Preventing Overflow Metabolism in Crabtree-Positive Microorganisms through On-Line Monitoring and Control of Fed-Batch Fermentations


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Strains and Culture Conditions
2.2. On-Line Process Monitoring
2.3. Specific Growth Rate Estimator
2.4. Control Strategy
2.5. Implementation and Methodology
3. Results and Discussion
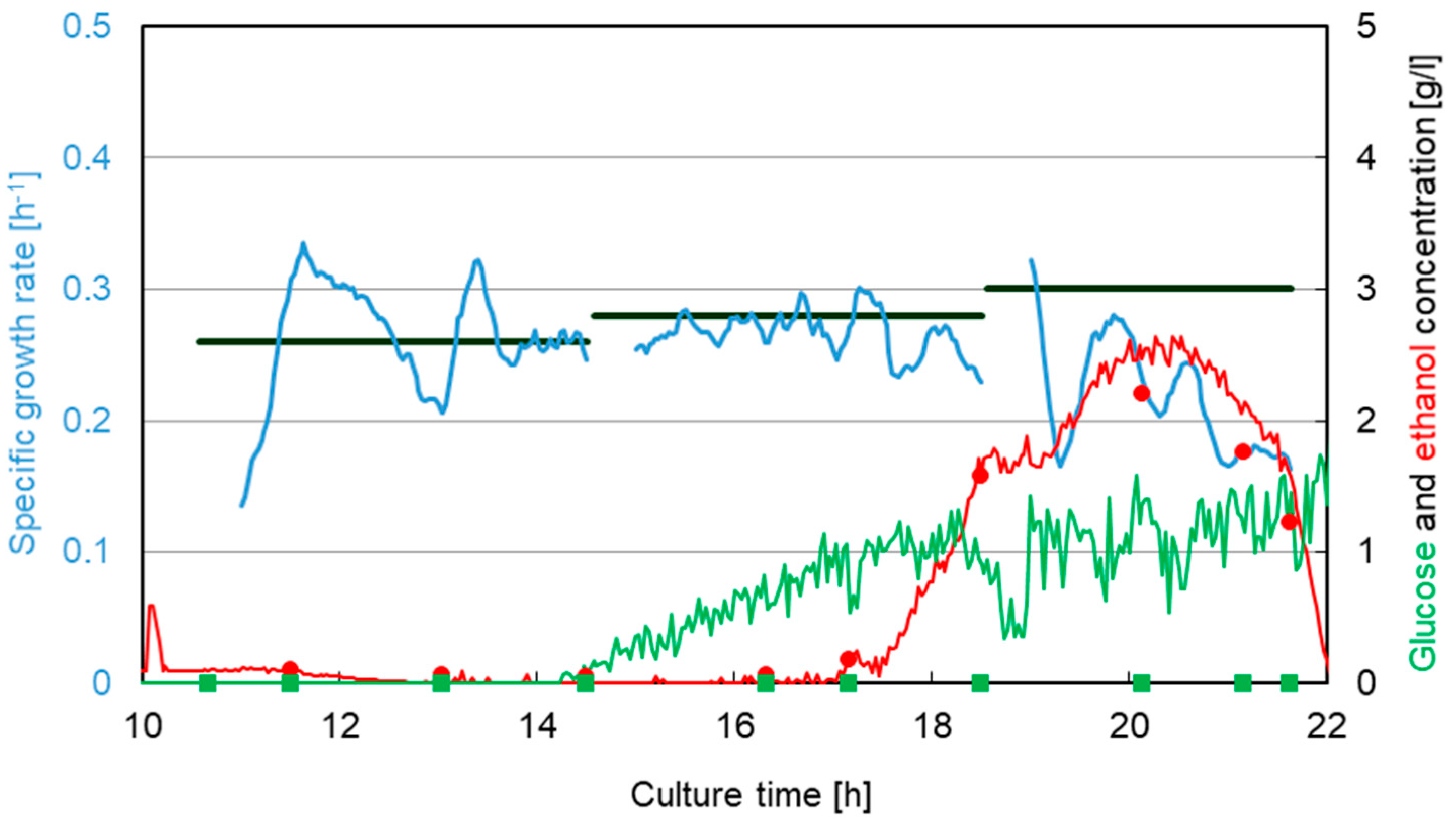
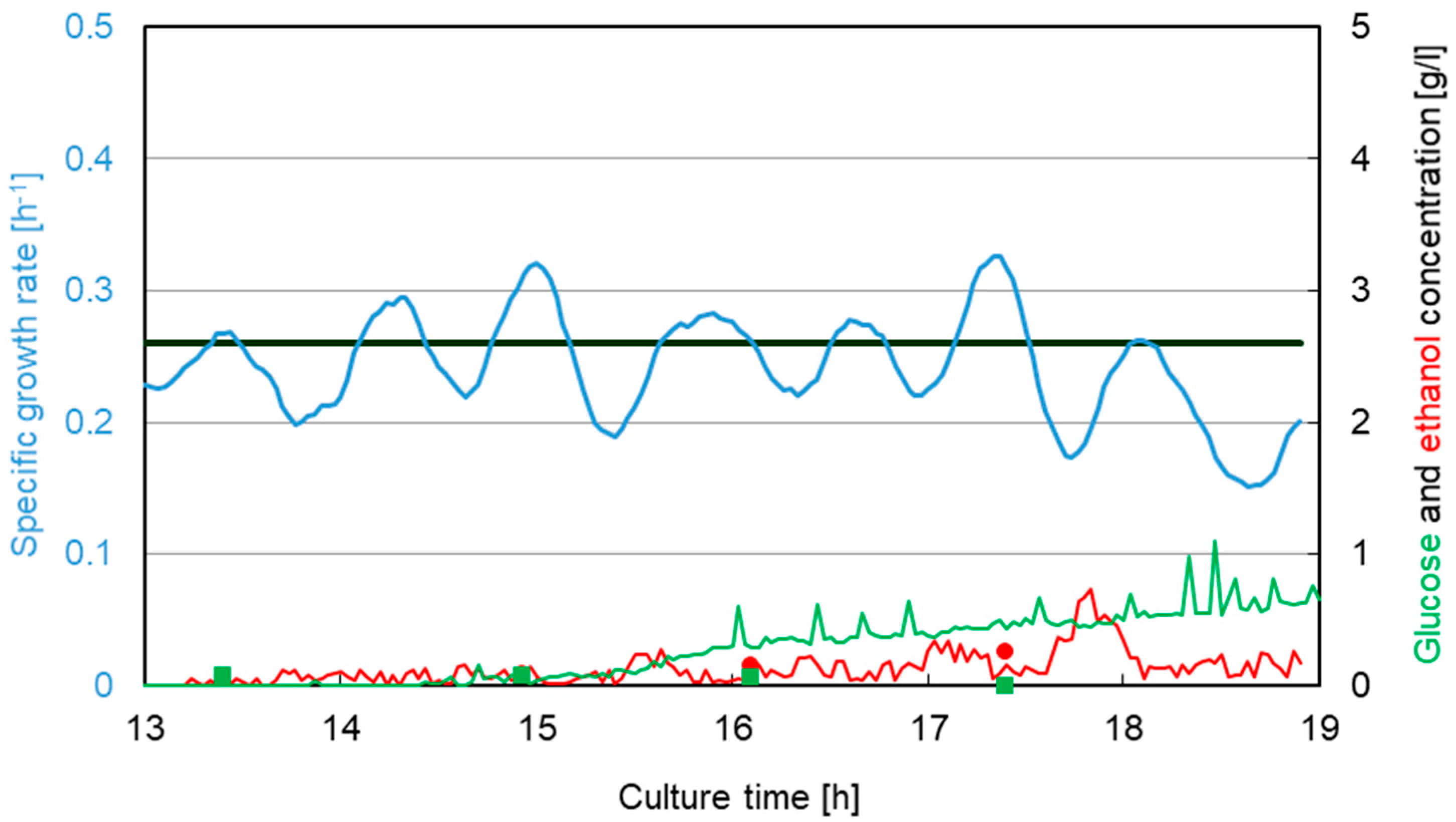
3.1. Saccharomyces cerevisiae
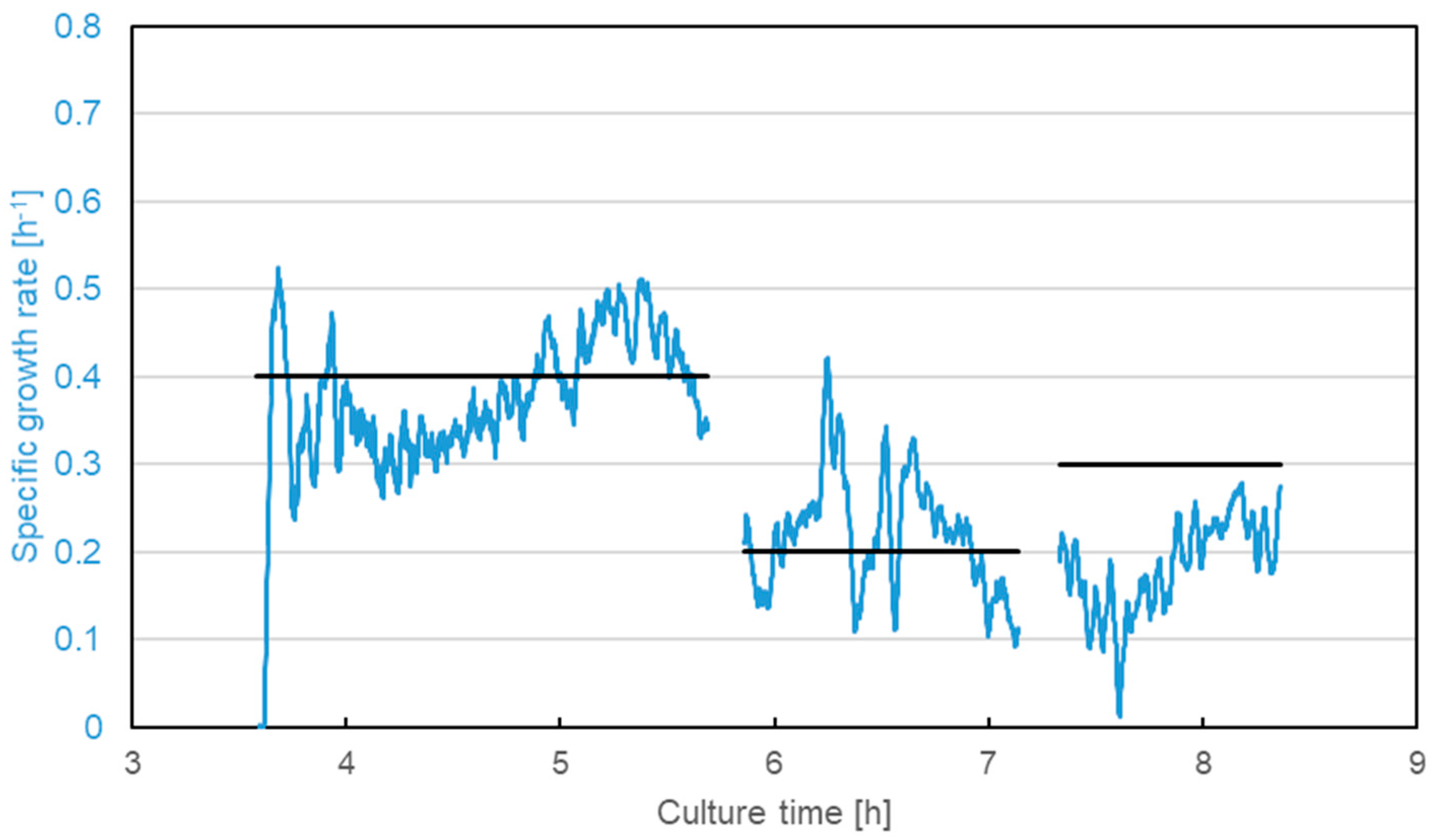
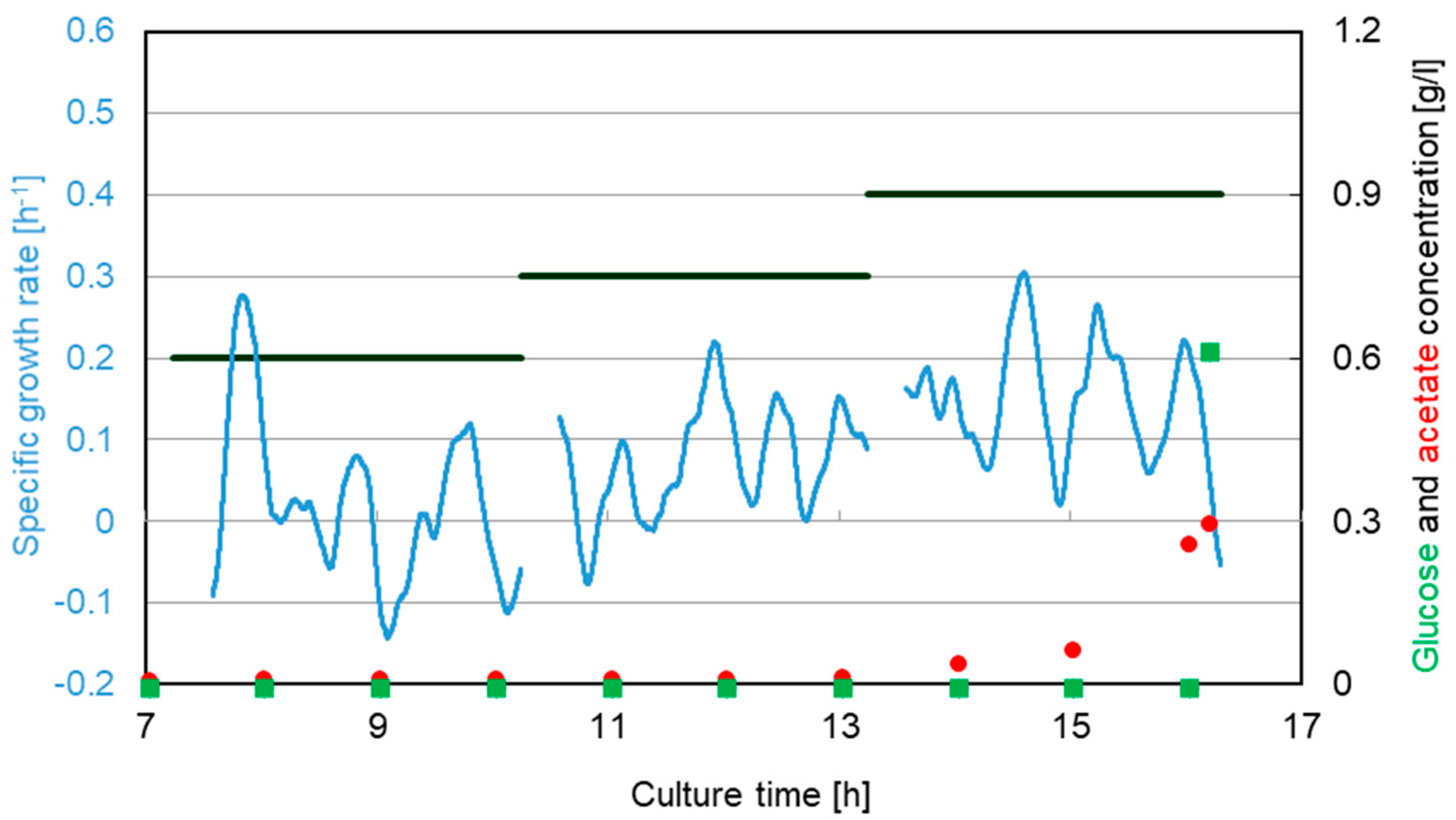
3.2. Escherichia coli
3.3. Effect on Biomass Yield
4. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mattanovich, D.; Jungo, C.; Wenger, J.; Dabros, M.; Maurer, M. Yeast suspension culture. In Industrial Scale Suspension Culture of Living Cells; Wiley: Hoboken, NJ, USA, 2014. [Google Scholar]
- Schuler, M.M.; Marison, I.W. Real-time monitoring and control of microbial bioprocesses with focus on the specific growth rate: Current state and perspectives. Appl. Microbiol. Biotechnol. 2012, 94, 1469–1482. [Google Scholar] [CrossRef] [PubMed]
- Rocha, I.; Veloso, A.C.A.; Carneiro, S.; Costa, R.; Ferreira, E.C. Implementation of a specific rate controller in a fed-batch E. coli fermentation. IFAC Proc. Vol. 2008, 41, 15565–15570. [Google Scholar] [CrossRef]
- Biener, R.; Steinkamper, A.; Horn, T. Calorimetric control of the specific growth rate during fed-batch cultures of saccharomyces cerevisiae. J. Biotechnol. 2012, 160, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Soons, Z.; van Straten, G.; van der Pol, L.A.; van Boxtel, A.J.B. Online automatic tuning and control for fed-batch cultivation. Bioprocess. Biosyst. Eng. 2008, 31, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Chopda, V.R.; Rathore, A.S.; Gomes, J. Maximizing biomass concentration in baker’s yeast process by using a decoupled geometric controller for substrate and dissolved oxygen. Bioresour. Technol. 2015, 196, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Tippmann, S.; Scalcinati, G.; Siewers, V.; Nielsen, J. Production of farnesene and santalene by saccharomyces cerevisiae using fed-batch cultivations with rq-controlled feed. Biotechnol. Bioeng. 2016, 113, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Cannizzaro, C.; Valentinotti, S.; von Stockar, U. Control of yeast fed-batch process through regulation of extracellular ethanol concentration. Bioprocess. Biosyst. Eng. 2004, 26, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Hocalar, A.; Turker, M. Model based control of minimal overflow metabolite in technical scale fed-batch yeast fermentation. Biochem. Eng. J. 2010, 51, 64–71. [Google Scholar] [CrossRef]
- Dabros, M.; Schuler, M.M.; Marison, I.W. Simple control of specific growth rate in biotechnological fed-batch processes based on enhanced online measurements of biomass. Bioprocess. Biosyst. Eng. 2010, 33, 1109–1118. [Google Scholar] [CrossRef] [PubMed]
- Mears, L.; Stocks, S.M.; Sin, G.; Gernaey, K.V. A review of control strategies for manipulating the feed rate in fed-batch fermentation processes. J. Biotechnol. 2017, 245, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Jenzsch, M.; Simutis, R.; Luebbert, A. Generic model control of the specific growth rate in recombinant escherichia coli cultivations. J. Biotechnol. 2006, 122, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Schenk, J.; Balazs, K.; Jungo, C.; Urfer, J.; Wegmann, C.; Zocchi, A.; Marison, I.W.; von Stockar, U. Influence of specific growth rate on specific productivity and glycosylation of a recombinant avidin produced by a pichia pastoris mut(+) strain. Biotechnol. Bioeng. 2008, 99, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Sivaprakasam, S.; Schuler, M.M.; Hama, A.; Hughes, K.M.; Marison, I.W. Biocalorimetry as a process analytical technology process analyser; robust in-line monitoring and control of aerobic fed-batch cultures of crabtree-negative yeast cells. J. Therm. Anal. Calorim. 2011, 104, 75–85. [Google Scholar] [CrossRef]
- Schuler, M.M.; Sivaprakasam, S.; Freeland, B.; Hama, A.; Hughes, K.-M.; Marison, I.W. Investigation of the potential of biocalorimetry as a process analytical technology (pat) tool for monitoring and control of crabtree-negative yeast cultures. Appl. Microbiol. Biotechnol. 2012, 93, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Horta, A.C.L.; da Silva, A.J.; Sargo, C.R.; Cavalcanti-Montano, I.D.; Galeano-Suarez, C.A.; Velez, A.M.; Santos, M.P.; Goncalves, V.M.; Giordano, R.C.; Zangirolami, T.C. On-line monitoring of biomass concentration based on a capacitance sensor: Assessing the methodology for different bacteria and yeast high cell density fed-batch cultures. Braz. J. Chem. Eng. 2015, 32, 821–829. [Google Scholar] [CrossRef]
- Munoz, J.A.; Brereton, R.G. Partial factorial designs for multivariate calibration: Extension to seven levels and comparison of strategy. Chemom. Intell. Lab. Syst. 1998, 43, 89–105. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. cerevisiae | E. coli (B strain) | E. coli (K12 strain) | ||||
|---|---|---|---|---|---|---|
| Batch (g/L) | Feed (g/L) | Batch (g/L) | Feed (g/L) | Batch (g/L) | Feed (g/L) | |
| Glucose | 4 | 300 | 1 | 506 | 1 | 50 |
| (NH4)2SO4 | 5 | 50 | 1.1 | 50 | 1 | 5 |
| KH2PO4 | 3 | 25 | 1 | 25 | 1 | 2.5 |
| MgSO4 x 7H2O | 0.5 | 3 | 1 | 5 | 1 | 0.5 |
| Yeast Extract | - | - | 1 | 13.5 | 1 | 1.2 |
| Peptone | - | - | 1 | 12.6 | 1 | 1.2 |
| S. cerevisiae | E. coli (B strain) | E. coli (K12 strain) | |||
|---|---|---|---|---|---|
| μSP (h−1) | YX/S (g g−1) | μSP (h−1) | YX/S (g g−1) | μSP (h−1) | YX/S (g g−1) |
| 0.26 | 0.43 | 0.20 | 0.21 | 0.20 | 0.30 |
| 0.28 | 0.42 | 0.30 | 0.22 | 0.30 | 0.44 |
| 0.30 | 0.41 | 0.40 | 0.17 | 0.40 | 0.29 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Habegger, L.; Rodrigues Crespo, K.; Dabros, M. Preventing Overflow Metabolism in Crabtree-Positive Microorganisms through On-Line Monitoring and Control of Fed-Batch Fermentations. Fermentation 2018, 4, 79. https://doi.org/10.3390/fermentation4030079
Habegger L, Rodrigues Crespo K, Dabros M. Preventing Overflow Metabolism in Crabtree-Positive Microorganisms through On-Line Monitoring and Control of Fed-Batch Fermentations. Fermentation. 2018; 4(3):79. https://doi.org/10.3390/fermentation4030079
Chicago/Turabian StyleHabegger, Loïc, Kelly Rodrigues Crespo, and Michal Dabros. 2018. "Preventing Overflow Metabolism in Crabtree-Positive Microorganisms through On-Line Monitoring and Control of Fed-Batch Fermentations" Fermentation 4, no. 3: 79. https://doi.org/10.3390/fermentation4030079
APA StyleHabegger, L., Rodrigues Crespo, K., & Dabros, M. (2018). Preventing Overflow Metabolism in Crabtree-Positive Microorganisms through On-Line Monitoring and Control of Fed-Batch Fermentations. Fermentation, 4(3), 79. https://doi.org/10.3390/fermentation4030079