
Efficient Secretory Expression for Mammalian Hemoglobins in Pichia pastoris

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents, Strains, and Culture Conditions
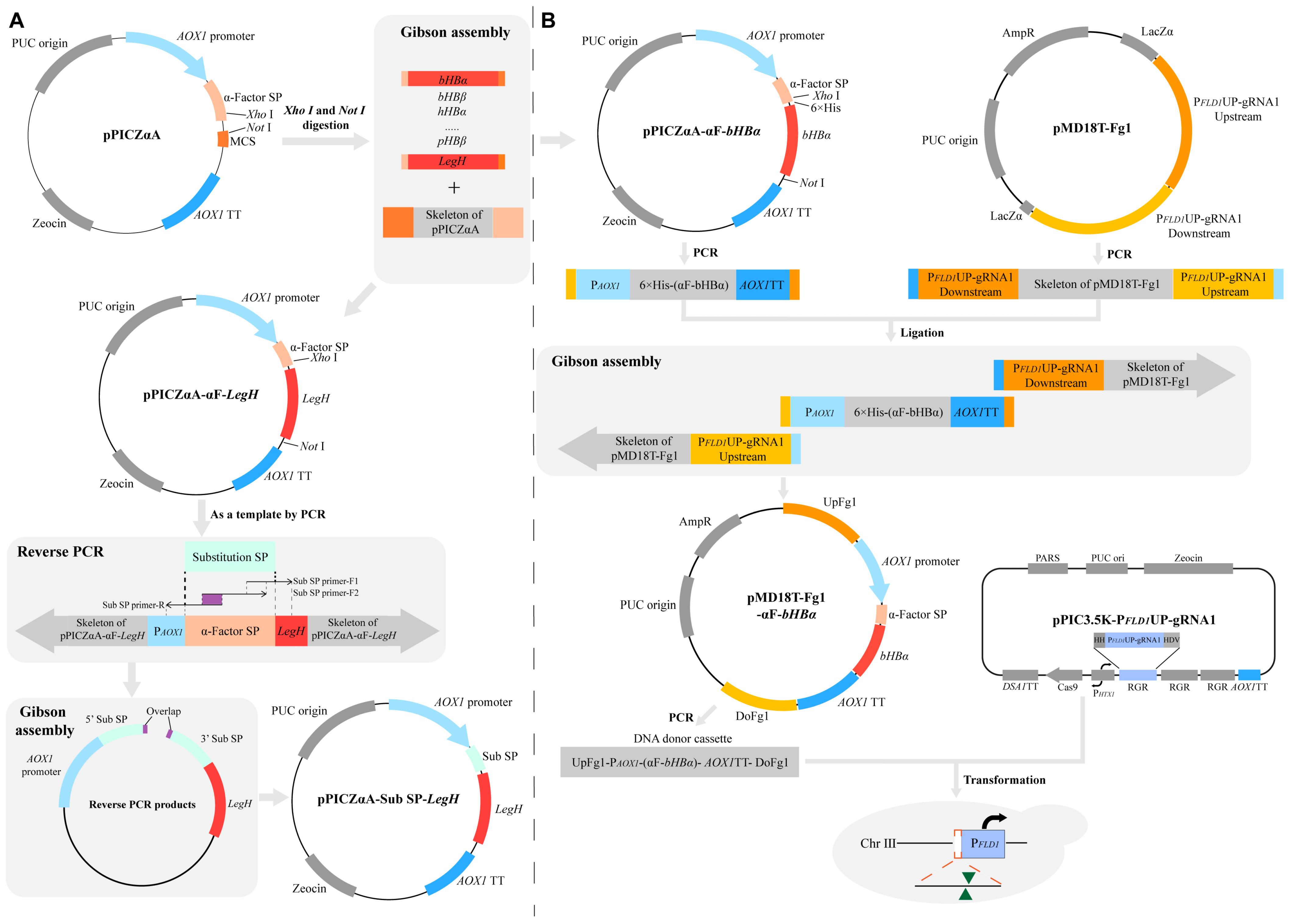
2.2. The Construction of Recombinant Plasmids
2.3. The Construction of Engineered Pichia pastoris Strains
2.4. Cultural Conditions for Hemoglobin Expression at Shaking-Flask Level
2.5. Cultural Conditions for Hemoglobin Expression at 5-L Fermenter Level
2.6. Quantification and Purification for Hemoglobins
2.7. Determination of Hemoglobin Functional Activity and Protein Sequence
2.8. Statistical Analysis
3. Results
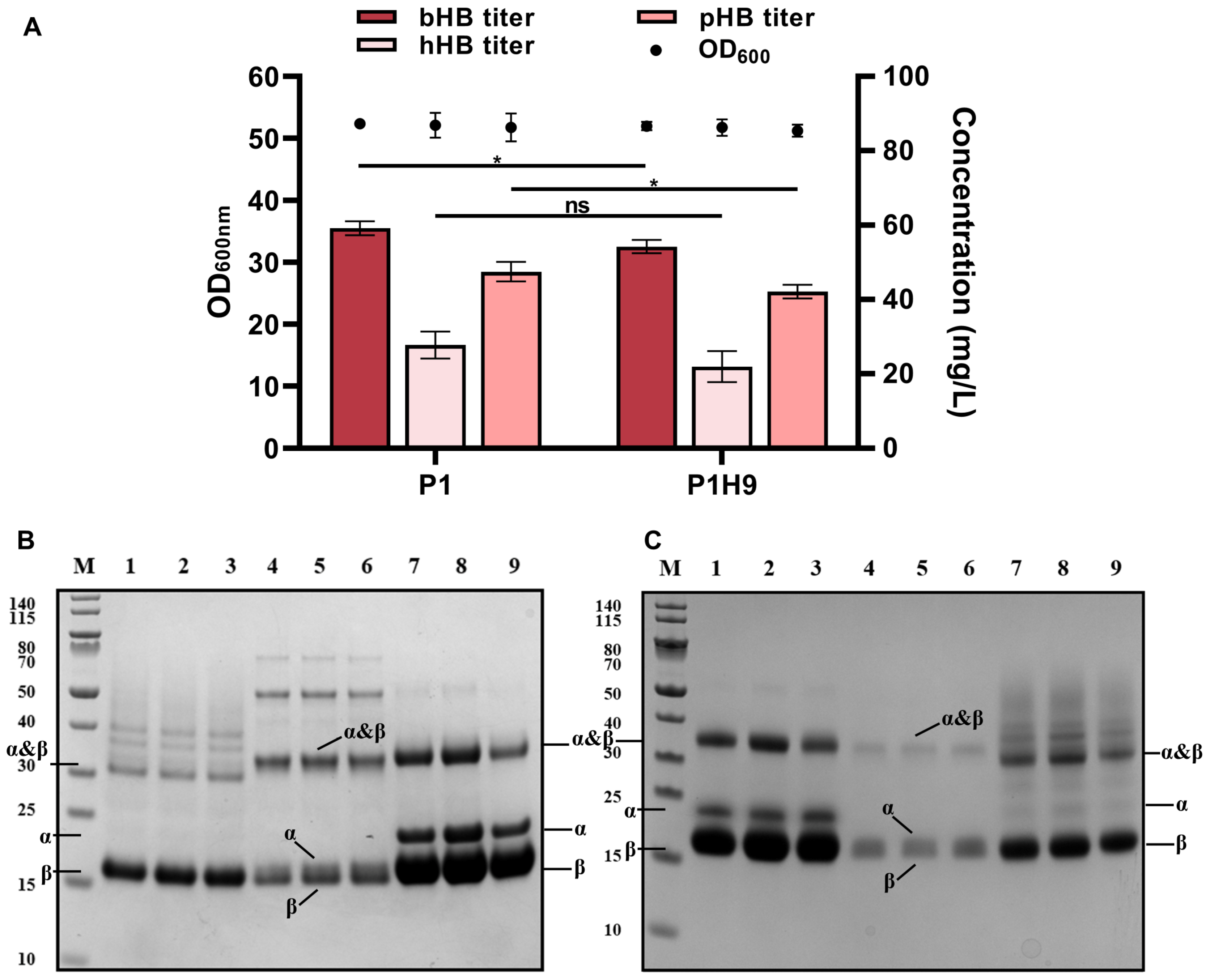
3.1. Selection of an Efficient System to Express Globin Components in HB
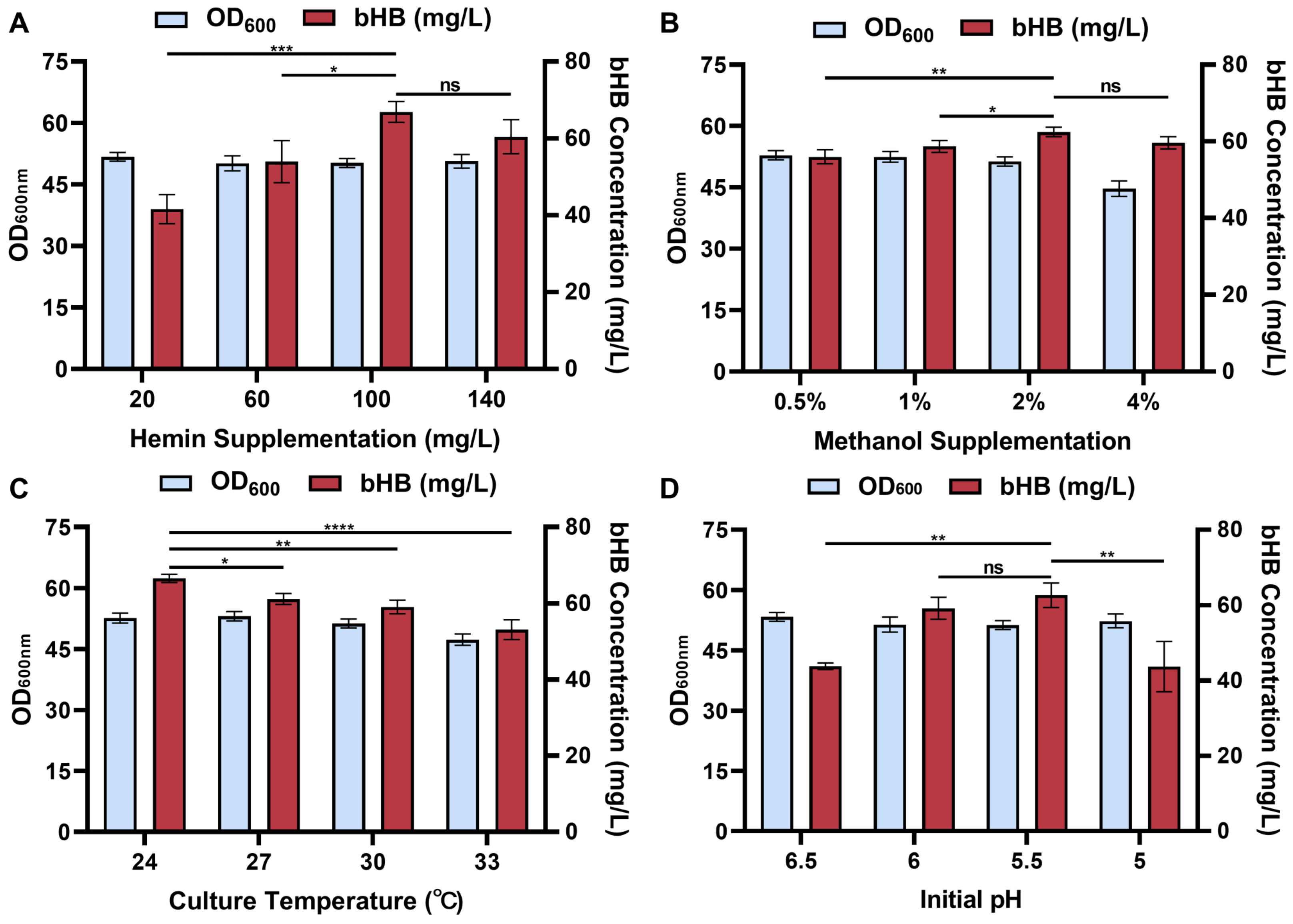
3.2. Optimization of Fermentation Conditions at Shaking-Flask Level
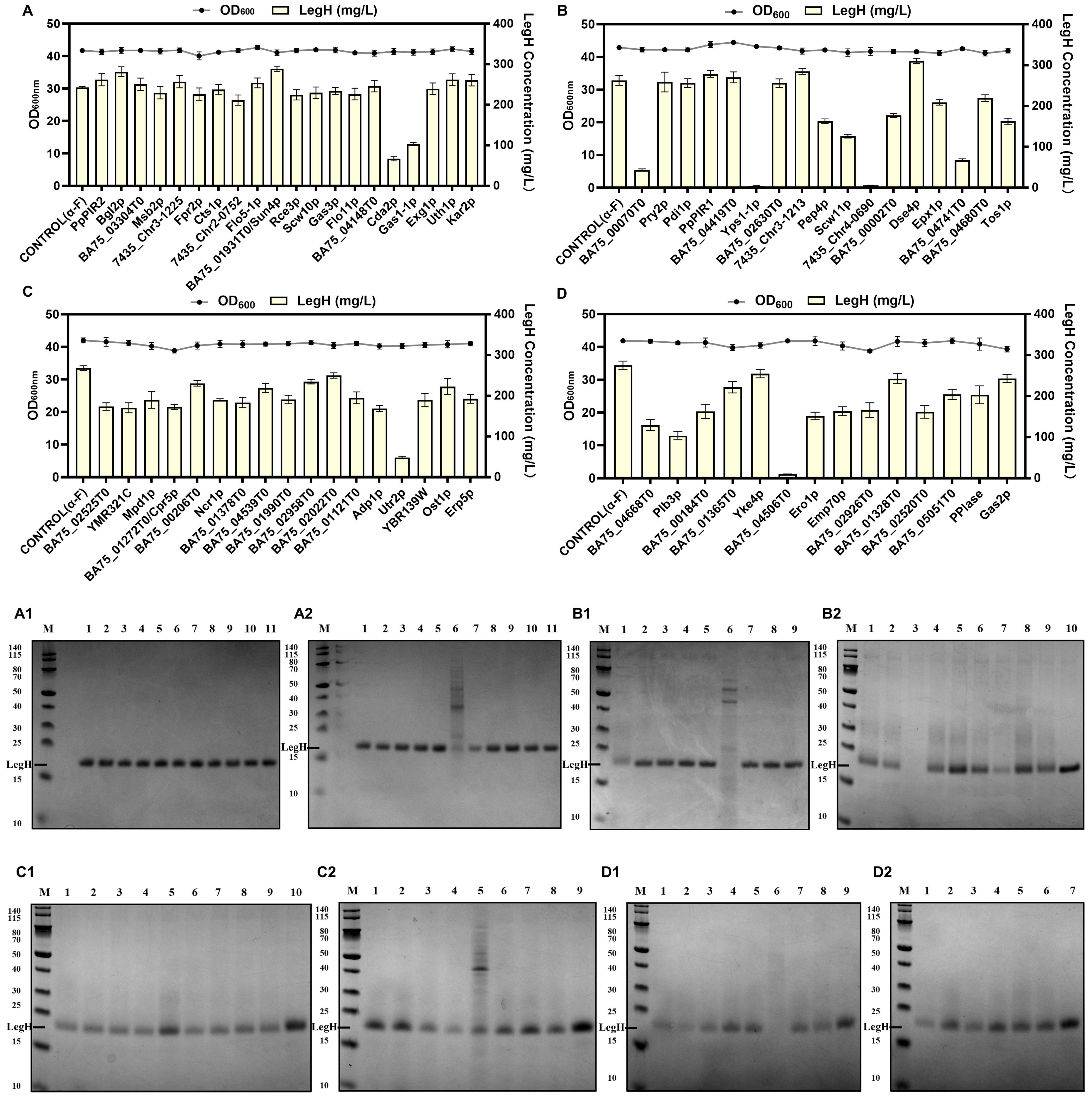
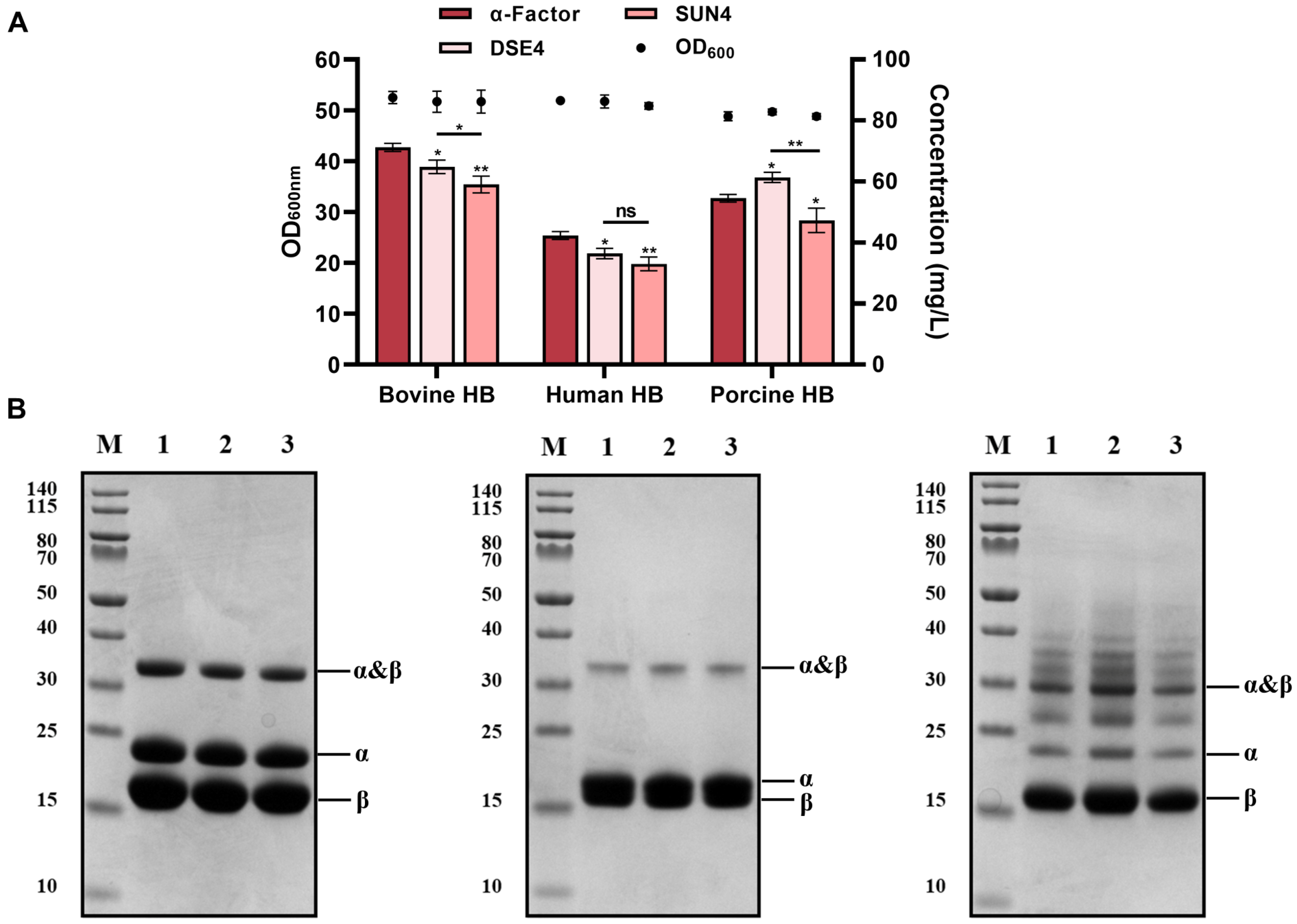
3.3. Screening Efficient Endogenous Signal Peptides in Pichia pastoris
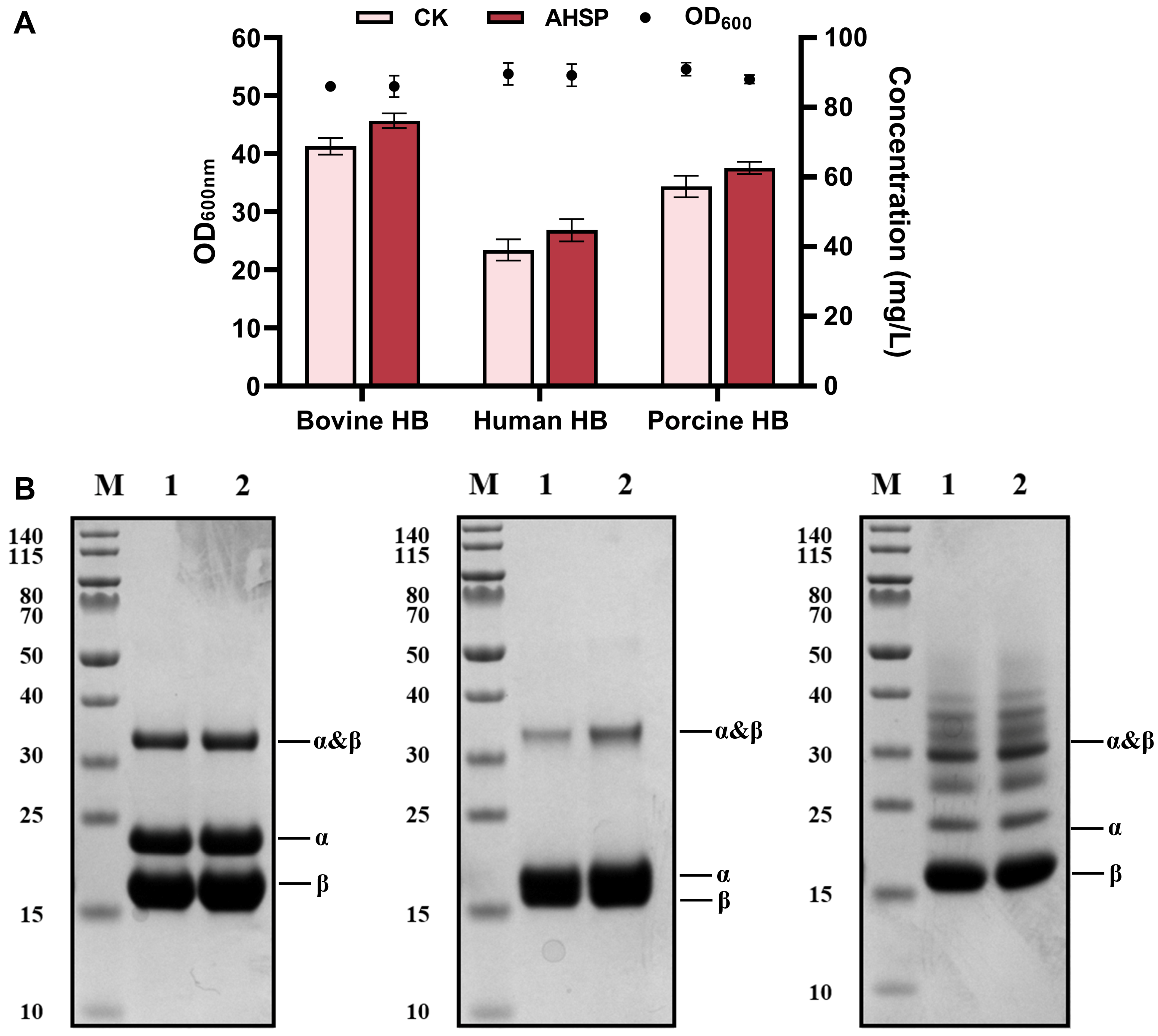
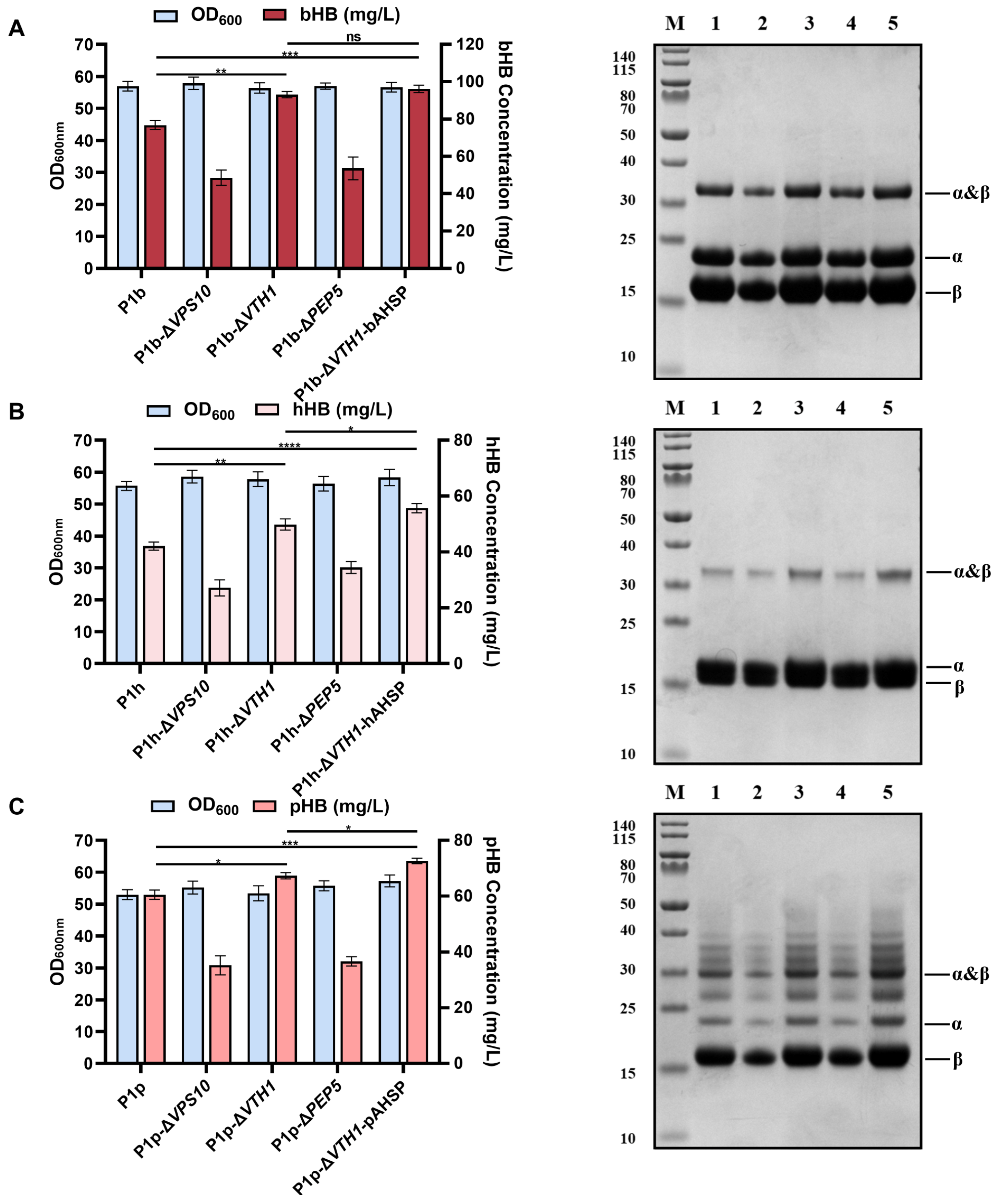
3.4. Enhancing the Synthesis of Mammalian Hemoglobins through Combinatorial Metabolic Engineering Strategies
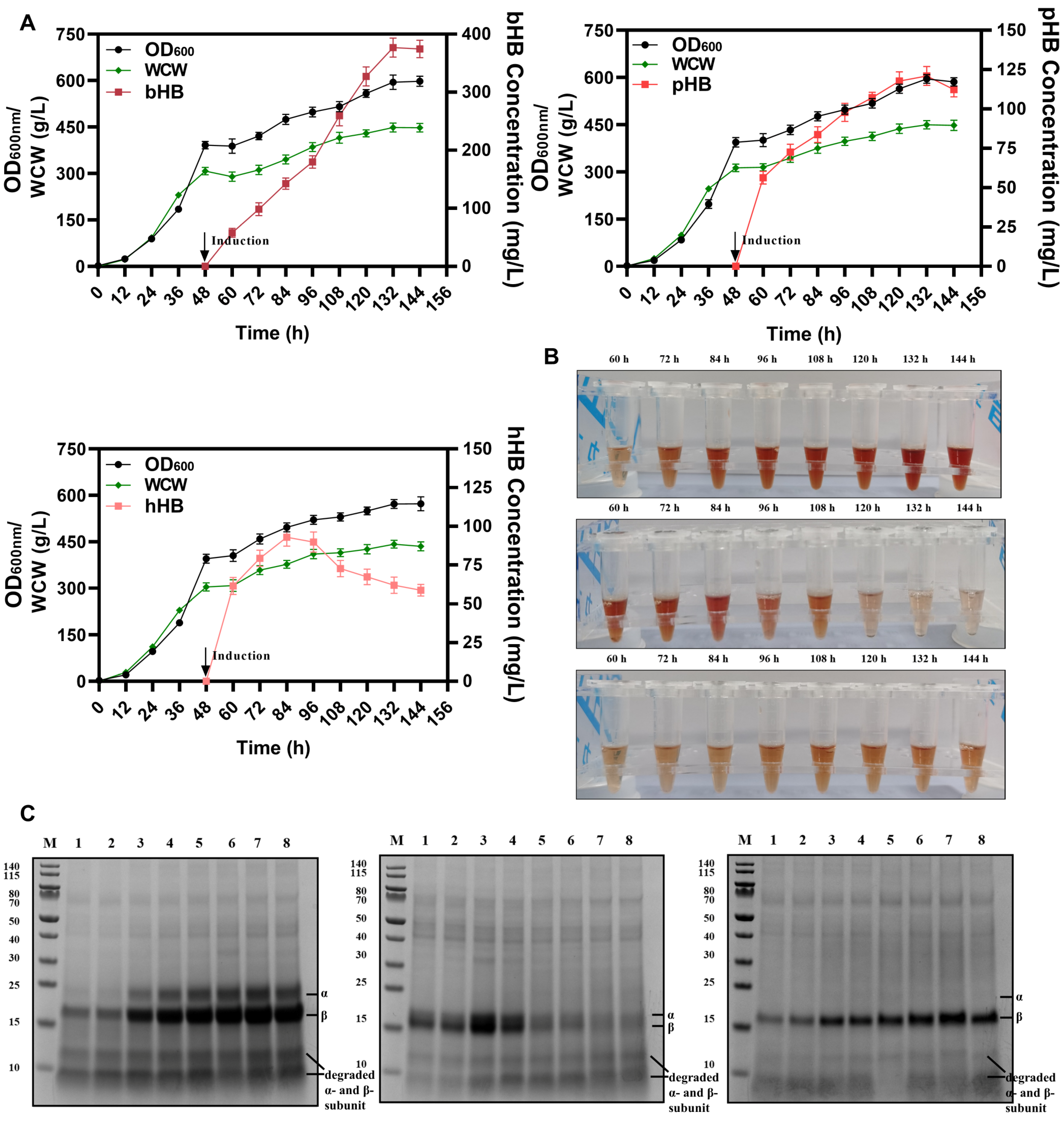
3.5. Secretory Expression of Mammalian Hemoglobins at 5-L Fermenter Level
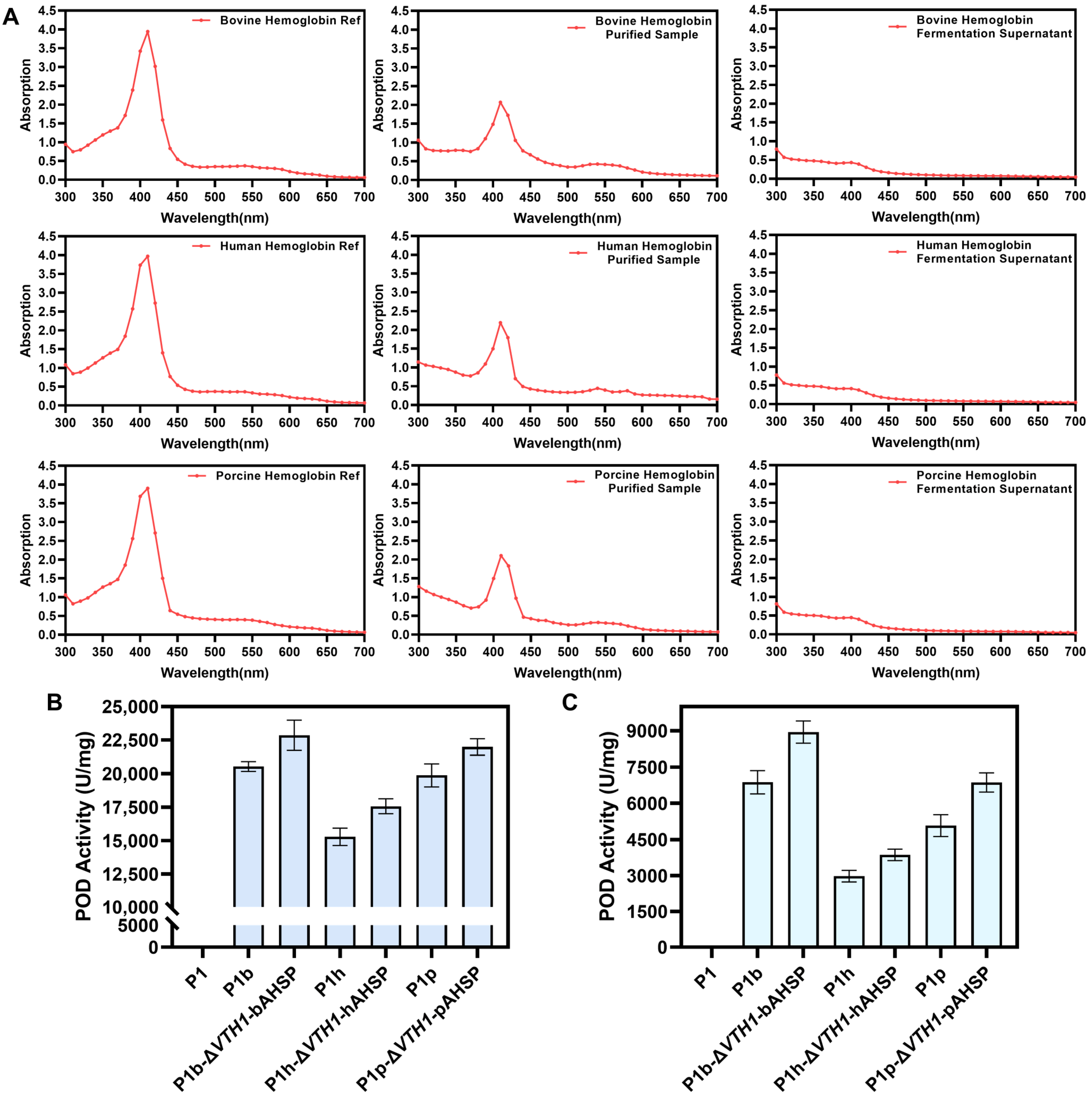
3.6. Biochemical Properties of Various Mammalian Hemoglobins Synthesized in Engineered Pichia pastoris Strains
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yuan, Y.; Tam, M.F.; Simplaceanu, V.; Ho, C. New look at hemoglobin allostery. Chem. Rev. 2015, 115, 1702–1724. [Google Scholar] [CrossRef] [PubMed]
- Gell, D.A. Structure and function of haemoglobins. Blood Cells Mol. Dis. 2018, 70, 13–42. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, K.; Hazato, T. Isolation and identification of an endogenous inhibitor of enkephalin-degrading enzymes from bovine spinal cord. Biochem. Biophys. Res. Commun. 1993, 194, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Di Caprio, G.; Schonbrun, E.; Gonçalves, B.P.; Valdez, J.M.; Wood, D.K.; Higgins, J.M. High-throughput assessment of hemoglobin polymer in single red blood cells from sickle cell patients under controlled oxygen tension. Proc. Natl. Acad. Sci. USA 2019, 116, 25236–25242. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.; Carter, E.L.; Harland, J.B.; Speelman, A.L.; Lehnert, N.; Ragsdale, S.W. Ferric heme as a CO/NO sensor in the nuclear receptor Rev-Erbß by coupling gas binding to electron transfer. Proc. Natl. Acad. Sci. USA 2021, 118, e2016717118. [Google Scholar] [CrossRef] [PubMed]
- Giardina, B. Hemoglobin: Multiple molecular interactions and multiple functions. An example of energy optimization and global molecular organization. Mol. Aspects Med. 2022, 84, 101040. [Google Scholar] [CrossRef]
- Tupta, B.; Stuehr, E.; Sumi, M.P.; Sweeny, E.A.; Smith, B.; Stuehr, D.J.; Ghosh, A. GAPDH is involved in the heme-maturation of myoglobin and hemoglobin. FASEB J. 2022, 36, e22099. [Google Scholar] [CrossRef]
- Bruice, P.Y. Organic Chemistry; Pearson: London, UK, 2014. [Google Scholar]
- Faggiano, S.; Ronda, L.; Bruno, S.; Abbruzzetti, S.; Viappiani, C.; Bettati, S.; Mozzarelli, A. From hemoglobin allostery to hemoglobin-based oxygen carriers. Mol. Aspects Med. 2022, 84, 101050. [Google Scholar] [CrossRef] [PubMed]
- Pizarro, F.; Olivares, M.; Valenzuela, C.; Brito, A.; Weinborn, V.; Flores, S.; Arredondo, M. The effect of proteins from animal source foods on heme iron bioavailability in humans. Food Chem. 2016, 196, 733–738. [Google Scholar] [CrossRef] [PubMed]
- Ikeda-Saito, M.; Raven, E. (Eds.) Dioxygen-Dependent Heme Enzymes; The Royal Society of Chemistry: London, UK, 2018. [Google Scholar]
- Gomes, I.; Dale, C.S.; Casten, K.; Geigner, M.A.; Gozzo, F.C.; Ferro, E.S.; Heimann, A.S.; Devi, L.A. Hemoglobin-derived peptides as novel type of bioactive signaling molecules. AAPS J. 2010, 12, 658–669. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Wu, Q.; Song, S.; She, R.; Zhao, Y.; Yang, Y.; Zhang, M.; Du, F.; Soomro, M.H.; Shi, R. Antimicrobial activity and safety evaluation of peptides isolated from the hemoglobin of chickens. BMC Microbiol. 2016, 16, 287. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; He, X.; Andoh-Kumi, K.; Fraser, R.Z.; Lu, M.; Goodman, R.E. Evaluating potential risks of food allergy and toxicity of soy leghemoglobin expressed in Pichia pastoris. Mol. Nutr. Food Res. 2018, 62, 1700297. [Google Scholar] [CrossRef] [PubMed]
- Rubio, N.R.; Xiang, N.; Kaplan, D.L. Plant-based and cell-based approaches to meat production. Nat. Commun. 2020, 11, 6276. [Google Scholar] [CrossRef] [PubMed]
- Simsa, R.; Yuen, J.; Stout, A.; Rubio, N.; Fogelstrand, P.; Kaplan, D.L. Extracellular heme proteins influence bovine myosatellite cell proliferation and the color of cell-based meat. Foods 2019, 8, 521. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhou, J.; Du, G.; Chen, J. Recent advances in the microbial synthesis of hemoglobin. Trends Biotechnol. 2021, 39, 286–297. [Google Scholar] [CrossRef] [PubMed]
- Fraser, R.Z.; Shitut, M.; Agrawal, P.; Mendes, O.; Klapholz, S. Safety evaluation of soy leghemoglobin protein preparation derived from Pichia pastoris, intended for use as a flavor catalyst in plant-based meat. Int. J. Toxicol. 2018, 37, 241–262. [Google Scholar] [CrossRef] [PubMed]
- Shu, Y.; Cheng, D.; Chen, X.; Wang, J. A reverse microemulsion of water/AOT/1-butyl-3-methylimidazolium hexafluorophosphate for selective extraction of hemoglobin. Sep. Purif. Technol. 2008, 64, 154–159. [Google Scholar] [CrossRef]
- Jia, Y.; Xu, X.; Ou, J.; Liu, X. Solid-phase extraction of hemoglobin from human whole blood with a coordination-polymer-derived composite material based on ZnO and mesoporous carbon. Chem. Eur. J. 2017, 23, 16026–16033. [Google Scholar] [CrossRef] [PubMed]
- Stephanopoulos, G. Synthetic biology and metabolic engineering. ACS Synth. Biol. 2012, 1, 514–525. [Google Scholar] [CrossRef]
- Hoffman, S.J.; Looker, D.L.; Roehrich, J.M.; Cozart, P.E.; Durfee, S.L.; Tedesco, J.L.; Stetler, G.L. Expression of fully functional tetrameric human hemoglobin in Escherichia coli. Proc. Natl. Acad. Sci. USA 1990, 87, 8521–8525. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.; Wang, X.; Bai, Y.; Wang, Y.; Wang, Y.; Tu, T.; Qin, X.; Su, X.; Luo, H.; Yao, B.; et al. Engineering Escherichia coli for efficient assembly of heme proteins. Microb. Cell Fact. 2023, 22, 59. [Google Scholar] [CrossRef]
- Wagenbach, M.; O’Rourke, K.; Vitez, L.; Wieczorek, A.; Hoffman, S.; Durfee, S.; Tedesco, J.; Stetler, G. Synthesis of wild type and mutant human hemoglobins in Saccharomyces cerevisiae. Nat. Biotechnol. 1991, 9, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Ishchuk, O.P.; Frost, A.T.; Muñiz-Paredes, F.; Matsumoto, S.; Laforge, N.; Eriksson, N.L.; Martínez, J.L.; Petranovic, D. Improved production of human hemoglobin in yeast by engineering hemoglobin degradation. Metab. Eng. 2021, 66, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Zhou, J.; Li, J.; Du, G.; Chen, J.; Wang, M.; Zhao, X. Systematic engineering of Saccharomyces cerevisiae for efficient synthesis of hemoglobins and myoglobins. Bioresour. Technol. 2023, 370, 556. [Google Scholar] [CrossRef] [PubMed]
- Baghban, R.; Farajnia, S.; Rajabibazl, M.; Ghasemi, Y.; Mafi, A.; Hoseinpoor, R.; Rahbarnia, L.; Aria, M. Yeast expression systems: Overview and recent advances. Mol. Biotechnol. 2019, 61, 365–384. [Google Scholar] [CrossRef] [PubMed]
- Werten, M.W.T.; Eggink, G.; Cohen Stuart, M.A.; de Wolf, F.A. Production of protein-based polymers in Pichia pastoris. Biotechnol. Adv. 2019, 37, 642–666. [Google Scholar] [CrossRef] [PubMed]
- Cos, O.; Ramón, R.; Montesinos, J.L.; Valero, F. Operational strategies, monitoring and control of heterologous protein production in the methylotrophic yeast Pichia pastoris under different promoters: A review. Microb. Cell Fact. 2006, 5, 17. [Google Scholar] [CrossRef]
- Ergün, B.G.; Laçın, K.; Çaloğlu, B.; Binay, B. Second generation Pichia pastoris strain and bioprocess designs. Biotechnol. Biofuels 2022, 15, 150. [Google Scholar] [CrossRef]
- Schwarzhans, J.-P.; Luttermann, T.; Geier, M.; Kalinowski, J.; Friehs, K. Towards systems metabolic engineering in Pichia pastoris. Biotechnol. Adv. 2017, 35, 681–710. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Zhang, Z. Engineering strategies for enhanced production of protein and bio-products in Pichia pastoris: A review. Biotechnol. Adv. 2018, 36, 182–195. [Google Scholar] [CrossRef] [PubMed]
- Owji, H.; Nezafat, N.; Negahdaripour, M.; Hajiebrahimi, A.; Ghasemi, Y. A comprehensive review of signal peptides: Structure, roles, and applications. Eur. J. Cell Biol. 2018, 97, 422–441. [Google Scholar] [CrossRef] [PubMed]
- Aw, R.; McKay, P.F.; Shattock, R.J.; Polizzi, K.M. A systematic analysis of the expression of the anti-HIV VRC01 antibody in Pichia pastoris through signal peptide optimization. Protein Expr. Purif. 2018, 149, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Li, C.; Ye, Y.; Lin, Y. Endogenous signal peptides efficiently mediate the secretion of recombinant proteins in Pichia pastoris. Biotechnol. Lett. 2013, 35, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Duan, G.; Ding, L.; Wei, D.; Zhou, H.; Chu, J.; Zhang, S.; Qian, J. Screening endogenous signal peptides and protein folding factors to promote the secretory expression of heterologous proteins in Pichia pastoris. J. Biotechnol. 2019, 306, 193–202. [Google Scholar] [CrossRef]
- Vadhana, A.K.P.; Samuel, P.; Berin, R.M.; Krishna, J.; Kamatchi, K.; Meenakshisundaram, S. Improved secretion of Candida antarctica lipase B with its native signal peptide in Pichia pastoris. Enzyme Microb. Technol. 2013, 52, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Zou, C.; Lu, L.; Wang, S.; Zhang, C.; Chen, X.; Lin, Y.; Huang, Y. The α-mating factor secretion signals and endogenous signal peptides for recombinant protein secretion in Komagataella phaffii. Biotechnol. Biofuels 2022, 15, 140. [Google Scholar] [CrossRef]
- Xue, S.; Liu, X.; Pan, Y.; Xiao, C.; Feng, Y.; Zheng, L.; Zhao, M.; Huang, M. Comprehensive analysis of signal peptides in Saccharomyces cerevisiae reveals features for efficient secretion. Adv. Sci. 2023, 10, 2203433. [Google Scholar] [CrossRef]
- De Schutter, K.; Lin, Y.-C.; Tiels, P.; Van Hecke, A.; Glinka, S.; Weber-Lehmann, J.; Rouzé, P.; Van de Peer, Y.; Callewaert, N. Genome sequence of the recombinant protein production host Pichia pastoris. Nat. Biotechnol. 2009, 27, 561–566. [Google Scholar] [CrossRef]
- Hong, E.; Davidson, A.R.; Kaiser, C.A. A pathway for targeting soluble misfolded proteins to the yeast vacuole. J. Cell Biol. 1996, 135, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Shen, Y.; Hou, J.; Peng, B.; Tang, H.; Bao, X. Secretory pathway engineering enhances secretion of cellobiohydrolase I from Trichoderma reesei in Saccharomyces cerevisiae. J. Biosci. Bioeng. 2014, 117, 45–52. [Google Scholar] [CrossRef]
- Iwaki, T.; Hosomi, A.; Tokudomi, S.; Kusunoki, Y.; Fujita, Y.; Giga-Hama, Y.; Tanaka, N.; Takegawa, K. Vacuolar protein sorting receptor in Schizosaccharomyces pombe. Microbiology 2006, 152, 1523–1532. [Google Scholar] [CrossRef] [PubMed]
- Seals, D.F.; Eitzen, G.; Margolis, N.; Wickner, W.T.; Price, A. A Ypt/Rab effector complex containing the Sec1 homolog Vps33p is required for homotypic vacuole fusion. Proc. Natl. Acad. Sci. USA 2000, 97, 9402–9407. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.; Woolford, C.A.; Jones, E.W. Pep3p/Pep5p complex: A putative docking factor at multiple steps of vesicular transport to the vacuole of Saccharomyces cerevisiae. Genetics 2000, 156, 105–122. [Google Scholar] [CrossRef] [PubMed]
- Stroupe, C.; Collins, K.M.; Fratti, R.A.; Wickner, W. Purification of active HOPS complex reveals its affinities for phosphoinositides and the SNARE Vam7p. EMBO J. 2006, 25, 1579–1589. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Zhao, X.; Zhou, J.; Lu, W.; Li, J.; Chen, J.; Du, G. Biosynthesis of high-active hemoproteins by the efficient heme-supply Pichia pastoris chassis. Adv. Sci. 2023, 10, 2302826. [Google Scholar] [CrossRef] [PubMed]
- Labun, K.; Montague, T.G.; Krause, M.; Torres Cleuren, Y.N.; Tjeldnes, H.; Valen, E. CHOPCHOP v3: Expanding the CRISPR web toolbox beyond genome editing. Nucleic Acids Res. 2019, 47, W171–W174. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Shi, X.; Song, L.; Liu, H.; Zhou, X.; Wang, Q.; Zhang, Y.; Cai, M. CRISPR–Cas9-mediated genomic multiloci integration in Pichia pastoris. Microb. Cell Fact. 2019, 18, 144. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zhao, X.; Wang, Z.; Wang, H.; Zhou, J.; Du, G.; Chen, J.; Li, J. Efficient secretory expression and purification of food-grade porcine myoglobin in Komagataella phaffii. J. Agric. Food Chem. 2021, 69, 10235–10245. [Google Scholar] [CrossRef] [PubMed]
- Li, P.Z.; Anumanthan, A.; Gao, X.G.; Ilangovan, K.; Suzara, V.V.; Düzgünes, N.; Renugopalakrishnan, V. Expression of recombinant proteins in Pichia pastoris. Appl. Biochem. Biotechnol. 2007, 142, 105–124. [Google Scholar] [CrossRef] [PubMed]
- Rakestraw, J.A.; Sazinsky, S.L.; Piatesi, A.; Antipov, E.; Wittrup, K.D. Directed evolution of a secretory leader for the improved expression of heterologous proteins and full-length antibodies in Saccharomyces cerevisiae. Biotechnol. Bioeng. 2009, 103, 1192–1201. [Google Scholar] [CrossRef] [PubMed]
- Mateljak, I.; Tron, T.; Alcalde, M. Evolved α-factor prepro-leaders for directed laccase evolution in Saccharomyces cerevisiae. Microb. Biotechnol. 2017, 10, 1830–1836. [Google Scholar] [CrossRef]
- Agrawal, G.K.; Jwa, N.-S.; Lebrun, M.-H.; Job, D.; Rakwal, R. Plant secretome: Unlocking secrets of the secreted proteins. Proteomics 2010, 10, 799–827. [Google Scholar] [CrossRef] [PubMed]
- Burgard, J.; Grünwald-Gruber, C.; Altmann, F.; Zanghellini, J.; Valli, M.; Mattanovich, D.; Gasser, B. The secretome of Pichia pastoris in fed-batch cultivations is largely independent of the carbon source but changes quantitatively over cultivation time. Microb. Biotechnol. 2020, 13, 479–494. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Xue, C.; Liu, W.; Zuo, S.; Wei, P.; Huang, L.; Lian, J.; Xu, Z. High-level secretory production of leghemoglobin in Pichia pastoris through enhanced globin expression and heme biosynthesis. Bioresour. Technol. 2022, 363, 884. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Gell, D.A.; Zhou, S.; Gu, L.; Kong, Y.; Li, J.; Hu, M.; Yan, N.; Lee, C.; Rich, A.M.; et al. Molecular mechanism of AHSP-mediated stabilization of α-hemoglobin. Cell 2004, 119, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Mollan, T.L.; Yu, X.; Weiss, M.J.; Olson, J.S. The role of alpha-hemoglobin stabilizing protein in redox chemistry, denaturation, and hemoglobin assembly. Antioxid. Redox Signal. 2009, 12, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Roth, G.; Vanz, A.L.; Lünsdorf, H.; Nimtz, M.; Rinas, U. Fate of the UPR marker protein Kar2/Bip and autophagic processes in fed-batch cultures of secretory insulin precursor producing Pichia pastoris. Microb. Cell Fact. 2018, 17, 123. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Rao, S.; Li, H.; Wu, C.; Wang, Q.; Li, T.; Huang, A. Enhancing HSA-GCSFm fusion protein production by Pichia pastoris with an on-line model-based exponential and DO-stat control modes. Biochem. Eng. J. 2022, 177, 108262. [Google Scholar] [CrossRef]
- Zhang, W.; Inan, M.; Meagher, M.M. Rational design and optimization of fed-batch and continuous fermentations. Methods Mol. Biol. 2007, 389, 43–63. [Google Scholar] [PubMed]
- Kosmachevskaya, O.V.; Nasybullina, E.I.; Shumaev, K.B.; Topunov, A.F. Expressed soybean leghemoglobin: Effect on Escherichia coli at oxidative and nitrosative stress. Molecules 2021, 26, 7207. [Google Scholar] [CrossRef] [PubMed]
- Falkner, J.A.; Kachman, M.; Veine, D.M.; Walker, A.; Strahler, J.R.; Andrews, P.C. Validated MALDI-TOF/TOF mass spectra for protein standards. J. Am. Soc. Mass Spectrom. 2007, 18, 850–855. [Google Scholar] [CrossRef] [PubMed]
- Anwised, P.; Jangpromma, N.; Temsiripong, T.; Patramanon, R.; Daduang, S.; Jitrapakdee, S.; Araki, T.; Klaynongsruang, S. Cloning, expression, and characterization of Siamese Crocodile (Crocodylus siamensis) hemoglobin from Escherichia coli and Pichia pastoris. Protein J. 2016, 35, 256–268. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, C.; Jiang, X.; Fago, A.; Weber, R.E.; Moriyama, H.; Storz, J.F. Expression and purification of recombinant hemoglobin in Escherichia coli. PLoS ONE 2011, 6, e20176. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.J.; Ho, N.T.; Simplaceanu, V.; Zou, M.; Green, B.N.; Tam, M.F.; Ho, C. Production of unmodified human adult hemoglobin in Escherichia coli. Proc. Natl. Acad. Sci. USA 1993, 90, 8108–8112. [Google Scholar] [CrossRef] [PubMed]
- Belcher, D.A.; Lucas, A.; Cabrales, P.; Palmer, A.F. Polymerized human hemoglobin facilitated modulation of tumor oxygenation is dependent on tumor oxygenation status and oxygen affinity of the hemoglobin-based oxygen carrier. Sci. Rep. 2020, 10, 11372. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sources | Strategy | Titer | Refs. |
|---|---|---|---|
| E. coli as host | |||
| Crocodylus siamensis | Intracellular inducible expression by vector | 3.0 mg/L (shaking-flask level) | [64] |
| Peromyscus maniculatus | Intracellular inducible expression by vector | / | [65] |
| Homo sapiens | Intracellular inducible expression by vector | 20% of total protein (shaking-flask level) | [66] |
| S. cerevisiae as host | |||
| H. sapiens | Intracellular integrated expression by vector; improving heme biosynthesis; reducing heme and hemoglobin degradation | 18% of total protein (shaking-flask level) | [25] |
| Bos taurus | Intracellular inducible expression by vector; heme synthetic pathway engineering | 25.2 ± 1.9 mg/L (shaking-flask level) | [26] |
| Sus scrofa | 16.7 ± 0.5 mg/L (shaking-flask level) | ||
| P. pastoris as host | |||
| C. siamensis | Extracellular inducible integrated expression | 6.0 mg/L (shaking-flask level) | [64] |
| B. taurus | Extracellular inducible integrated expression; signal peptide screening; knock-out of vacuolar protease related genes; fermentation conditions optimization; | 96.1 ± 1.6 mg/L (shaking-flask level) 376.9 ± 13.3 mg/L (5-L fermenter) | This study |
| H. sapiens | 55.7 ± 1.4 mg/L (shaking-flask level) 101.1 ± 6.7 mg/L (5-L fermenter) | ||
| S. scrofa | 72.6 ± 0.5 mg/L (shaking-flask level) 119.2 ± 7.3 mg/L (5-L fermenter) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Zhang, T.; Luo, Z.; Zhou, J.; Li, J.; Chen, J.; Du, G.; Zhao, X. Efficient Secretory Expression for Mammalian Hemoglobins in Pichia pastoris. Fermentation 2024, 10, 208. https://doi.org/10.3390/fermentation10040208
Li C, Zhang T, Luo Z, Zhou J, Li J, Chen J, Du G, Zhao X. Efficient Secretory Expression for Mammalian Hemoglobins in Pichia pastoris. Fermentation. 2024; 10(4):208. https://doi.org/10.3390/fermentation10040208
Chicago/Turabian StyleLi, Chenyang, Tao Zhang, Zhengshan Luo, Jingwen Zhou, Jianghua Li, Jian Chen, Guocheng Du, and Xinrui Zhao. 2024. "Efficient Secretory Expression for Mammalian Hemoglobins in Pichia pastoris" Fermentation 10, no. 4: 208. https://doi.org/10.3390/fermentation10040208
APA StyleLi, C., Zhang, T., Luo, Z., Zhou, J., Li, J., Chen, J., Du, G., & Zhao, X. (2024). Efficient Secretory Expression for Mammalian Hemoglobins in Pichia pastoris. Fermentation, 10(4), 208. https://doi.org/10.3390/fermentation10040208