The Impact of Non-Saccharomyces Yeast on Traditional Method Sparkling Wine



Abstract
:
1. Introduction
1.1. Non-Saccharomyces Yeasts
1.2. Non-Saccharomyces Yeasts and Sparkling Wine
2. Nitrogenous Compounds and Non-Saccharomyces Yeast
2.1. Yeast Acclimation and Nitrogen Requirements
2.2. Amino Acids and Ammonia
2.3. Biogenic Amines
2.4. Volatile Aroma Compounds
3. Yeast-Derived Proteins
4. Organic Acids
5. Effect of Non-Saccharomyces Yeasts on Sparkling Wine Sensory Profiles
5.1. Sensory Effects of Non-Saccharomyces Yeasts on Sparkling Wines
5.2. Sensorial Influence from Lees Aging
5.3. Glycerol
6. Conclusions and Further Research
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Jackson, R.S. Specific and Distinctive Wine Styles. In Wine Science: In Principles and Applications; Academic Press/Elsevier: Hoboken, NJ, USA, 2014; pp. 677–759. [Google Scholar]
- Vigentini, I.; Cardenas, S.B.; Valdetara, F.; Faccincani, M.; Panont, C.A.; Picozzi, C.; Foschino, R. Use of Native Yeast Strains for In-Bottle Fermentation to Face the Uniformity in Sparkling Wine Production. Front. Microbiol. 2017, 8, 1225. [Google Scholar] [CrossRef] [PubMed]
- Kemp, B.; Alexandre, H.; Robillard, B.; Marchal, R. Effect of production phase on bottle-fermented sparkling wine quality. J. Agric. Food Chem. 2015, 14, 19–38. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Magariño, S.; Ortega-Herasa, M.; Bueno-Herrera, M.; Martínez-Lapuente, L.; Guadalupe, Z.; Ayestarán, B. Grape variety, aging on lees and aging in bottle after disgorging influence on volatile composition and foamability of sparkling wines. LWT-Food Sci. Technol. 2015, 61, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Martínez-García, R.; García-Martínez, T.; Puig-Pujol, A.; Mauricio, J.C.; Moreno, J. Changes in sparkling wine aroma during the second fermentation under CO2 pressure in sealed bottle. Food Chem. 2017, 237, 1030–1040. [Google Scholar] [CrossRef] [PubMed]
- Pozo-Bayón, M.A.; Martínez-Rodríguez, M.; Pueyo, E.; Moreno-Arribas, M.V. Chemical and biochemical features involved in sparkling wine production: From a traditional to an improved winemaking technology. Trends Food Sci. Technol. 2009, 20, 289–299. [Google Scholar] [CrossRef]
- Torresi, S.; Frangipane, M.T.; Anelli, G. Biotechnologies in sparkling wine production. Interesting approaches for quality improvement: A review. Food Chem. 2011, 129, 1232–1241. [Google Scholar] [CrossRef] [PubMed]
- Riu-Aumatell, M.; Torrens, J.; Buxaderas, S.; López-Tamames, E. Cava (Spanish sparkling wine) aroma: Composition and determination methods. In Recent Advances in Pharmaceutical Sciences III; Muñoz-Torrero, D., Cortés, A., Mariño, E.L., Eds.; Transworld Research Network: Kerala, India, 2013; pp. 45–60. [Google Scholar]
- Caliari, V.; Burin, V.M.; Rosier, J.P.; BordignonLuiz, M.T. Aromatic profile of Brazilian sparkling wines produced with classical and innovative grape varieties. Food Res. Int. 2014, 62, 965–973. [Google Scholar] [CrossRef]
- Mafata, M. The Effect of Grape Temperature on The Phenolic Extraction and Sensory Perception of Méthode Cap Classique Wines. Ph.D. Thesis, Stellenbosch University, Stellenbosch, South Africa, March 2017. [Google Scholar]
- Buxaderas, S.; López-Tamames, E. Sparkling wines: Features and trends from tradition. Adv. Food Nutr. Res. 2012, 66, 1–45. [Google Scholar] [CrossRef] [PubMed]
- Duteurtre, B. Assemblage. In Le Champagne: De La Tradition à La Science; Lavoisier/Tec & Doc: Paris, France, 2006; pp. 116–125. [Google Scholar]
- Ribéreau-Gayon, P.; Dubourdieu, D.; Doneche, B.; Lonvaud, A. Other Winemaking Methods. In Handbook of Enology, the Microbiology of Wine and Vinifications, 2nd ed.; John Wiley and Sons Ltd.: Hoboken, NJ, USA, 2006; pp. 445–480. [Google Scholar]
- Duteurtre, B. Degorgement et bouchage. In Le Champagne: De La Tradition à La Science, 2nd ed.; Lavoisier/Tec & Doc: Paris, France, 2006; pp. 205–232. [Google Scholar]
- Masneuf-Pomarede, I.; Bely, M.; Marullo, P.; Albertin, W. The genetics of non-conventional wine yeasts: Current knowledge and future challenges. Front. Microbiol. 2016, 6, 1563. [Google Scholar] [CrossRef] [PubMed]
- Romani, C.; Lencioni, L.; Gobbi, M.; Mannazzu, I.; Ciani, M.; Domizio, P. Schizosaccharomyces japonicus: A polysaccharide-overproducing yeast to be used in winemaking. Fermentation 2018, 4, 14. [Google Scholar] [CrossRef]
- Domizio, P.; Romani, C.; Lencioni, L.; Comitini, F.; Gobbi, M.; Mannazzu, I.; Ciani, M. Outlining a future for non-Saccharomyces yeasts: Selection of putative spoilage wine strains to be used in association with Saccharomyces cerevisiae for grape juice fermentation. Int. J. Food Microbiol. 2011, 147, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [PubMed]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 5, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Varela, C.; Siebert, T.; Cozzolino, D.; Rose, L.; Mclean, H.; Henschke, P.A. Discovering a chemical basis for differentiating wines made by fermentation with ‘wild’ indigenous and inoculated yeasts: Role of yeast volatile compounds. Aust. J. Grape Wine Res. 2009, 15, 238–248. [Google Scholar] [CrossRef]
- García, M.; Esteve-Zarzoso, B.; Arroyo, T. Non-Saccharomyces Yeasts: Biotechnological Role for Wine Production. In Grape and Wine Biotechnology; Morata, A., Ed.; IntechOpen: London, UK, 2016; pp. 249–271. [Google Scholar] [CrossRef]
- Chasseriaud, L.; Coulon, J.; Marullo, P.; Albertin, W.; Bely, M. New oenological practice to promote non-Saccharomyces species of interest: Saturating grape juice with carbon dioxide. Appl. Microbiol. Biotechnol. 2018, 102, 3779–3791. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, B.; Bauer, F.F.; Setati, M.E. The impact of Saccharomyces cerevisiae on a wine yeast consortium in natural and inoculated fermentations. Front. Microbiol. 2017, 8, 1988. [Google Scholar] [CrossRef] [PubMed]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past and Future of Non-Saccharomyces Yeasts: From Spoilage Microorganisms to Biotechnological Tools for Improving Wine Aroma Complexity. Front. Microbiol. 2016, 7, 411. [Google Scholar] [CrossRef] [PubMed]
- Gschaedler, A. Contribution of non-conventional yeasts in alcoholic beverages. Curr. Opin. Food Sci. 2017, 13, 73–77. [Google Scholar] [CrossRef]
- Lleixà, J.; Martín, V.; Portillo, M.D.; Carrau, F.; Beltran, G.; Mas, A. Comparison of Fermentation and Wines Produced by Inoculation of Hanseniaspora vineae and Saccharomyces cerevisiae. Front. Microbiol. 2016, 7, 338. [Google Scholar] [CrossRef]
- Comitini, F.; Capece, A.; Ciani, M.; Romano, P. New insights on the use of wine yeasts. Curr. Opin. Food Sci. 2017, 13, 44–49. [Google Scholar] [CrossRef]
- Petruzzi, L.; Capozzi, V.; Berbegal, C.; Corbo, M.R.; Bevilacqua, A.; Spano, G.; Sinigaglia, M. Microbial Resources and Enological Significance: Opportunities and Benefits. Front. Microbiol. 2017, 8, 995. [Google Scholar] [CrossRef] [PubMed]
- Benito, S.; Hofmann, T.; Laier, M.; Lochbühler, B.; Schüttler, A.; Ebert, K.; Fritsch, S.; Röcker, J.; Rauhut, D. Effect on quality and composition of Riesling wines fermented by sequential inoculation with non-Saccharomyces and Saccharomyces cerevisiae. Eur. Food Res. Technol. 2015, 241, 707–717. [Google Scholar] [CrossRef]
- Varela, B.; Borneman, A.R. Yeasts found in vineyards and wineries. Yeast 2017, 34, 111–128. [Google Scholar] [CrossRef] [PubMed]
- Tristezza, M.; Tufariello, M.; Capozzi, V.; Spano, G.; Mita, G.; Grieco, F. The Oenological Potential of Hanseniaspora uvarum in Simultaneous and Sequential Co-fermentation with Saccharomyces cerevisiae for Industrial Wine Production. Front. Microbiol. 2016, 7, 670. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Jin, G.J.; Xu, Y.H.; Tao, Y.S. Wine aroma response to different participation of selected Hanseniaspora uvarum in mixed fermentation with Saccharomyces cerevisiae. Food Res. Int. 2018, 108, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Medina, K.; Boido, E.; Fariña, L.; Gioia, O.; Gomez, M.E.; Barquet, M.; Gaggero, C.; Dellacassa, E.; Carrau, F. Increased flavour diversity of Chardonnay wines by spontaneous fermentation and co-fermentation with Hanseniaspora vineae. Food Chem. 2013, 141, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Loira, I.; Vejarano, R.; Banuelos, M.A.; Morata, A.; Tesfaye, W.; Uthurry, C.; Villa, A.; Cintora, I.; Suarez-Lepe, J.A. Influence of sequential fermentation with Torulaspora delbrueckii and Saccharomyces cerevisiae on wine quality. LWT-Food Sci. Technol. 2014, 59, 915–922. [Google Scholar] [CrossRef]
- González-Royo, E.; Pascual, O.; Kontoudakis, N.; Esteruelas, M.; Esteve-Zarzoso, B.; Mas, A.; Canals, J.M.; Zamora, F. Oenological consequences of sequential inoculation with non-Saccharomyces yeasts (Torulaspora delbrueckii or Metschnikowia pulcherrima) and Saccharomyces cerevisiae in base wine for sparkling wine production. Eur. Food Res. Technol. 2015, 240, 999–1012. [Google Scholar] [CrossRef]
- Medina-Trujillo, L.; González-Royo, E.; Sieczkowski, N.; Heras, J.; Fort, F.; Canals, J.M.; Zamora, F. Effect of sequential inoculation (Torulaspora delbrueckii/Saccharomyces cerevisiae) in the first fermentation on the foam properties of sparkling wine (Cava). Eur. Food Res. Technol. 2017, 243, 681–688. [Google Scholar] [CrossRef]
- Belda, I.; Ruiz, J.; Beisert, B.; Navascués, E.; Marquina, D.; Calderon, F.; Rauhut, D.; Benito, S.; Santos, A. Influence of Torulaspora delbrueckii in varietal thiol (3-SH and 4-MSP) release in wine sequential fermentations. Int. J. Food Microbiol. 2017, 257, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Marcon, A.R.; Schwarz, L.V.; Dutra, S.V.; Moura, S.; Agostini, F.; Delamare, A.P.L.; Echeverrigaray, S. Contribution of a Brazilian Torulaspora delbrueckii isolate and a commercial Saccharomyces cerevisiae to the aroma profile and sensory characteristics of Moscato Branco wines. Aust. J. Grape Wine Res. 2018. [Google Scholar] [CrossRef]
- Varela, C.; Barker, A.; Tran, T.; Borneman, A.; Curtin, C. Sensory profile and volatile aroma composition of reduced alcohol Merlot wines fermented with Metschnikowia pulcherrima and Saccharomyces uvarum. Int. J. Food Microbiol. 2017, 252, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Escott, C.; Loira, I.; Del Fresno, J.M.; Morata, M.; Tesfaye, W.; Calderon, F.; Suarez-Lepe, J.A.; Han, S.; Benito, S. Use of non-Saccharomyces yeasts and oenological tannin in red winemaking: Influence on colour, aroma and sensorial properties of young wines. Food Microbiol. 2018, 69, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Benucci, I.; Cerreti, M.; Liburdi, K.; Nardi, T.; Vagnolic, P.; Ortiz-Julien, A.; Esti, M. Pre-fermentative cold maceration in presence of non-Saccharomyces strains: Evolution of chromatic characteristics of Sangiovese red wine elaborated by sequential inoculation. Food Res. Int. 2018, 107, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Englezos, V.; Torchio, F.; Cravero, F.; Marengo, F.; Giacosa, S.; Gerbi, V.; Rantsiou, K.; Rolle, L.; Cocolin, L. Aroma profile and composition of Barbera wines obtained by mixed fermentations of Starmerella bacillaris (synonym Candida zemplinina) and Saccharomyces cerevisiae. LWT-Food Sci. Technol. 2016, 73, 567–575. [Google Scholar] [CrossRef]
- Morata, A.; Benito, S.; Loira, I.; Palomero, F.; Gonzalez, M.C.; Suarez-Lepe, J.A. Formation of pyranoanthocyanins by Schizosaccharomyces pombe during the fermentation of red must. Int. J. Food Microbiol. 2012, 159, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Benito, A.; Jeffares, D.; Palomero, F.; Calderón, F.; Bai, F.-Y.; Bähler, J.; Benito, S. Selected Schizosaccharomyces pombe Strains Have Characteristics That Are Beneficial for Winemaking. PLoS ONE 2016, 11, e0151102. [Google Scholar] [CrossRef] [PubMed]
- Gobbi, M.; Comitini, F.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Lachancea thermotolerans and Saccharomyces cerevisiae in simultaneous and sequential co-fermentation: A strategy to enhance acidity and improve the overall quality of wine. Food Microbiol. 2013, 33, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Lencioni, L.; Romani, C.; Gobbi, M.; Comitini, F.; Ciani, M.; Domizio, P. Controlled mixed fermentation at winery scale using Zygotorulaspora florentina and Saccharomyces cerevisiae. Int. J. Food Microbiol. 2016, 3, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Albertin, W.; Zimmer, A.; Miot-Sertier, C.; Bernard, M.; Coulon, J.; Moine, V.; Colonna-Ceccaldi, B.; Bely, M.; Marullo, P.; Masneuf-Pomarede, I. Combined effect of the Saccharomyces cerevisiae lag phase and the non-Saccharomyces consortium to enhance wine fruitiness and complexity. Appl. Microbiol. Biotechnol. 2017, 101, 7603–7620. [Google Scholar] [CrossRef] [PubMed]
- Garavaglia, J.; Schneider, R.D.; Mendes, S.D.; Welke, J.E.; Zini, C.A.; Caramao, E.B.; Valente, P. Evaluation of Zygosaccharomyces bailii BCV 08 as a co-starter in wine fermentation for the improvement of ethyl esters production. Microbiol. Res. 2015, 173, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Escribano, R.; González-Arenzana, L.; Portu, J.; Garijo, P.; López-Alfaro, I.; López, R.; Santamaría, P.; Gutiérrez, A.R. Wine aromatic compound production and fermentative behaviour within different non-Saccharomyces species and clones. J. Appl. Microbiol. 2018, 124, 1521–1531. [Google Scholar] [CrossRef] [PubMed]
- Loira, I.; Morata, A.; Comuzzo, P.; Callejo, M.J.; González, C.; Calderón, F.; Suárez Lepe, J.A. Use of Schizosaccharomyces pombe and Torulaspora delbrueckii strains in mixed and sequential fermentations to improve red wine sensory quality. Food Res. Int. 2015, 76, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Canonico, L.; Comitini, F.; Ciani, M. Torulaspora delbrueckii for secondary fermentation in sparkling wine production. Food Microbiol. 2018, 74, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Ivit, N.N.; Loira, I.; Morata, A.; Benito, S.; Palomero, F.; Suarez-Lepe, J.A. Making natural sparkling wines with non-Saccharomyces yeasts. Eur. Food Res. Technol. 2018, 244, 925–935. [Google Scholar] [CrossRef]
- Whitener, M.E.B.; Stanstrup, J.; Panzeri, V.; Carlin, S.; Divol, B.; Du Toit, M.; Vrhovsek, U. Untangling the wine metabolome by combining untargeted SPME–GCxGCTOF-MS and sensory analysis to profile Sauvignon blanc co-fermented with seven different yeasts. Metabolomics 2016, 12, 53. [Google Scholar] [CrossRef]
- Balikci, E.K.; Tanguler, H.; Jolly, N.P.; Erten, H. Influence of Lachancea thermotolerans on cv. Emir wine fermentation. Yeast 2016, 33, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Rodríguez, A.J.; Carrascosa, A.V.; Martin-Alvarez, P.J.; Moreno-Arribas, V.; Polo, M.C. Influence of the yeast strain on the changes of the amino acids, peptides and proteins during sparkling wine production by the traditional method. J. Ind. Microbiol. Biotechnol. 2002, 29, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Perpetuini, G.; Di Gianvito, P.; Arfelli, G.; Schirone, M.; Corsetti, A.; Tofalo, R.; Suzzi, G. Biodiversity of autolytic ability in flocculent Saccharomyces cerevisiae strains suitable for traditional sparkling wine fermentation. Yeast 2016, 33, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Di Gianvito, P.; Perpetuini, G.; Tittarelli, F.; Schirone, M.; Arfelli, G.; Piva, A.; Patrignani, F.; Lanciotti, R.; Olivastric, L.; Suzzi, G.; et al. Impact of Saccharomyces cerevisiae strains on traditional sparkling wines production. Food Res. Int. 2018, 109, 552–560. [Google Scholar] [CrossRef] [PubMed]
- Borrull, A.; Poblet, M.; Rozes, N. New insights into the capacity of commercial wine yeasts to grow on sparkling wine media. Factor screening for improving wine yeast selection. Food Microbiol. 2015, 48, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Penacho, V.; Valero, E.; Gonzalez, R. Transcription profiling of sparkling wine second fermentation. Int. J. Food Microbiol. 2012, 153, 176–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garofalo, C.; Arena, M.P.; Laddomada, B.; Cappello, M.S.; Bleve, G.; Grieco, F.; Beneduce, L.; Berbegal, C.; Spano, G.; Capozzi, V. Starter Cultures for Sparkling Wine. Fermentation 2016, 2, 21. [Google Scholar] [CrossRef]
- Suárez Lepe, J.A.; Iñigo Leal, B. Vinificaciones especiales desde el punto de vista microbiológico. In Microbiología Enológica, Fundamentos de Vinificación, 3rd ed.; Ediciones Mundi-Prensa: Madrid, Spain, 2004; pp. 607–673. [Google Scholar]
- Jackson, R.S. Fermentation. In Wine Science: In Principles and Applications, 4th ed.; Academic Press/Elsevier: Hoboken, NJ, USA, 2014; pp. 427–534. [Google Scholar]
- Martínez-Rodríguez, A.J.; Polo, M.C. Effect of the addition of bentonite to the tirage solution on the nitrogen composition and sensory quality of sparkling wines. Food Chem. 2003, 81, 383–388. [Google Scholar] [CrossRef]
- Alexandre, H.; Guilloux-Benatier, M. Yeast autolysis in sparkling wine—A review. Aust. J. Grape Wine Res. 2006, 12, 119–127. [Google Scholar] [CrossRef]
- Moreno-Arribas, M.V.; Polo, M.C.; Pozo-Bayón, M.A. Peptides. In Wine Chemistry and Biochemistry; Moreno-Arribas, M.V., Polo, M.C., Eds.; Springer Science Business Media LLC: Berlin, Germany, 2009; pp. 191–209. [Google Scholar]
- Martí-Raga, M.; Sancho, M.; Guillamon, J.M.; Mas, A.; Beltran, G. The effect of nitrogen addition on the fermentative performance during sparkling wine production. Food Res. Int. 2015, 67, 126–135. [Google Scholar] [CrossRef]
- Gobert, A.; Tourdot-Maréchal, T.; Morge, C.; Sparrow, C.; Liu, Y.; Quintanilla-Casas, B.; Vichi, S.; Alexandre, H. Non-Saccharomyces yeasts nitrogen source preferences: Impact on sequential fermentation and wine volatile compounds profile. Front. Microbiol. 2017, 8, 2175. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Arribas, V.; Pueyo, E.; Polo, M.C.; Martin-Alvarez, P.J. Changes in the Amino Acid Composition of the Different Nitrogenous Fractions during the Aging of Wine with Yeasts. J. Agric. Food Chem. 1998, 46, 4042–4051. [Google Scholar] [CrossRef]
- Charpentier, C.; Feuillat, M. Yeast autolysis. In Wine Microbiology and Biotechnology; Fleet, G., Ed.; Harwood Academic Publishers: London, UK, 1993; pp. 225–242. [Google Scholar]
- Borrull, A.; Lopez-Martínez, G.; Miro-Abella, E.; Salvado, Z.; Poblet, M.; Cordero-Otero, R.; Rozes, N. New insights into the physiological state of Saccharomyces cerevisiae during ethanol acclimation for producing sparkling wines. Food Microbiol. 2016, 54, 20–29. [Google Scholar] [CrossRef]
- Leroy, M.J.; Charpentier, M.; Duteurtre, B.; Feuillat, M.; Charpentier, C. Yeast Autolysis during Champagne Aging. Am. J. Enol. Vitic. 1990, 41, 21–28. [Google Scholar]
- Martí-Raga, M.; Marullo, P.; Beltran, G.; Mas, A. Nitrogen modulation of yeast fitness and viability during sparkling wine production. Food Microbiol. 2016, 54, 106–114. [Google Scholar] [CrossRef]
- Feuillat, M.; Charpentier, C. Autolysis of yeasts in Champagne. Am. J. Enol. Vitic. 1982, 33, 6–13. [Google Scholar]
- Ribéreau-Gayon, P.; Glories, Y.; Maujean, A.; Dubourdieu, D. Nitrogen Compounds. In Handbook of Enology the Chemistry of Wine Stabilization and Treatments, 2nd ed.; John Wiley and Sons Ltd.: Hoboken, NJ, USA, 2006; Volume 2, pp. 109–140. [Google Scholar]
- Lambrechts, M.G.; Pretorius, I.S. Yeast and its Importance to Wine Aroma-A Review. S. Afr. J. Enol. Vitic. 2000, 21, 97–129. [Google Scholar]
- Bozdogan, A.; Canbas, A. Influence of yeast strain, immobilisation and ageing time on the changes of free amino acids and amino acids in peptides in bottle-fermented sparkling wines obtained from Vitis vinifera cv. Emir. Int. J. Food Sci. Technol. 2011, 46, 1113–1121. [Google Scholar] [CrossRef]
- Bozdogan, A.; Canbas, A. The effect of yeast strain, immobilisation, and ageing time on the amount of free amino acids and amino acids in peptides of sparkling wines obtained from cv. Dimrit grapes. S. Afr. J. Enol. Vitic. 2012, 33, 257–263. [Google Scholar] [CrossRef]
- Puig-Deu, M.; Lopez-Tamames, E.; Buxaderas, S.; Torre-Boronat, M.C. Quality of base and sparkling wines as influenced by the type of fining agent added pre-fermentation. Food Chem. 1999, 66, 35–42. [Google Scholar] [CrossRef]
- Lleixà, J.; Manzano, M.; Mas, A.; Portillo, M.C. Saccharomyces and non-Saccharomyces competition during microvinification under different sugar and nitrogen conditions. Front. Microbiol. 2016, 7, 1959. [Google Scholar] [CrossRef] [PubMed]
- Palomero, F.; Morata, A.; Benito, S.; Calderón, F.; Suárez-Lepe, J.A. New genera of yeasts for over-lees aging of red wine. Food Chem. 2009, 112, 432–441. [Google Scholar] [CrossRef]
- Marcobal, A.; Martin-Alvarez, P.J.; Polo, M.C.; Munoz, R.; Moreno-Arribas, M.V. Formation of Biogenic Amines throughout the Industrial Manufacture of Red Wine. J. Food Prot. 2006, 69, 397–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anli, R.E.; Bayram, M. Biogenic Amines in Wines. Food Rev. Int. 2008, 25, 86–102. [Google Scholar] [CrossRef]
- Ke, R.; Weic, Z.; Bogdald, C.; Göktaşe, R.K.; Xiaoa, R. Profiling wines in China for the biogenic amines: A nationwide survey and pharmacokinetic fate modelling. Food Chem. 2018, 250, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Martuscelli, M.; Arfelli, G.; Manetta, A.C.; Suzzi, G. Biogenic amines content as a measure of the quality of wines of Abruzzo (Italy). Food Chem. 2013, 140, 590–597. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, C.; Bordiga, M.; Pérez-Álvarez, E.P.; Travaglia, F.; Arlorio, M.; Salinasa, M.R.; Coïsson, J.D.; Garde-Cerdán, T. The impacts of temperature, alcoholic degree and amino acids content on biogenic amines and their precursor amino acids content in red wine. Food Res. Int. 2017, 99, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Marques, A.P.; Leitao, M.C.; San Romao, M.V. Biogenic amines in wines: Influence of oenological factor. Food Chem. 2008, 107, 853–860. [Google Scholar] [CrossRef]
- Costantini, A.; Vaudano, E.; Del Prete, V.; Danei, M.; Garcia-Moruna, E. Biogenic Amine Production by Contaminating Bacteria Found in Starter Preparations Used in Winemaking. J. Agric. Food Chem. 2009, 57, 10664–10669. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ruiz, A.; Gonzalez-Rompinelli, E.M.; Bartollome, B.; Moreno-Arribas, M.V. Potential of wine-associated lactic acid bacteria to degrade biogenic amines. Int. J. Food Microbiol. 2011, 148, 115–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granchi, L.; Romano, P.; Mangani, S.; Guerrini, S.; Vincenzini, M. Production of biogenic amines by wine microorganisms. Bull. OIV 2005, 78, 595–610. [Google Scholar]
- Caruso, M.; Fiore, C.; Contursi, M.; Salzano, G.; Paparella, A.; Romano, P. Formation of biogenic amines as criteria for the selection of wine yeasts. World J. Microbiol. Biotechnol. 2002, 18, 159–163. [Google Scholar] [CrossRef]
- Lehtonen, P. Determination of amines and amino acids in wine: A review. Am. J. Enol. Vitic. 1996, 47, 127–133. [Google Scholar]
- Sanlibaba, P.; Uymaz, B. Biogenic Amine Formation in Fermented Foods: Cheese and Wine. Eur. Int. J. Sci. Technol. 2015, 4, 81–92. [Google Scholar]
- Wantke, F.; Moritz, K.; Sesztak-Greinecker, G.; Gotz, M.; Hemmer, W. Histamine Content in Red and Sparkling Wine and Relationship with Wine Quality. J. Allergy Clin. Immunol. 2008, 121, 194. [Google Scholar] [CrossRef]
- Konakovsky, V.; Focke, M.; Hoffmann-Sommergruber, K.; Schmid, R.; Scheiner, O.; Moser, P.; Hemmer, W.; Jarisch, R. Levels of Histamine and other Biogenic Amines in Red Wines and Sparkling Wines. J. Allergy Clin. Immunol. 2011, 127, AB242. [Google Scholar] [CrossRef]
- Ancin-Azpilicueta, C.; Gonzalez-Marco, A.; Jimenez-Moreno, N. Current knowledge about the presence of amines in wine. Crit. Rev. Food Sci. Nutr. 2008, 48, 257–275. [Google Scholar] [CrossRef] [PubMed]
- Pozo-Bayón, M.A.; Monagas, M.; Bartolome, B.; Moreno-Arribas, V. Wine futures related to safety and consumer health: An integrated perspective. Crit. Rev. Food Sci. Nutr. 2012, 52, 31–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rapp, A.; Mandery, H. Wine aroma. Experentia 1986, 42, 873–884. [Google Scholar] [CrossRef]
- Azzolini, M.; Fedrizzi, B.; Tosi, E.; Finato, F.; Vagnoli, P.; Scrinzi, C.; Zapparoli, G. Effects of Torulaspora delbrueckii and Saccharomyces cerevisiae mixed cultures on fermentation and aroma of Amarone wine. Eur. Food Res. Technol. 2012, 235, 303–313. [Google Scholar] [CrossRef]
- Varela, C. The impact of non-Saccharomyces yeasts in the production of alcoholic beverages. Appl. Microbiol. Biotechnol. 2016, 100, 9861–9874. [Google Scholar] [CrossRef] [PubMed]
- Kemp, B.; Hogan, C.; Xu, S.; Dowling, L.; Inglis, D. The impact of wine style and sugar addition in liqueur d’expedition (dosage) solutions on traditional method sparkling wine composition. Beverages 2017, 3, 7. [Google Scholar] [CrossRef]
- Gallardo-Chacon, J.; Vichi, S.; López-Tamames, E.; Buxaderas, S. Changes in the Sorption of Diverse Volatiles by Saccharomyces cerevisiae Lees during Sparkling Wine Aging. J. Agric. Food Chem. 2010, 58, 12426–12430. [Google Scholar] [CrossRef] [PubMed]
- Mamede, M.E.O.; Cardello, H.M.A.B.; Pastore, G.M. Evaluation of an aroma similar to that of sparkling wine: Sensory and gas chromatography analyses of fermented grape musts. Food Chem. 2005, 89, 63–68. [Google Scholar] [CrossRef]
- Kemp, B.; Condé, B.; Jégou, S.; Howell, K.; Vasserot, Y.; Marchal, R. Chemical Compounds and Mechanisms involved in the Formation and Stabilization of Foam in Sparkling Wines. Crit. Rev. Food Sci. Nutr. 2018. [Google Scholar] [CrossRef] [PubMed]
- Waters, E.J.; Wallace, W.; Williams, P.J. Identification of heat unstable wine proteins and their resistance to peptidases. J. Agric. Food Chem. 1992, 40, 1514–1519. [Google Scholar] [CrossRef]
- Goncalves, F.; Heyraud, A.; de Pinho, M.N.; Rinaudo, M. Characterisation of white wine mannoproteins. J. Agric. Food Chem. 2002, 50, 6097–6101. [Google Scholar] [CrossRef] [PubMed]
- Fleet, G.H. Cell wall. In The Yeasts: Yeast Organelles; Rose, A.H., Harrison, J.S., Eds.; Academic Press: London, UK, 1991; Volume 4, pp. 199–277. [Google Scholar]
- Klis, F.M.; Boorsma, A.; De Grot, P.W.J. Cell wall construction in Saccharomyces cerevisiae. Yeast 2006, 23, 185–202. [Google Scholar] [CrossRef] [PubMed]
- Giovani, G.; Rosi, I.; Bertuccioli, M. Quantification and characterization of cell wall polysaccharides released by non-Saccharomyces yeast during alcoholic fermentation. Int. J. Food Microbiol. 2012, 160, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Domizio, P.; Liu, Y.; Bission, L.F.; Barile, D. Use of non-Saccharomyces wine yeasts as novel sources of mannoproteins in wine. Food Microbiol. 2014, 43, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Benito, S.; Palomero, F.; Morata, A.; Calderón, F.; Suárez-Lepe, J.A. New applications for Schizosaccharomyces pombe in the alcoholic fermentation of red wines. Int. Food J. Sci. Technol. 2012, 47, 2101–2108. [Google Scholar] [CrossRef]
- Bertuccioli, M.; Ferrari, S. Laboratory experience on the influence of yeast in mouthfeel. In Proceedings of the Les Entretiens Scientifiques De Lallemand, Montreal, QC, Canada, 25 September 1999. [Google Scholar]
- Polo, M.C.; Gonzalez de Llano, M.D.; Ramos, M. Derivatization and liquid chromatographic separation of peptides. In Food Analysis by HPLC; Nollet, M.L., Ed.; Dekker: New York, NY, USA, 1992; pp. 117–140. [Google Scholar]
- Contreras, A.; Hidalgo, C.; Schmidt, S.; Henshke, P.A.; Curtin, C.; Varela, C. The application of non-Saccharomyces yeast in fermentations with limited aeration as a strategy for the production of wine with reduced alcohol content. Int. J. Food Microbiol. 2015, 105, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Alonso-del-Rio, J.; Lairón-Peris, M.; Barrio, E.; Querol, A. Effect on the prevalence of Saccharomyces non cerevisiae species against S. cerevisiae wine strain on wine fermentation: Competition, physiological fitness, and influence in final wine composition. Front. Microbiol. 2017, 8, 150. [Google Scholar] [CrossRef]
- Pérez-Torrado, R.; Barrio, E.; Querol, A. Alternative yeasts for winemaking: Saccharomyces non-cerevisiae and its hybrids. Crit. Rev. Food Sci. Nutr. 2018, 58, 1780–1790. [Google Scholar] [CrossRef] [PubMed]
- Morata, A.; Loira, I.; Tesfaye, W.; Bañuelos, M.A.; González, C.; Suárez-Lepé, J.A. Lachancea thermotolerans applications in wine technology. Fermentation 2018, 4, 53. [Google Scholar] [CrossRef]
- Kapsopoulou, K.; Kapaklis, A.; Spyropoulos, H. Growth and fermentation characteristics of a strain of the wine yeast Kluyveromyces thermotolerans isolated in Greece. World J. Microbiol. Biotechnol. 2005, 21, 1599–1602. [Google Scholar] [CrossRef]
- Balmaseda, A.; Bordona, A.; Reguant, C.; Bauttista-Gallego, J. Non–Saccharomyces in wine: Effect upon Oenococcus oeni and malolactic fermentation. Front. Microbiol. 2018, 9, 534. [Google Scholar] [CrossRef] [PubMed]
- Belda, I.; Navascués, E.; Marquina, D.; Santos, A.; Calderon, F.; Benito, S. Dynamic analysis of physiological properties of Torulaspora delbrueckii in wine fermentations and its incidence on wine quality. Appl. Microbiol. Biotechnol. 2015, 99, 1911–1922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puertas, B.; Jiménez, M.J.; Cantos-Villar, E.; Cantoral, J.M.; Rodriguez, M.E. Use of Torulaspora delbuckii and Saccharomyces cerevisiae in semi-industrial inoculation to improve quality of Palomino and Chardonnay wines in warm countries. J. Appl. Microbiol. 2017, 122, 733–746. [Google Scholar] [CrossRef] [PubMed]
- Benito, S. The impact of Torulaspora delbrueckii yeast in winemaking. Appl. Microbiol. Biotechnol. 2018, 102, 3081–3094. [Google Scholar] [CrossRef] [PubMed]
- Maicas, S.; Ferrer, S.; Pardo, I. NAD(P)H regeneration is the key for heterolactic fermentation of hexoses in Oenococcus oeni. Microbiology 2002, 148, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Benito, S.; Palomero, P.; Calderón, F.; Palmero, D.; Suárez-Lepé, J.A. Selection of appropriate Schizosaccharomyces strains for winemaking. Food Microbiol. 2014, 42, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.K.; Lan, Y.B.; Zhu, B.Q.; Xiang, X.F.; Duana, C.Q.; Shia, Y. Changes in monosaccharides, organic acids and amino acids during Cabernet Sauvignon wine ageing based on a simultaneous analysis using gas chromatography–mass spectrometry. J. Food Sci. Agric. 2017, 98, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Vannier, A.; Brun, O.X.; Feinberg, M.H. Application of sensory analysis to champagne wine characterisation and discrimination. Food Qual. Prefer. 1999, 10, 101–107. [Google Scholar] [CrossRef]
- Buxaderas, S.; López-Tamames, E. Managing the quality of sparkling wine. In Managing Wine Quality, Viticulture and Wine Quality; Reynolds, A., Ed.; Woodhead Food Series: Sawston, UK, 2010; Volume 2, pp. 553–588. [Google Scholar]
- Martínez-Rodríguez, A.; Carrascosa, A.V.; Barcenilla, J.M.; Pozo-Bayón, M.A.; Polo, M.C. Autolytic capacity and foam analysis as additional criteria for the selection of yeast strains for sparkling wine production. Food Microbiol. 2001, 18, 183–191. [Google Scholar] [CrossRef]
- Hidalgo, P.; Pueyo, E.; Pozo-Bayón, M.A.; Martínez-Rodríguez, A.J.; Martinez-Alvarez, P.; Polo, M.C. Sensory and Analytical Study of Rosé Sparkling Wines Manufactured by Second Fermentation in the Bottle. J. Agric. Food Chem. 2004, 52, 6640–6645. [Google Scholar] [CrossRef] [PubMed]
- McMahon, K.M.; Diako, C.; Aplin, J.; Mattinson, D.S.; Culver, C.; Rossa, C.F. Trained and consumer panel evaluation of sparkling wines sweetened to brut or demi sec residual sugar levels with three different sugars. Food Res. Int. 2017, 99, 173–185. [Google Scholar] [CrossRef] [PubMed]
- López de Lerma, N.; Peinado, R.A.; Puig-Pujol, A.; Mauricioc, J.C.; Moreno, J.; García-Martínez, T. Influence of two yeast strains in free, bioimmobilized or immobilized with alginate forms on the aromatic profile of long aged sparkling wines. Food Chem. 2018, 250, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Liger-Belair, G.; Polidoric, G.; Zéninaria, V. Unraveling the evolving nature of gaseous and dissolved carbon dioxide in champagne wines: A state-of-the-art review, from the bottle to the tasting glass. Anal. Chim. Acta 2012, 732, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Obiols, J.M.; De la Presa-Owens, C.; Buxaderas, S.; Bori, J.L.; De la Torre-Boronat, C. Protocolo de evaluación de la formación de la efervescencia y espuma en un vino espumoso. ACE Revista de Enología 1998, 15, 3–15. [Google Scholar]
- Gallart, M.; Tomás, X.; Suberiola, G.; López-Tamames, E.; Buxaderas, S. Relationship between foam parameters obtained by the gas-sparging method and sensory evaluation of sparkling wines. J. Sci. Food Agric. 2004, 84, 127–133. [Google Scholar] [CrossRef]
- Torrens, J.; Riu-Aumatell, M.; Vichi, S.; López-Tamames, E.; Buxaderas, S. Assessment of Volatile and Sensory Profiles between Base and Sparkling Wines. J. Agric. Food Chem. 2010, 58, 2455–2461. [Google Scholar] [CrossRef] [PubMed]
- Duteurtre, B. L’organisation champenoise. In Le Champagne: De la Tradition à la Science, 2nd ed.; Lavoisier/Tec & Doc: Paris, France, 2006; pp. 19–32. [Google Scholar]
- Jolly, N.P.; Augustyn, O.P.H.; Pretorius, I.S. The Role and Use of Non-Saccharomyces Yeasts in Wine Production. S. Afr. J. Enol. Vitic. 2006, 27, 15–39. [Google Scholar] [CrossRef]
- Arroyo-Lopez, F.N.; Perez-Torrado, R.; Querol, A.; Barrio, E. Modulation of the glycerol and ethanol syntheses in the yeast Saccharomyces kudriavzevii differs from that exhibited by Saccharomyces cerevisiae and their hybrid. Food Microbiol. 2010, 27, 628–637. [Google Scholar] [CrossRef] [PubMed]
- Soden, A.; Francis, I.L.; Oakey, H.; Henschke, P.A. Effects of co-fermentation with Candida stellata and Saccharomyces cerevisiae on the aroma and composition of Chardonnay wine. Aust. J. Grape Wine Res. 2000, 6, 21–30. [Google Scholar] [CrossRef]
- Domizio, P.; Liu, Y.; Bisson, L.F.; Barile, D. Cell wall polysaccharides released during the alcoholic fermentation by Schizosaccharomyces pombe and S. japonicus: Quantification and characterization. Food Microbiol. 2017, 61, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Bely, M.; Stoeckle, P.; Masneuf-Pomarede, I.; Dubourdieu, D. Impact of mixed Torulaspora delbrueckii Saccharomyces cerevisiae culture on high-sugar fermentation. Int. J. Food Microbiol. 2008, 122, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Renault, P.; Miot-Sertier, C.; Marullo, P.; Hernandez-Orte, P.; Lagarrigue, L.; Lonvaud- Funel, A.; Bely, M. Genetic characterization and phenotypic variability in Torulaspora delbrueckii species: Potential applications in the wine industry. Int. J. Food Microbiol. 2009, 134, 201–210. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
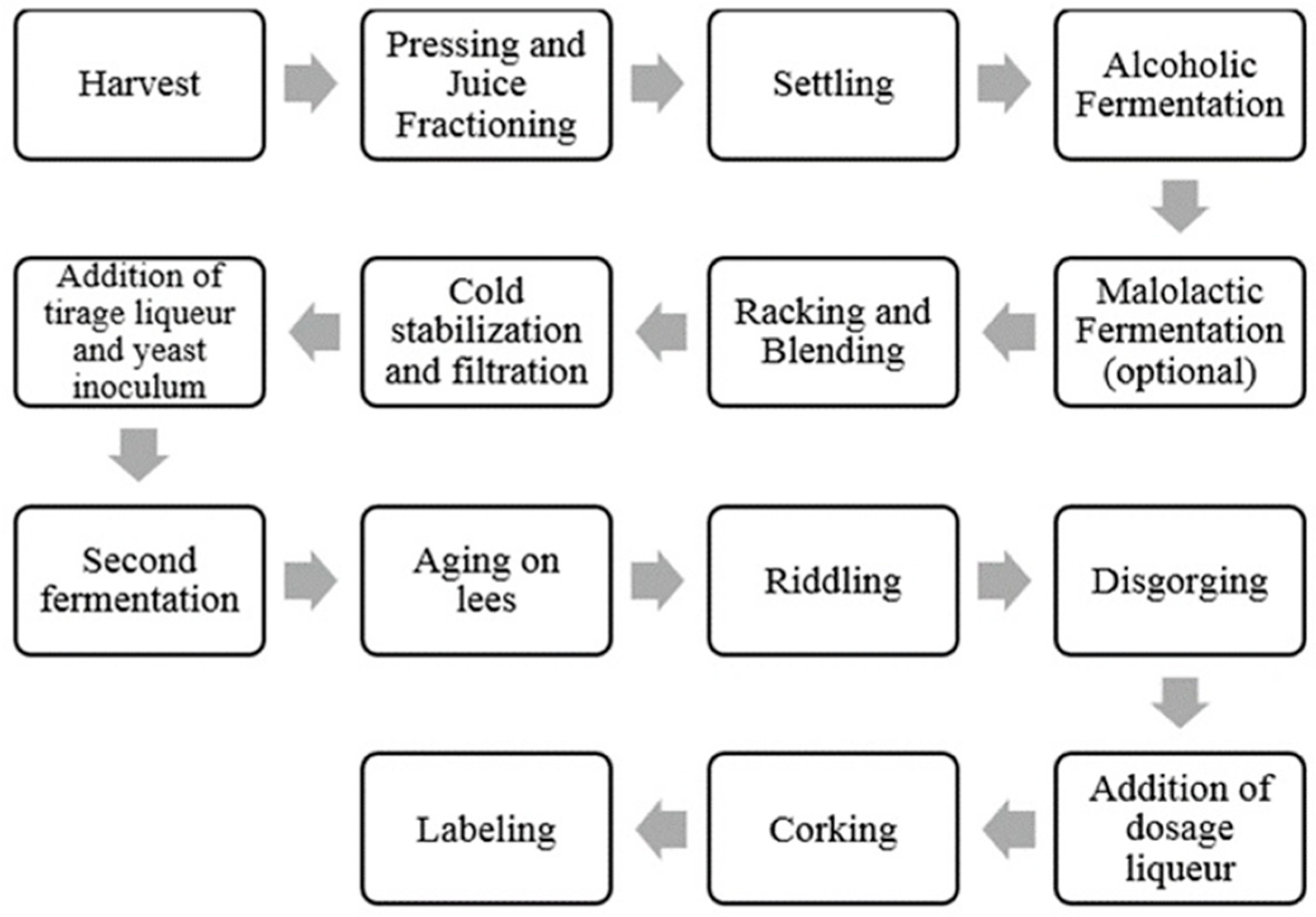{kind=link}
| Yeast Name | Production Stage | Isolation Method | Grape Variety Used | Studied Parameters | Effect on Sparkling Wine | Reference |
|---|---|---|---|---|---|---|
| T. delbrueckii | Second fermentation | Originally isolated from natural matrices from different environments. | White base wine made of Verdicchio from Ancona, Italy. | Ethanol, volatile acidity, sensory analysis and VOCs. | Positive, distinctive effects on overall aroma and sensory characteristics of wines were reported. | [51] |
| S. pombe | Second fermentation | Previously isolated yeasts from the archive of Chemistry and Food Technology Department of Universidad Politecnica de Madrid. | White base wine made of Airen grapes and red base wine made of Tempranillo grapes, Spain. | Alcohol, total acidity, pH, sugars, organic acids, glycerol, anthocyanins, VOCs, amino acids, biogenic amines, sensory analysis. | Changes on color, acidity, volatile compounds, biogenic amines of the final products as well as on the sensorial evaluation was observed. | [52] |
| S. ludwigii | ||||||
| T. delbrueckii (sequential inoculation with S. cerevisiae) | First fermentation followed by a second fermentation | Base wine from the study done by González-Royo et al. [35] fermented with S. cerevisiae bayanus (EC1118 Lallemand Inc., Montreal, QC, Canada) | V. vinifera cv. Macabeo. | Proteins, polysaccharides and foaming properties. | Better foaming properties were observed. | [36] |
| T. delbrueckii (sequential inoculation with S. cerevisiae) | First fermentation | Commercial yeasts of T. delbrueckii (BiodivaTM) and M. pulcherrima (Flavia®) from Lallemand Inc., Montreal, QC, Canada. | V. vinifera cv. Macabeo. | Ethanol, titratable acidity, pH, volatile acidity, glycerol, proteins, foaming properties, polysaccharides, VOCs, sensory analysis. | Base wines with different characteristics were obtained. Positive effects on foaming properties were observed. | [35] |
| M. pulcherrima(sequential inoculation of S. cerevisiae) |
| Yeast | Production Stage | Higher Production in Comparison to S. cerevisiae | Lower Production in Comparison to S. cerevisiae | Reference |
|---|---|---|---|---|
| T. delbrueckii + S. cerevisiae | First fermentation for base wine production | Total higher alcohols Total lactones 3-methyl-1-butanol 2-methylpropanol 1-butanol 2-phenyl ethyl acetate | β-phenylethanol 1-hexanol Ethyl lactate Ethyl decanoate Ethyl octanoate | [35] |
| M. pulcherrima + S. cerevisiae | Total higher alcohols Total major esters Total minor esters Total lactones 3-methyl-1-butanol 2-methylpropanol Diethyl succinate 2-phenyl ethyl acetate Ethyl isovalerate Methyl vanillate | Methionol Acetaldehyde Ethyl lactate Ethyl decanoate Ethyl acetate Ethyl octanoate Butyl acetate Linalool acetate | ||
| S. ludwigii 979 | Second fermentation in bottle + 4 months of aging on lees | Diacetyl Acetoin 2-methyl-1-butanol Ethyl acetate | Acetaldehyde 2.3-butan-ediol Isoamyl acetate | [52] |
| S. pombe 938 | Acetoin | Isoamyl acetate 2.3-butan-ediol | ||
| T. delbrueckii 130 | Second fermentation in bottle + 12 months of aging on lees | Ethyl hexanoate Ethyl octanoate Isoamyl acetate Hexanol | Acetaldehyde n-propanol Isobutanol Isoamyl alcohol | [51] |
| T. delbrueckii 313 | Ethyl hexanoate Ethyl octanoate Isoamyl acetate Hexanol | Acetaldehyde Ethyl butyrate n-propanol Isobutanol Isoamyl alcohol | ||
| S. cerevisiae + T. delbrueckii 130 | Ethyl hexanoate Ethyl octanoate | Acetaldehyde Ethyl butyrate Ethyl acetate n-propanol Isobutanol Isoamyl alcohol | ||
| S. cerevisiae + T. delbrueckii 313 | Ethyl hexanoate Ethyl octanoate Isoamyl acetate Hexanol | Acetaldehyde Ethyl butyrate Ethyl acetate n-propanol Isobutanol Isoamyl alcohol |
| Yeast | Production Stage | Sensory Evaluation | Effect on the Sensory Profile | Reference |
|---|---|---|---|---|
| T. delbrueckii + S. cerevisiae | First fermentation for base wine production | Sensory triangle test, panel with 9 tasters | It was distinguishable by 6 of the 9 tasters and 5 of them preferred them over control wine. | [35] |
| M. pulcherrima + S. cerevisiae | It was distinguishable by 8 of the 9 tasters and 4 of them preferred them over control wine. Smoky and flowery aromas. | |||
| T. delbrueckii (sequential inoculation with S. cerevisiae) | First fermentation followed by a second fermentation | Sensory triangle test, panel with 12 tasters | It was distinguishable by 9 of the 12 tasters and 8 of them preferred them over control wine. Better integrated effervescence and less aggressiveness in the mouth. | [36] |
| S. ludwigii 979 | Second fermentation in bottle + 4 months of aging on lees | Prepared evaluation sheet, panel with 11 tasters | In the red sparkling wines, higher limpidity and effervescence, in white sparkling wines higher limpidity but lower aroma intensity and quality in comparison to control. | [52] |
| S. pombe 7VA | In red sparkling wines, higher aroma intensity and higher scores for herbal, buttery, yeasty, acetic acid and oxidation aromas, in white sparkling wines higher limpidity; lower aroma quality, higher buttery, yeasty and reduction; lower flowery and fruity aromas in comparison to control. | |||
| T. delbrueckii 130 | Second fermentation in bottle + 12 months of aging on lees | Prepared evaluation sheet, panel with 11 tasters | It was characterized for the sensorial attributes of white flowers, bread crust, sapidity and acidity, with significant differences from other sparkling wines, except the attribute of sapidity. | [51] |
| T. delbrueckii 313 | Significant differences were detected in the main sensory attributes in comparison to control wine. Higher scores for the aromatic descriptors (white flowers, citrus, honey, odor intensity, softness). Control wine showed significantly higher astringency in comparison to all other studied fermentations. | |||
| S. cerevisiae + T. delbrueckii 130 | ||||
| S. cerevisiae + T. delbrueckii 313 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ivit, N.N.; Kemp, B. The Impact of Non-Saccharomyces Yeast on Traditional Method Sparkling Wine. Fermentation 2018, 4, 73. https://doi.org/10.3390/fermentation4030073
Ivit NN, Kemp B. The Impact of Non-Saccharomyces Yeast on Traditional Method Sparkling Wine. Fermentation. 2018; 4(3):73. https://doi.org/10.3390/fermentation4030073
Chicago/Turabian StyleIvit, Nedret Neslihan, and Belinda Kemp. 2018. "The Impact of Non-Saccharomyces Yeast on Traditional Method Sparkling Wine" Fermentation 4, no. 3: 73. https://doi.org/10.3390/fermentation4030073
APA StyleIvit, N. N., & Kemp, B. (2018). The Impact of Non-Saccharomyces Yeast on Traditional Method Sparkling Wine. Fermentation, 4(3), 73. https://doi.org/10.3390/fermentation4030073