition during wine fermentation by a commercial strain of S. cerevisiae. Appl. Microbiol. Biotechnol. 2011, 89, 1537–1549. [Google Scholar] [CrossRef]
- Dufour, M.; Zimmer, A.; Thibon, C.; Marullo, P. Enhancement of volatile thiol release of Saccharomyces cerevisiae strains using molecular breeding. Appl. Microbiol. Biotechnol. 2013, 97, 5893–5905. [Google Scholar] [CrossRef] [PubMed]
- Ugliano, M.; Henschke, P.A. Yeasts and wine flavour. In Wine Chemistry and Biochemistry; Springer: New York, NY, USA, 2009; pp. 313–392. [Google Scholar]
- Julia, W.; Martina, G.; Thomas, B. Phenolic Substances in Beer: Structural Diversity, Reactive Potential and Relevance for Brewing Process and Beer Quality. Compr. Rev. Food Sci. Food Saf. 2018, 17, 953–988. [Google Scholar]
- Vanbeneden, N.; Van Roey, T.; Willems, F.; Delvaux, F.; Delvaux, F.R. Release of phenolic flavour precursors during wort production: Influence of process parameters and grist composition on ferulic acid release during brewing. Food Chem. 2008, 111, 83–91. [Google Scholar] [CrossRef]
- Sterckx, F.L.; Missiaen, J.; Saison, D.; Delvaux, F.R. Contribution of monophenols to beer flavour based on flavour thresholds, interactions and recombination experiments. Food Chem. 2011, 126, 1679–1685. [Google Scholar] [CrossRef] [PubMed]
- Vanbeneden, N.; Gils, F.; Delvaux, F.; Delvaux, F.R. Formation of 4-Vinyl and 4-Ethyl Derivatives from Hydroxycinnamic Acids: Occurrence of Volatile Phenolic Flavour Compounds in Beer and Distribution of Pad1-Activity among Brewing Yeasts. Food Chem. 2008, 107, 221–230. [Google Scholar] [CrossRef]
- Lentz, M. The Impact of Simple Phenolic Compounds on Beer Aroma and Flavor. Fermentation 2018, 4, 20. [Google Scholar] [CrossRef]
- White, M.D.; Payne, K.A.P.; Fisher, K.; Marshall, S.A.; Parker, D.; Rattray, N.J.W.; Trivedi, D.K.; Goodacre, R.; Rigby, S.E.J.; Scrutton, N.S.; et al. UbiX is a flavin prenyltransferase required for bacterial ubiquinone biosynthesis. Nature 2015, 522, 502–506. [Google Scholar] [CrossRef] [PubMed]
- Mukai, N.; Masaki, K.; Fujii, T.; Kawamukai, M.; Iefuji, H. PAD1 and FDC1 are essential for the decarboxylation of phenylacrylic acids in Saccharomyces cerevisiae. J. Biosci. Bioeng. 2010, 109, 564–569. [Google Scholar] [CrossRef] [PubMed]
- Gallone, B.; Mertens, S.; Gordon, J.L.; Maere, S.; Verstrepen, K.J.; Steensels, J. Origins, evolution, domestication and diversity of Saccharomyces beer yeasts. Curr. Opin. Biotechnol. 2018, 49, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Kerruish, D.W.; Cormican, P.; Kenny, E.M.; Kearns, J.; Colgan, E.; Boulton, C.A.; Stelma, S.N. The origins of the Guinness stout yeast. Commun. Biol. 2024, 7, 68. [Google Scholar] [CrossRef] [PubMed]
- Michel, M.; Meier-Dörnberg, T.; Jacob, F.; Methner, F.J.; Wagner, R.S.; Hutzler, M. Review: Pure Non-Saccharomyces Starter Cultures for Beer Fermentation with a Focus on Secondary Metabolites and Practical Applications. J. Inst. Brew. 2016, 122, 569–587. [Google Scholar] [CrossRef]
- Catallo, M.; Nikulin, J.; Johansson, L.; Krogerus, K.; Laitinen, M.; Magalhães, F.; Piironen, M.; Mikkelson, A.; Randazzo, C.L.; Solieri, L.; et al. Sourdough derived strains of Saccharomyces cerevisiae and their potential for farmhouse ale brewing. J. Inst. Brew. 2020, 126, 168–175. [Google Scholar] [CrossRef]
- Tataridis, P.; Drosou, F.; Kanellis, A.; Kechagia, D.; Logothetis, S.; Chatzilazarou, A.; Dourtoglou, V. Differentiating beer aroma, flavor and alcohol content through the use of Torulaspora delbrueckii. In Proceedings of the Young Scientists Symposium for Brewing, Malting and Distilling, Chico, CA, USA, 21–23 April 2016; pp. 21–23. [Google Scholar]
- Gibson, B.; Krogerus, K.; Ekberg, J.; Mikkelson, A.; Pentikäinen, S.; Wilpola, A.; Vidgren, V. Non-conventional Yeast as a New Tool for Beer Flavour Modification. In Proceedings of the EBC Congress, Porto, Portugal, 24–28 May 2015. [Google Scholar]
- Canonico, L.; Agarbati, A.; Comitini, F.; Ciani, M. Torulaspora delbrueckii in the brewing process: A new approach to enhance bioflavour and to reduce ethanol content. Food Microbiol. 2016, 56, 45–51. [Google Scholar] [CrossRef]
- Steensels, J.; Daenen, L.; Malcorps, P.; Derdelinckx, G.; Verachtert, H.; Verstrepen, K.J. Brettanomyces yeasts—From spoilage organisms to valuable contributors to industrial fermentations. Int. J. Food Microbiol. 2015, 206, 24–38. [Google Scholar] [CrossRef]
- Blomqvist, J.; Eberhard, T.; Schnürer, J.; Passoth, V. Fermentation Characteristics of Dekkera Bruxellensis Strains. Appl. Microbiol. Biotechnol. 2010, 87, 1487–1497. [Google Scholar] [CrossRef] [PubMed]
- Harrouard, J.; Eberlein, C.; Ballestra, P.; Dols-Lafargue, M.; Masneuf-Pomarede, I.; Miot-Sertier, C.; Schacherer, J.; Albertin, W. Brettanomyces bruxellensis: Overview of the genetic and phenotypic diversity of an anthropized yeast. Mol. Ecol. 2023, 32, 2374–2395. [Google Scholar] [CrossRef] [PubMed]
- Colomer, M.S.; Funch, B.; Forster, J. The Raise of Brettanomyces Yeast Species for Beer Production. Curr. Opin. Biotechnol. 2019, 56, 30–35. [Google Scholar] [CrossRef]
- Pirrone, A.; Prestianni, R.; Naselli, V.; Todaro, A.; Farina, V.; Tinebra, I.; Raffaele, G.; Badalamenti, N.; Maggio, A.; Gaglio, R.; et al. Influence of indigenous Hanseniaspora uvarum and Saccharomyces cerevisiae from sugar-rich substrates on the aromatic composition of loquat beer. Int. J. Food Microbiol. 2022, 379, 109868. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.Z.; Liu, X.Z.; Li, X.; Sheng, X.F.; Li, T.T.; Tang, W.Y.; Yu, Z.H.; Wang, Y.M. Effect of Hanseniaspora uvarum-Saccharomyces cerevisiae mixed fermentation on aroma characteristics of Rosa roxburghii Tratt, Blueberry, and Plum Wines. Molecules 2022, 27, 8097. [Google Scholar] [CrossRef] [PubMed]
- De Francesco, G.; Turchetti, B.; Sileoni, V.; Marconi, O.; Perretti, G. Screening of new strains of Saccharomycodes ludwigii and Zygosaccharomyces rouxii to produce low-alcohol beer. J. Inst. Brew. 2015, 121, 113–121. [Google Scholar] [CrossRef]
- Sohrabvandi, S.; Razavi, S.H.; Mousavi, S.M.; Amir, M. Viability of probiotic bacteria in low alcohol- and non- alcoholic beer during refrigerated storage. Philipp. Agric. Sci. 2010, 93, 24–28. [Google Scholar]
- Methner, Y.; Hutzler, M.; Matoulková, D.; Jacob, F.; Michel, M. Screening for the Brewing Ability of Different Non-Saccharomyces Yeasts. Fermentation 2019, 5, 101. [Google Scholar] [CrossRef]
- Nieto-Sarabia, V.L.; Melgar-Lalanne, G.; Ballinas-Cesatti, C.B.; García-García, F.A.; Jose-Salazar, J.A.; Flores-Ortiz, C.M.; Cristiani-Urbina, E.; Morales-Barrera, L. Brewing a Craft Belgian-Style Pale Ale Using Pichia kudriavzevii 4A as a Starter Culture. Microorganisms 2023, 11, 977. [Google Scholar] [CrossRef]
- Bhatia, S.K.; Bhatia, R.K.; Choi, Y.-K.; Kan, E.; Kim, Y.-G.; Yang, Y.-H. Biotechnological potential of microbial consortia and future perspectives. Crit. Rev. Biotechnol. 2018, 38, 1209–1229. [Google Scholar] [CrossRef] [PubMed]
- Rapp, K.M.; Jenkins, J.P.; Betenbaugh, M.J. Partners for life: Building microbial consortia for the future. Curr. Opin. Biotechnol. 2020, 66, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Mittermeier, F.; Bäumler, M.; Arulrajah, P.; de Jesus Garcia Lima, J.; Hauke, S.; Stock, A.; Weuster-Botz, D. Artificial microbial consortia for bioproduction processes. Eng. Life Sci. 2023, 23, e2100152. [Google Scholar] [CrossRef]
- Giudici, P.; Solieri, L.; Pulvirenti, A.M.; Cassanelli, S. Strategies and perspectives for genetic improvement of wine yeasts. Appl. Microbiol. Biotechnol. 2005, 66, 622–628. [Google Scholar] [CrossRef]
- Steensels, J.L.; Meersman, E.; Snoek, T.; Saels, V.; Verstrepen, K.J. Large-scale selection and breeding to generate industrial yeasts with superior aroma production. Appl. Environ. Microbiol. 2014, 80, 6965–6975. [Google Scholar] [CrossRef]
- Gibson, B.; Dahabieh, M.; Krogerus, K.; Jouhten, P.; Magalhães, F.; Pereira, R.; Siewers, V.; Vidgren, V. Adaptive Laboratory Evolution of Ale and Lager Yeasts for Improved Brewing Efficiency and Beer Quality. Annu. Rev. Food Sci. Technol. 2020, 11, 23–44. [Google Scholar] [CrossRef] [PubMed]
- DiCarlo, J.E.; Conley, A.J.; Penttila, M.; Jantti, J.; Wang, H.H.; Church, G.M. Yeast Oligo-Mediated Genome Engineering (YOGE). ACS Synth. Biol. 2013, 2, 741–749. [Google Scholar] [CrossRef]
- Barbieri, E.M.; Muir, P.; Akhuetie-Oni, B.O.; Yellman, C.M.; Isaacs, F.J. Precise Editing at DNA Replication Forks Enables Multiplex Genome Engineering in Eukaryotes. Cell 2017, 171, 1453–1467. [Google Scholar] [CrossRef]
- Zeng, W.; Guo, L.; Xu, S.; Chen, J.; Zhou, J. High-throughput screening technology in industrial biotechnology. Trends Biotechnol. 2020, 38, 888–906. [Google Scholar] [CrossRef]
- Diderich, J.A.; Weening, S.M.; Van den Broek, M.; Pronk, J.T.; Daran, J.G. Selection of Pof− Saccharomyces eubayanus variants for the construction of S. cerevisiae × S. eubayanus hybrids with reduced 4-vinyl guaiacol formation. Front. Microbiol. 2018, 9, 1640. [Google Scholar] [CrossRef]
- Woolston, B.M.; Edgar, S.; Stephanopoulos, G. Metabolic Engineering: Past and Future. Annu. Rev. Chem. Biomol. Eng. 2013, 4, 259–288. [Google Scholar] [CrossRef] [PubMed]
- Gibson, D.J.; Young, L.; Chuang, R.-Y.; Venter, J.C.; Hutchison, C.A.; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef] [PubMed]
- Casini, A.; Storch, M.; Baldwin, G.; Ellis, T. Bricks and blueprints: Methods and standards for DNA assembly. Nat. Rev. Mol. Cell Biol. 2015, 16, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.E.; DeLoache, W.C.; Cervantes, B.; Dueber, J.E. A Highly Characterized Yeast Toolkit for Modular, Multipart Assembly. ACS Synth. Biol. 2015, 4, 975–986. [Google Scholar] [CrossRef] [PubMed]
- Jakočiūnas, T.; Bonde, I.; Herrgård, M.; Harrison, S.J.; Kristensen, M.; Pedersen, L.E.; Jensen, M.K.; Keasling, J.D. Multiplex metabolic pathway engineering using CRISPR/Cas9 in Saccharomyces cerevisiae. Metab. Eng. 2015, 28, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Dahabieh, M.S.; Thevelein, J.M.; Gibson, B. Multimodal Microorganism Development: Integrating Top-Down Biological Engineering with Bottom-Up Rational Design. Trends Biotechnol. 2020, 38, 241–253. [Google Scholar] [CrossRef]
- de Vries, G.A.R.; de Groot, P.A.; van den Broek, M.; Daran, J.G. CRISPR-Cas9 mediated gene deletions in lager yeast Saccharomyces pastorianus. Microb. Cell Factories 2017, 16, 222:1–222:18. [Google Scholar]
- Mertens, S.; Gallone, B.; Steensels, J.; Herrera-Malaver, B.; Cortebeek, J.; Nolmans, R.; Saels, V.; Vyas, V.K.; Verstrepen, K.J. Reducing phenolic off-flavors through CRISPR-based gene editing of the FDC1 gene in Saccharomyces cerevisiae x Saccharomyces eubayanus hybrid lager beer yeasts. PLoS ONE 2019, 14, e0224525. [Google Scholar]
- Raschmanová, H.; Weninger, A.; Glieder, A.; Kovar, K.; Vogl, T. Implementing CRISPR-Cas technologies in conventional and non-conventional yeasts: Current state and future prospects. Biotechnol. Adv. 2018, 36, 641–665. [Google Scholar] [CrossRef]
- Krogerus, K.; Fletcher, E.; Rettberg, N.; Gibson, B.; Preiss, R. Efficient Breeding of Industrial Brewing Yeast Strains Using CRISPR/Cas9-Aided Mating-Type Switching. Appl. Microbiol. Biotechnol. 2021, 105, 8359–8376. [Google Scholar]
- Kiene, F.; Pretorius, I.S.; Rauhut, D.; von Wallbrunn, C.; van Wyk, N. Construction and analysis of a yeast for the simultaneous release and esterification of the varietal thiol 3-sulfanylhexan-1-ol. J. Agric. Food Chem. 2021, 69, 11919–11925. [Google Scholar] [CrossRef] [PubMed]
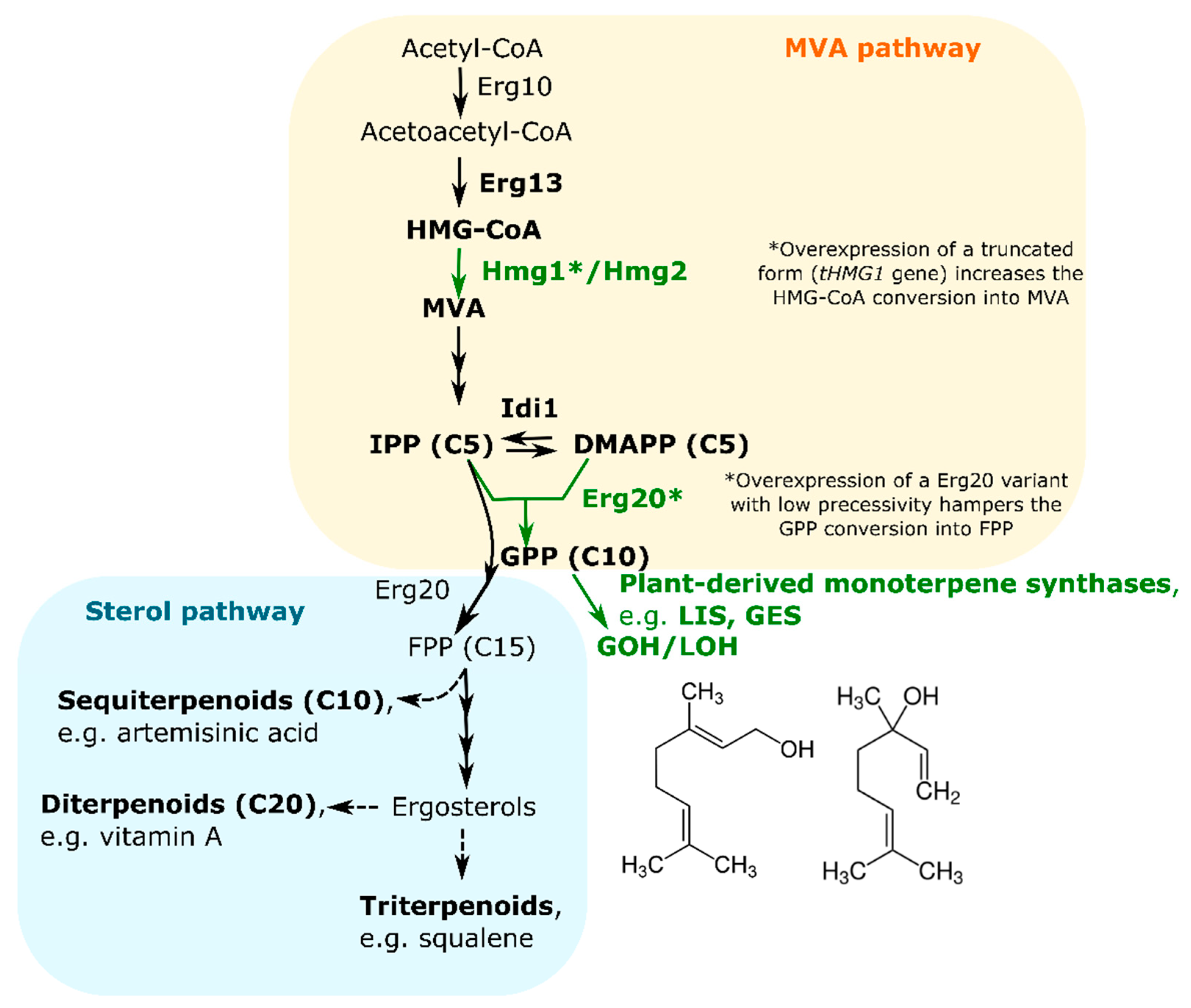
- Burg, J.S.; Espenshade, P.J. Regulation of HMG-CoA reductase in mammals and yeast. Prog. Lipid Res. 2011, 50, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Ignea, C.; Trikka, F.A.; Nikolaidis, A.K.; Georgantea, P.; Ioannou, E.; Loupassaki, S.; Kefalas, P.; Kanellis, A.K.; Roussis, V.; Makris, A.M.; et al. Efficient diterpene production in yeast by engineering Erg20p into a geranylgeranyl diphosphate synthase. Metab. Eng. 2015, 27, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Polakowski, T.; Stahl, U.; Lang, C. Overexpression of a cytosolic hydroxymethylglutaryl-CoA reductase leads to squalene accumulation in yeast. Appl. Microbiol. Biotechnol. 1998, 49, 66–71. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Class | Compound (IUPAC Name) | Aroma Attributes | Aroma Threshold |
|---|---|---|---|
| Acetate esters | Ethyl acetate | Fruity, solvent | 30 mg/L |
| Isoamyl acetate | Banana, apple, solvent | 1.2–2 mg/L | |
| 2-Phenylethyl acetate | Roses, honey, apple, sweetish | 0.2–3.8 mg/L | |
| MCFA 1 ethyl esters | Ethyl butyrate | Papaya, butter, sweetish, apple | 0.4 mg/L |
| Ethyl decanoate | Fatty acids, apple, solvent | 1.5 mg/L | |
| Ethyl hexanoate | Apple, fruity | 0.2–0.23 mg/L | |
| Ethyl octanoate | Apple, aniseed | 0.9–1.0 mg/L | |
| Higher alcohols | n-Propanol (Propan-1-ol) | Alcohol | 600 mg/L |
| isobutyl alcohol (2-methylpropan-1-ol) | Solvent | 100 mg/L | |
| Isoamyl alcohol (3-Methylbutan-1-ol) | Alcohol, banana | 50–65 mg/L | |
| 2-Methyl-1-butanol or active amyl alcohol (2-Methylbutan-1-ol) | Alcohol, solvent | 50–70 mg/L | |
| 2-Phenylethanol | Floral, rose | 40 mg/L | |
| Monoterpene alcohols | Geraniol | Floral, roses, fruity, citrus | 6 µg/L |
| Nerol | Lemon, fruity | 500 µg/L | |
| Linallol | Floral, citrus | 3–5 µg/L | |
| β-Citronellol | Floral, fruity, citrus | 8 µg/L | |
| Thiols and their esters | 3-mercaptohexan-1-ol | Passion fruit, grape fruit, gooseberry, guava | 55–60 ng/L |
| 3-mercaptohexyl acetate | Passion fruit, grape fruit, box tree, gooseberry, guava | 4 ng/L | |
| 4-mercapto-4-methylpentan-2-one | Box tree, passion fruit, broom, black currant | 1.5 ng/L |
| Gene | Functions | References |
|---|---|---|
| TIR1 | Cell wall mannoprotein for maintaining the integrity of the cell wall | [38,39] |
| GAP1 | Amino acid transporter | [38] |
| BAT1 | Transaminase which transfers the amino groups between branched-chain amino acids and α-keto acids | [40,41] |
| BAT2 | Transaminase which transfers the amino groups between branched-chain amino acids and α-keto acids | [40,41] |
| PDC1 | Decarboxylase which removes the carboxyl group from an α-keto acid | [42] |
| PDC5 | Decarboxylase which removes the carboxyl group from an α-keto acid | [43] |
| ARO10 | Decarboxylase which removes the carboxyl group from an α-keto acid | [44] |
| ADH1 | Alcohol dehydrogenase which reduces aldehyde to alcohol using NADH as a cofactor | [45,46] |
| ARO4 | 3-deoxy-7-phosphoheptulonate synthase which synthesizes a tyrosine precursor | [47] |
| Transporter | Function | Family |
|---|---|---|
| Gap1 | General AA transporter and nutrient sensor | Plasma membrane transporter |
| Agp1 | Transporter of all AAs except for arginine and lysine | |
| Agp2 | AA transporter and sensor that regulates the expression of importers for carnitine and polyamines | |
| Bap2 | Leucine transporter | |
| Gnp1 | High affinity glutamine, cysteine, and proline permease | |
| Put4 | Proline, alanine, and glycine transporter | |
| Lyp1 | Lysine transporter | |
| Alp1 | Arginine transporter | |
| Avt3 | General neutral AA vacuolar exporter | Vacuolar transporter |
| Avt4 | General neutral AA vacuolar exporter | |
| Avt7 | Vacuolar exporter of glutamine and proline | |
| Vba5 | Plasma membrane transporter for arginine and lysine | |
| Agc1 | Aspartate/glutamate carrier | Mitochondrial transporter |
| Ort1 | Ornithine, arginine, and lysine carrier | |
| Uga4 | Vacuolar membrane protein | Other transporter |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nasuti, C.; Solieri, L. Yeast Bioflavoring in Beer: Complexity Decoded and Built up Again. Fermentation 2024, 10, 183. https://doi.org/10.3390/fermentation10040183
Nasuti C, Solieri L. Yeast Bioflavoring in Beer: Complexity Decoded and Built up Again. Fermentation. 2024; 10(4):183. https://doi.org/10.3390/fermentation10040183
Chicago/Turabian StyleNasuti, Chiara, and Lisa Solieri. 2024. "Yeast Bioflavoring in Beer: Complexity Decoded and Built up Again" Fermentation 10, no. 4: 183. https://doi.org/10.3390/fermentation10040183
APA StyleNasuti, C., & Solieri, L. (2024). Yeast Bioflavoring in Beer: Complexity Decoded and Built up Again. Fermentation, 10(4), 183. https://doi.org/10.3390/fermentation10040183