
The Fermentation Quality, Antioxidant Activity, and Bacterial Community of Mulberry Leaf Silage with Pediococcus, Bacillus, and Wheat Bran

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain Activation
2.2. Silage Preparation
2.3. Determination of Chemical Composition
2.4. Fermentation Parameters Detection and Analysis
2.5. Microbiological Evaluation
2.6. Determination of Active Substance and Antioxidant Activity
2.7. Analysis of Bacterial Community
2.8. Statistical Analysis
3. Results
3.1. The Chemical Composition and Microbial Population of Mulberry Leaves before Ensiling
3.2. Chemical Composition, Fermentation Quality and Microbial Population of Mulberry Leaf Silage
3.3. Active Components and Antioxidant Activity of Fresh Mulberry Leaves and Mulberry Leaf Silage
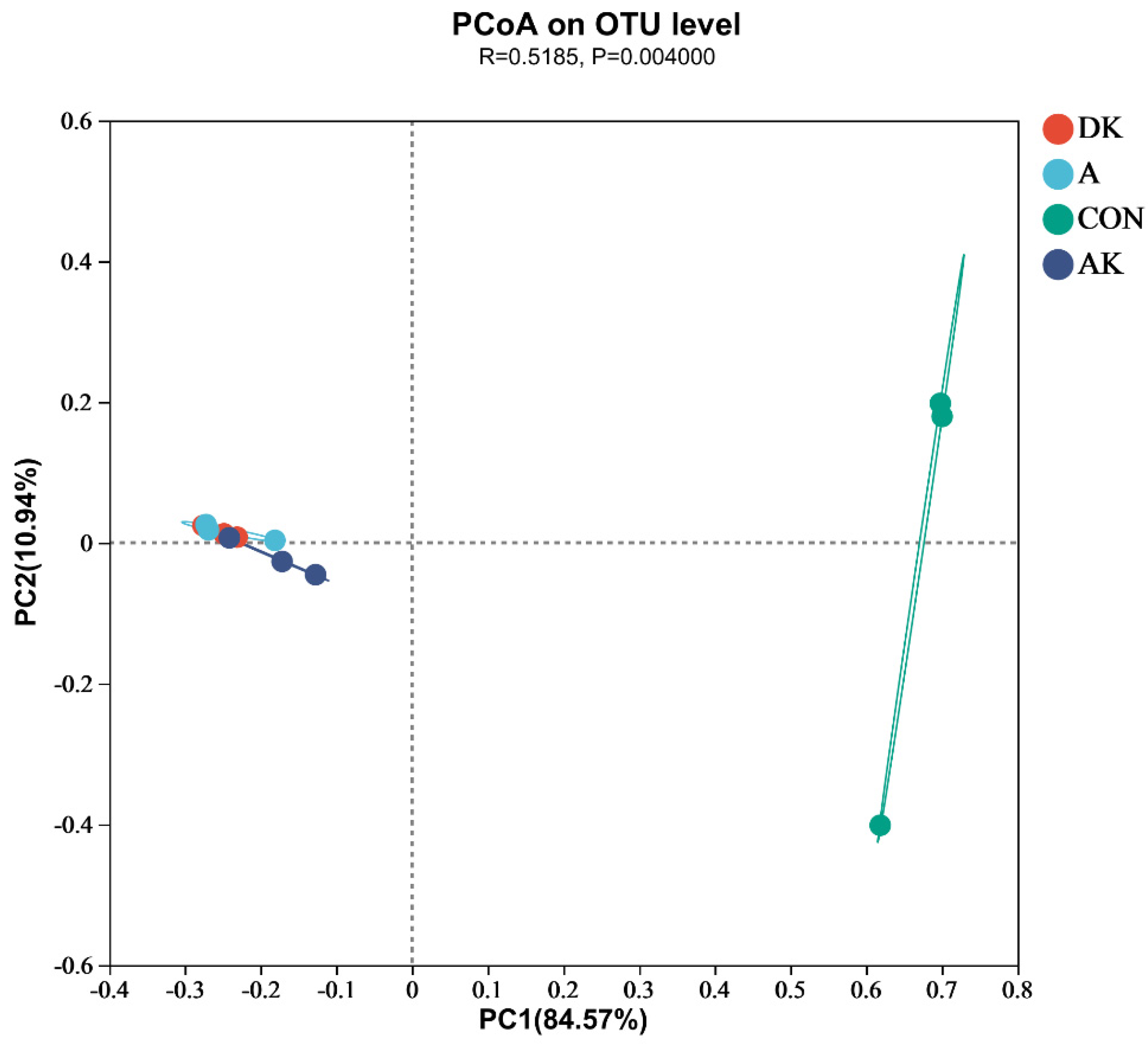
3.4. Bacterial Diversity of Fresh Mulberry Leaves and Mulberry Leaf Silage
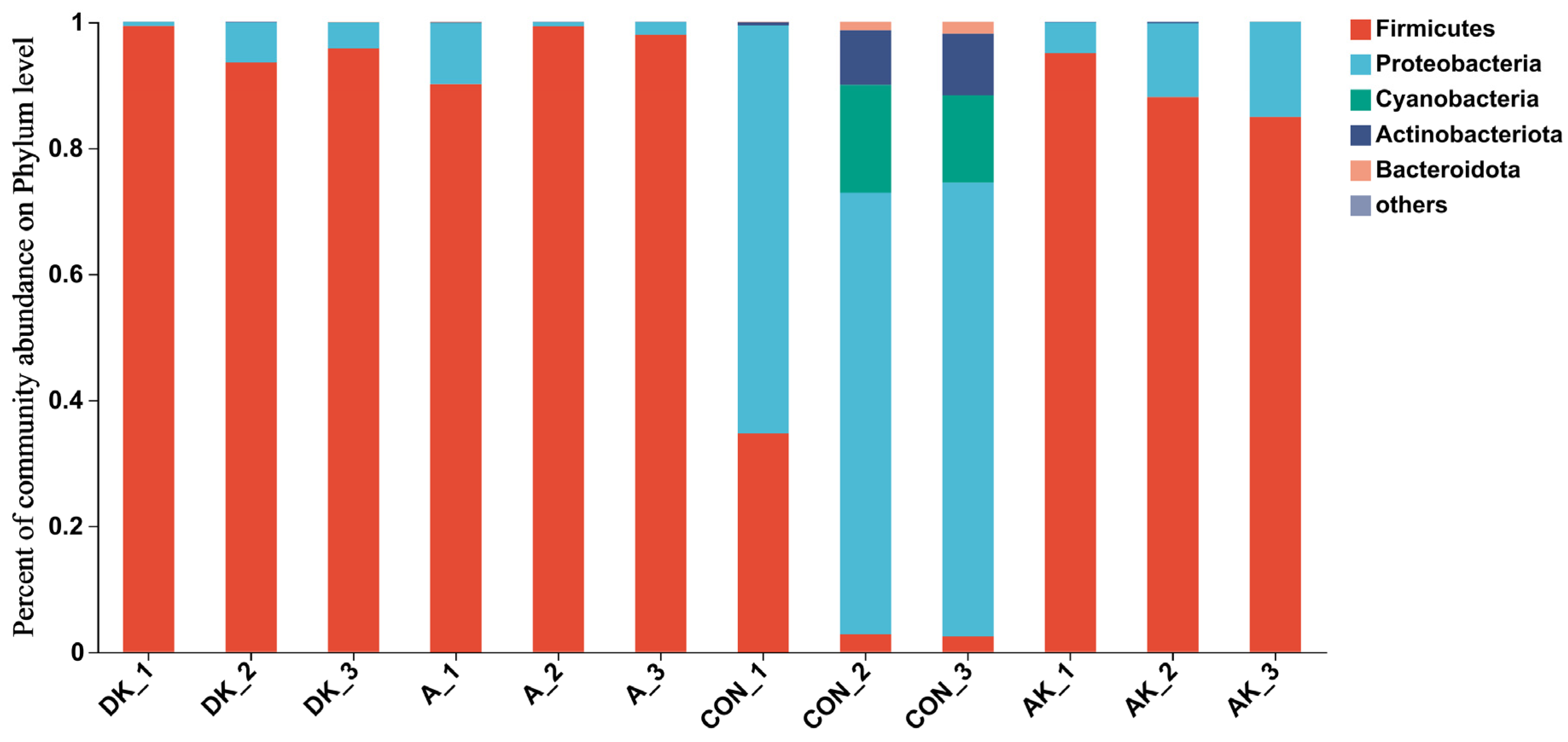
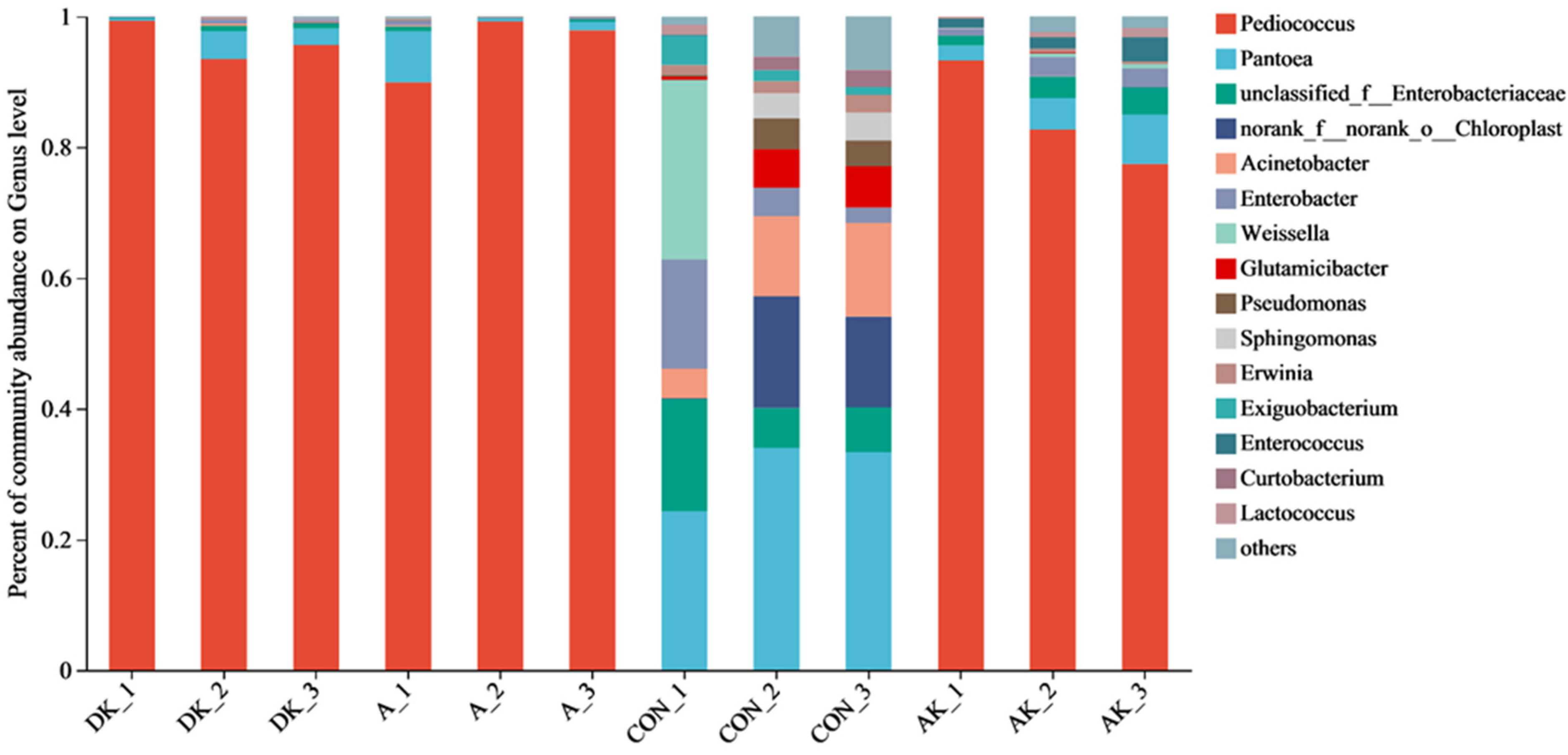
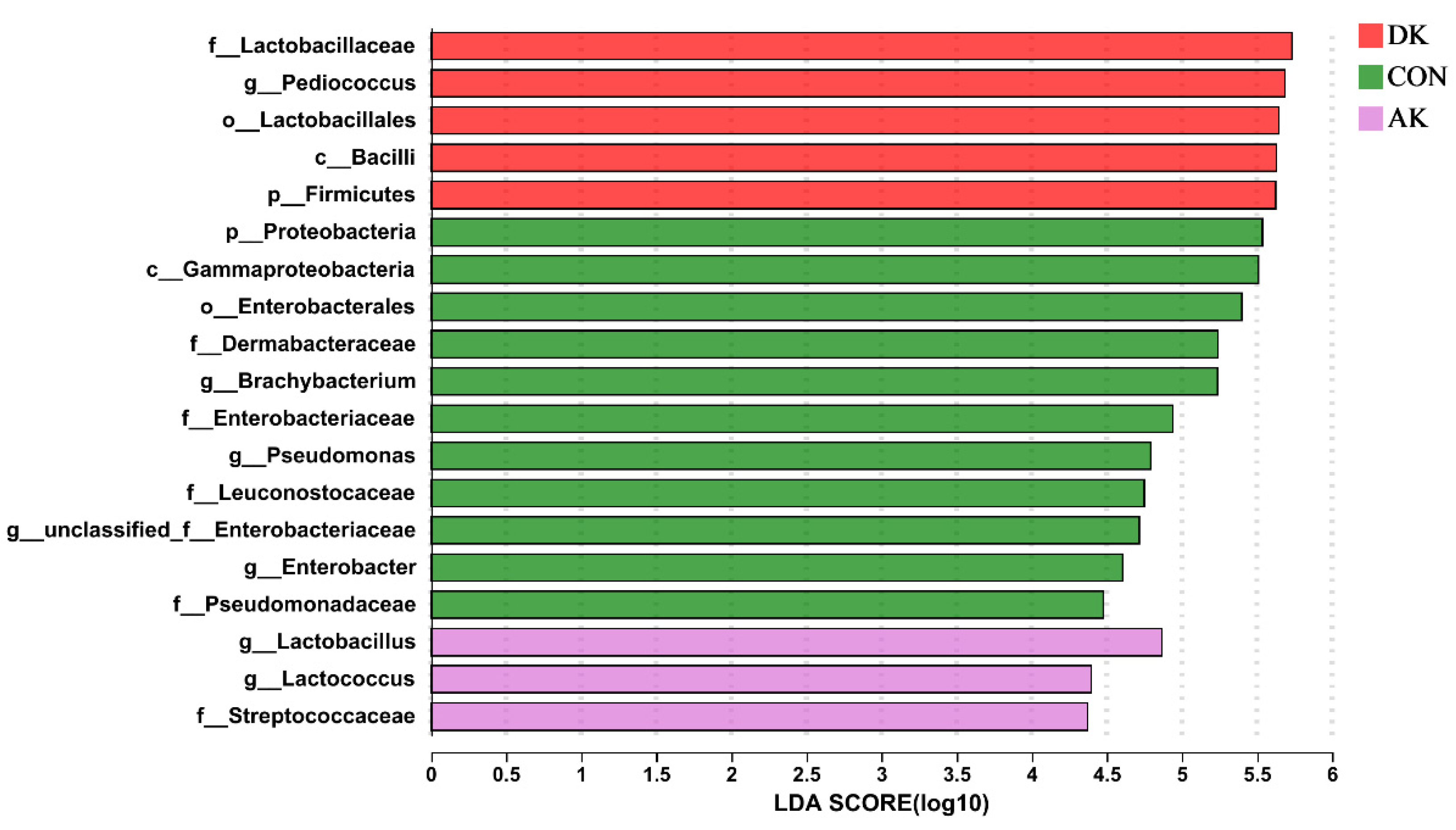
3.5. Bacterial Abundance of Fresh Mulberry Leaves and Mulberry Leaf Silage
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, Y.; Wang, J.Q.; Mei, J.; Huang, L.X.; Liu, H.Y. Effects of Mulberry Branch and Leaves Silage on Microbial Community, Rumen Fermentation Characteristics, and Milk Yield in Lactating Dairy Cows. Fermentation 2022, 8, 86. [Google Scholar] [CrossRef]
- Cheng, Q.M.; Li, M.Y.; Fan, X.Y.; Chen, Y.L.; Sun, H.; Xie, Y.X.; Zheng, Y.L.; Chen, C.; Li, P. Effects of epiphytic and exogenous lactic acid bacteria on fermentation quality and microbial community compositions of paper mulberry silage. Front. Microbiol. 2022, 13, 973500. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Mu, L.; Wang, Z.L.; Liu, J.Y.; Liu, T.L.; Wanapat, M.; Huang, B.Z. Assessment of mulberry leaf as a potential feed supplement for animal feeding in PR China. Asian-Australas. J. Anim. Sci. 2019, 32, 1145–1152. [Google Scholar] [CrossRef] [PubMed]
- Kandylis, K.; Hadjigeorgiou, I.; Harizanis, P. The nutritive value of mulberry leaves (Morus alba) as a feed supplement for sheep. Trop. Anim. Health Prod. 2009, 41, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Shen, Q.M.; Zhong, S.; Chen, Y.L.; Yang, Y.X. Comparison of Rumen Microbiota and Serum Biochemical Indices in White Cashmere Goats Fed Ensiled or Sun-Dried Mulberry Leaves. Microorganisms 2020, 8, 981. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, J.L.; Wang, M.Z.; Hou, Q.R.; Feng, D.; Pi, Y.; Zhao, W.G. Effects of Dietary Mulberry Leaf Powder in Concentrate on the Rumen Fermentation and Ruminal Epithelium in Fattening Hu Sheep. Animals 2019, 9, 218. [Google Scholar] [CrossRef]
- Al-kirshi, R.; Alimon, A.R.; Zulkifli, I.; Sazili, A.; Zahari, M.W.; Ivan, M. Utilization of mulberry leaf meal (Morus alba) as protein supplement in diets for laying hens. Ital. J. Anim. Sci. 2010, 9, 265–267. [Google Scholar] [CrossRef]
- Chen, C.; Razali, U.H.M.; Saikim, F.H.; Mahyudin, A.; Noor, N. Morus alba L. Plant: Bioactive Compounds and Potential as a Functional Food Ingredient. Foods 2021, 10, 689. [Google Scholar] [CrossRef]
- Thabti, I.; Elfalleh, W.; Tlili, N.; Ziadi, M.; Campos, M.G.; Ferchichi, A. Phenols, Flavonoids, and Antioxidant and Antibacterial Activity of Leaves and Stem Bark of Morus Species. Int. J. Food Prop. 2014, 17, 842–854. [Google Scholar] [CrossRef]
- Katsube, T.; Imawaka, N.; Kawano, Y.; Yamazaki, Y.; Shiwaku, K.; Yamane, Y. Antioxidant flavonol glycosides in mulberry (Morus alba L.) leaves isolated based on LDL antioxidant activity. Food Chem. 2006, 97, 25–31. [Google Scholar] [CrossRef]
- Park, E.; Lee, S.M.; Lee, J.E.; Kim, J.H. Anti-inflammatory activity of mulberry leaf extract through inhibition of NF-κB. J. Funct. Foods 2013, 5, 178–186. [Google Scholar] [CrossRef]
- Ma, J.Y.; Ma, H.; Liu, S.J.; Wang, J.; Wang, H.L.; Zang, J.J.; Long, S.F.; Piao, X.S. Effect of Mulberry Leaf Powder of Varying Levels on Growth Performance, Immuno-Antioxidant Status, Meat Quality and Intestinal Health in Finishing Pigs. Antioxidants 2022, 11, 2243. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.C.; Lee, M.T.; Chang, S.C.; Chang, Y.L.; Shih, C.H.; Yu, B.; Lee, T.T. Effects of mulberry leaves on production performance and the potential modulation of antioxidative status in laying hens. Poult. Sci. 2017, 96, 1191–1203. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Han, Y.; Zhao, Y.F.; Chen, D.Q.; Wang, D.F.; Yang, Y.; Su, C.Z.; Shen, X.Y. Effect of Mulberry Leaf TMR on Growth Performance, Meat Quality and Expression of Meat Quality Master Genes (ADSL, H-FABP) in Crossbred Black Goats. Foods 2022, 11, 4032. [Google Scholar] [CrossRef] [PubMed]
- Xv, Z.C.; He, G.L.; Wang, X.L.; Shun, H.; Chen, Y.J.; Lin, S.M. Mulberry leaf powder ameliorate high starch-induced hepatic oxidative stress and inflammation in fish model. Anim. Feed. Sci. Technol. 2021, 278, 115012. [Google Scholar] [CrossRef]
- Zhang, Y.C.; Li, D.X.; Wang, X.K.; Lin, Y.L.; Zhang, Q.; Chen, X.Y.; Yang, F.Y. Fermentation quality and aerobic stability of mulberry silage prepared with lactic acid bacteria and propionic acid. Anim. Sci. J. 2019, 90, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.P.; Yang, Y.X.; Zhang, M.J.; Jiao, F.; Gan, T.T.; Lin, Z.W.; Huang, Y.Z.; Wang, H.X.; Liu, S.; Bao, L.J.; et al. Optimized Ensiling Conditions and Microbial Community in Mulberry Leaves Silage With Inoculants. Front. Microbiol. 2022, 13, 813363. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yu, Z.; Wang, X.G. Isolating and evaluating lactic acid bacteria strains with or without sucrose for effectiveness of silage fermentation. Grassl. Sci. 2015, 61, 167–176. [Google Scholar] [CrossRef]
- Kung, L.M.; Shaver, R.D.; Grant, R.J.; Schmidt, R.J. Silage review: Interpretation of chemical, microbial, and organoleptic components of silages. J. Dairy Sci. 2018, 101, 4020–4033. [Google Scholar] [CrossRef]
- Trabi, E.B.; Yuan, X.J.; Li, J.F.; Dong, Z.H.; Shah, A.A.; Shao, T. Effect of Glucose and Lactic Acid Bacteria on the Fermentation Quality, Chemical Compositions and in vitro Digestibility of Mulberry (Morns alba) Leaf Silage. Pak. J. Zool. 2017, 49, 2271–2277. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, X.Y.; Wang, C.; He, L.W.; Zhou, W.; Yang, F.Y.; Zhang, Q. The bacterial community and fermentation quality of mulberry (Morus alba) leaf silage with or without Lactobacillus casei and sucrose. Bioresour. Technol. 2019, 293, 122059. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.L.; Wang, F.; Fang, Y.; Zhou, D.W.; Wang, S.P.; Wu, D.Q.; Wang, L.X.; Zhong, R.Z. High-potency white-rot fungal strains and duration of fermentation to optimize corn straw as ruminant feed. Bioresour. Technol. 2020, 312, 123512. [Google Scholar] [CrossRef] [PubMed]
- You, J.T.; Zhang, H.; Zhu, H.F.; Xue, Y.L.; Cai, Y.M.; Zhang, G.J. Microbial Community, Fermentation Quality, and in vitro Degradability of Ensiling Caragana With Lactic Acid Bacteria and Rice Bran. Front. Microbiol. 2022, 13, 804429. [Google Scholar] [CrossRef] [PubMed]
- He, L.W.; Zhou, W.; Wang, C.; Yang, F.Y.; Chen, X.Y.; Zhang, Q. Effect of cellulase and Lactobacillus casei on ensiling characteristics, chemical composition, antioxidant activity, and digestibility of mulberry leaf silage. J. Dairy Sci. 2019, 102, 9919–9931. [Google Scholar] [CrossRef] [PubMed]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- Murphy, R.P. A method for the extraction of plant samples and the determination of total soluble carbohydrates. J. Sci. Food Agric. 1958, 9, 714–717. [Google Scholar] [CrossRef]
- Broderick, G.A.; Kang, A.J.H. Automated Simultaneous Determination of Ammonia and Total Amino Acids in Ruminal Fluid and In Vitro Media. J. Dairy Sci. 1980, 63, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.Q.; Santhanam, R.K.; Xue, Z.H.; Guo, Q.W.; Gao, X.D.; Chen, H.X. Effect of different drying methods on the physicochemical properties and antioxidant activities of mulberry leaves polysaccharides. Int. J. Biol. Macromol. 2018, 119, 1137–1143. [Google Scholar] [CrossRef]
- Cuesta, G.; Suarez, N.; Bessio, M.I.; Ferreira, F.; Massaldi, H. Quantitative determination of pneumococcal capsular polysaccharide serotype 14 using a modification of phenol-sulfuric acid method. J. Microbiol. Methods 2003, 52, 69–73. [Google Scholar] [CrossRef]
- Jiang, X.; Liu, X.; Xu, H.J.; Sun, Y.K.; Zhang, Y.G.; Wang, Y. Improvement of the nutritional, antioxidant and bioavailability properties of corn gluten-wheat bran mixture fermented with lactic acid bacteria and acid protease. Lwt-Food Sci. Technol. 2021, 144, 111161. [Google Scholar] [CrossRef]
- Liao, B.Y.; Zhu, D.Y.; Thakur, K.; Li, L.; Zhang, J.G.; Wei, Z.J. Thermal and Antioxidant Properties of Polysaccharides Sequentially Extracted from Mulberry Leaves (Morus alba L.). Molecules 2017, 22, 2271. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.X.; Xie, Y.F.; Wang, W.; Yan, Y.H.; Ye, H.; Jabbar, S.; Zeng, X.X. Extraction optimization, characterization and antioxidant activity in vitro of polysaccharides from mulberry (Morus alba L.) leaves. Carbohydr. Polym. 2015, 128, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.X.; Liao, S.T.; Shen, W.Z.; Liu, F.; Tang, C.M.; Chen, C.Y.O.; Sun, Y.M. Phenolics and Antioxidant Activity of Mulberry Leaves Depend on Cultivar and Harvest Month in Southern China. Int. J. Mol. Sci. 2012, 13, 16544–16553. [Google Scholar] [CrossRef] [PubMed]
- He, L.W.; Lv, H.J.; Chen, N.; Wang, C.; Zhou, W.; Chen, X.Y.; Zhang, Q. Improving fermentation, protein preservation and antioxidant activity of Moringa oleifera leaves silage with gallic acid and tannin acid. Bioresour. Technol. 2020, 297, 122390. [Google Scholar] [CrossRef] [PubMed]
- He, L.W.; Wang, C.; Xing, Y.Q.; Zhou, W.; Pian, R.Q.; Yang, F.Y.; Chen, X.Y.; Zhang, Q. Dynamics of proteolysis, protease activity and bacterial community of Neolamarckia cadamba leaves silage and the effects of formic acid and Lactobacillus farciminis. Bioresour. Technol. 2019, 294, 122127. [Google Scholar] [CrossRef]
- Cai, Y.M.; Benno, Y.; Ogawa, M.; Ohmomo, S.; Kumai, S.; Nakase, T. Influence of Lactobacillus spp. from an inoculant and of Weissella and Leuconostoc spp. from forage crops on silage fermentation. Appl. Environ. Microbiol. 1998, 64, 2982–2987. [Google Scholar] [CrossRef] [PubMed]
- Opazo, R.; Ortúzar, F.; Navarrete, P.; Espejo, R.; Romero, J. Reduction of Soybean Meal Non-Starch Polysaccharides and α-Galactosides by Solid-State Fermentation Using Cellulolytic Bacteria Obtained from Different Environments. PLoS ONE 2012, 7, e44783. [Google Scholar] [CrossRef]
- Chi, Z.; Deng, M.; Tian, H.C.; Liu, D.W.; Li, Y.K.; Liu, G.B.; Sun, B.L.; Guo, Y.Q. Effects of Mulberry Leaves and Pennisetum Hybrid Mix-Silage on Fermentation Parameters and Bacterial Community. Fermentation 2022, 8, 197. [Google Scholar] [CrossRef]
- Jain, J.; Kumar, A.; Singh, D.; Singh, B. Purification and kinetics of a protease-resistant, neutral, and thermostable phytase from Bacillus subtilis subsp. subtilis JJBS250 ameliorating food nutrition. Prep. Biochem. Biotechnol. 2018, 48, 718–724. [Google Scholar] [CrossRef]
- He, L.W.; Chen, N.; Lv, H.J.; Wang, C.; Zhou, W.; Chen, X.Y.; Zhang, Q. Gallic acid influencing fermentation quality, nitrogen distribution and bacterial community of high-moisture mulberry leaves and stylo silage. Bioresour. Technol. 2020, 295, 122255. [Google Scholar] [CrossRef]
- Ni, K.K.; Wang, F.F.; Zhu, B.G.; Yang, J.X.; Zhou, G.A.; Pan, Y.; Tao, Y.; Zhong, J. Effects of lactic acid bacteria and molasses additives on the microbial community and fermentation quality of soybean silage. Bioresour. Technol. 2017, 238, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Polumackanycz, M.; Wesolowski, M.; Viapiana, A. Morus alba L. and Morus nigra L. Leaves as a Promising Food Source of Phenolic Compounds with Antioxidant Activity. Plant Food Hum. Nutr. 2021, 76, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Méndez-García, C.; Peláez, A.I.; Mesa, V.; Sánchez, J.; Golyshina, O.V.; Ferrer, M. Microbial diversity and metabolic networks in acid mine drainage habitats. Front. Microbiol. 2015, 6, 475. [Google Scholar]
- Liu, B.Y.; Huan, H.L.; Gu, H.R.; Xu, N.X.; Shen, Q.; Ding, C.L. Dynamics of a microbial community during ensiling and upon aerobic exposure in lactic acid bacteria inoculation-treated and untreated barley silages. Bioresour. Technol. 2019, 273, 212–219. [Google Scholar] [CrossRef]
- Wang, Y.; He, L.W.; Xing, Y.Q.; Zhou, W.; Pian, R.Q.; Yang, F.Y.; Chen, X.Y.; Zhang, Q. Bacterial diversity and fermentation quality of Moringa oleifera leaves silage prepared with lactic acid bacteria inoculants and stored at different temperatures. Bioresour. Technol. 2019, 284, 349–358. [Google Scholar] [CrossRef]
- Zhou, Y.; Drouin, P.; Lafrenière, C. Effect of temperature (5–25 °C) on epiphytic lactic acid bacteria populations and fermentation of whole-plant corn silage. J. Appl. Microbiol. 2016, 121, 657–671. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Contents |
|---|---|
| Chemical composition | |
| Dry matter (%) | 28.70 ± 0.09 |
| Crude protein (% DM) | 27.46 ± 0.14 |
| Crude fat (% DM) | 4.55 ± 0.06 |
| Neutral detergent fiber (% DM) | 36.45 ± 0.60 |
| Acid detergent fiber (% DM) | 15.69 ± 0.14 |
| Water-soluble carbohydrate (% DM) | 7.47 ± 0.13 |
| Microbial population | |
| Lactic acid bacteria (log cfu/g FM) | 4.32 ± 0.24 |
| Yeasts (log cfu/g FM) | 4.37 ± 0.04 |
| Mold (log cfu/g FM) | <2.00 |
| Coliform bacteria (log cfu/g FM) | 3.66 ± 0.06 |
| Item | Treatments | SEM | p-Value | ||
|---|---|---|---|---|---|
| A | DK | AK | |||
| Chemical composition | |||||
| Dry matter (%) | 28.50 b | 28.99 a | 29.03 a | 0.13 | 0.01 |
| Crude protein (% DM) | 27.08 | 28.00 | 28.06 | 0.51 | 0.18 |
| Crude fat (% DM) | 4.83 | 4.98 | 4.60 | 0.26 | 0.39 |
| Neutral detergent fiber (% DM) | 25.96 | 25.25 | 24.38 | 1.01 | 0.36 |
| Acid detergent fiber (% DM) | 11.75 | 11.63 | 11.89 | 0.44 | 0.84 |
| Water soluble carbohydrate (% DM) | 1.99 | 3.48 | 2.94 | 0.46 | 0.05 |
| Fermentation quality | |||||
| pH | 4.27 | 4.67 | 4.31 | 0.14 | 0.06 |
| Lactic acid (mg/g FM) | 37.25 | 26.94 | 34.47 | 3.35 | 0.05 |
| Acetic acid (mg/g FM) | 3.08 ab | 3.78 a | 2.60 b | 0.34 | 0.04 |
| Propionic acid (mg/g FM) | ND | ND | ND | - | - |
| Butyric acid (mg/g FM) | ND | ND | ND | - | - |
| Ammonia-N (%DM) | 0.02 b | 0.03 a | 0.02 b | 0.00 | <0.01 |
| Microbial population | |||||
| Lactic acid bacteria (log cfu/g FM) | 7.97 | 5.57 | 6.65 | 1.29 | 0.26 |
| Yeasts (log cfu/g FM) | <2.00 | <2.00 | <2.00 | - | - |
| Mold (log cfu/g FM) | <2.00 | <2.00 | <2.00 | - | - |
| Coliform bacteria (log cfu/g FM) | <2.00 | <2.00 | <2.00 | - | - |
| Item | Treatments | SEM | p-Value | |||
|---|---|---|---|---|---|---|
| CON | A | DK | AK | |||
| Bioactive components | ||||||
| Flavonoid (mg/g DM) | 6.72 | 7.41 | 6.06 | 6.58 | 0.54 | 0.17 |
| Polyphenol (mg/g DM) | 13.60 b | 19.07 a | 17.89 a | 13.99 b | 1.17 | <0.01 |
| Polysaccharides (mg/g DM) | 27.83 a | 13.02 b | 12.93 b | 12.00 b | 1.32 | <0.01 |
| Antioxidant activity | ||||||
| DPPH (%) | 91.18 | 92.29 | 93.51 | 95.16 | 1.25 | 0.06 |
| ABTS (%) | 95.45 | 98.53 | 97.46 | 98.03 | 1.43 | 0.23 |
| HAS (%) | 42.72 b | 75.57 a | 74.30 a | 77.36 a | 8.32 | <0.01 |
| FRAP (%) | 81.90 | 91.51 | 90.98 | 91.04 | 4.02 | 0.12 |
| Treatments | Sobs | Chao1 | Ace | Shannon | Coverage |
|---|---|---|---|---|---|
| CON | 106.33 a | 114.16 a | 112.82 | 2.35 a | 1.00 |
| A | 33.67 b | 43.83 b | 43.07 | 0.21 c | 1.00 |
| DK | 33.33 b | 43.01 b | 43.1 | 0.21 c | 1.00 |
| AK | 35.67 b | 42.75 b | 50.8 | 0.75 b | 1.00 |
| SEM | 22.62 | 22.29 | 23.65 | 0.22 | - |
| p-value | 0.03 | 0.03 | 0.05 | <0.01 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Li, G.; Zhang, H.; Yang, T.; Abbas, Z.; Jiang, X.; Zhang, H.; Zhang, R.; Si, D. The Fermentation Quality, Antioxidant Activity, and Bacterial Community of Mulberry Leaf Silage with Pediococcus, Bacillus, and Wheat Bran. Fermentation 2024, 10, 214. https://doi.org/10.3390/fermentation10040214
Li J, Li G, Zhang H, Yang T, Abbas Z, Jiang X, Zhang H, Zhang R, Si D. The Fermentation Quality, Antioxidant Activity, and Bacterial Community of Mulberry Leaf Silage with Pediococcus, Bacillus, and Wheat Bran. Fermentation. 2024; 10(4):214. https://doi.org/10.3390/fermentation10040214
Chicago/Turabian StyleLi, Jinzhuan, Guiming Li, Haosen Zhang, Tiantian Yang, Zaheer Abbas, Xiaohan Jiang, Heng Zhang, Rijun Zhang, and Dayong Si. 2024. "The Fermentation Quality, Antioxidant Activity, and Bacterial Community of Mulberry Leaf Silage with Pediococcus, Bacillus, and Wheat Bran" Fermentation 10, no. 4: 214. https://doi.org/10.3390/fermentation10040214
APA StyleLi, J., Li, G., Zhang, H., Yang, T., Abbas, Z., Jiang, X., Zhang, H., Zhang, R., & Si, D. (2024). The Fermentation Quality, Antioxidant Activity, and Bacterial Community of Mulberry Leaf Silage with Pediococcus, Bacillus, and Wheat Bran. Fermentation, 10(4), 214. https://doi.org/10.3390/fermentation10040214