Spent Yeast from Brewing Processes: A Biodiverse Starting Material for Yeast Extract Production

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Propagation and Fermentation
2.2. Yeast Pre-Treatment
2.3. Yeast Quality Control
2.4. Yeast Cell Disruption Methods
2.4.1. Ultrasonic Sonotrode
2.4.2. Standard Autolysis
2.4.3. Autolysis with Contamination of Lactobacillus brevis
2.4.4. Autolysis to Improve γ-aminobutyric Acid (GABA) Production
2.5. Production of Yeast Extract
2.6. Analysis
2.6.1. Protein and Amino Acids
2.6.2. Fat
2.6.3. Water and Ash Content
2.6.4. Folate
2.6.5. Antioxidative and Reduction Potential
2.6.6. Wort Density
2.6.7. Calculations of Apparent Attenuation
2.7. Statistical Evaluation
3. Results and Discussion
3.1. General Nutrient Composition
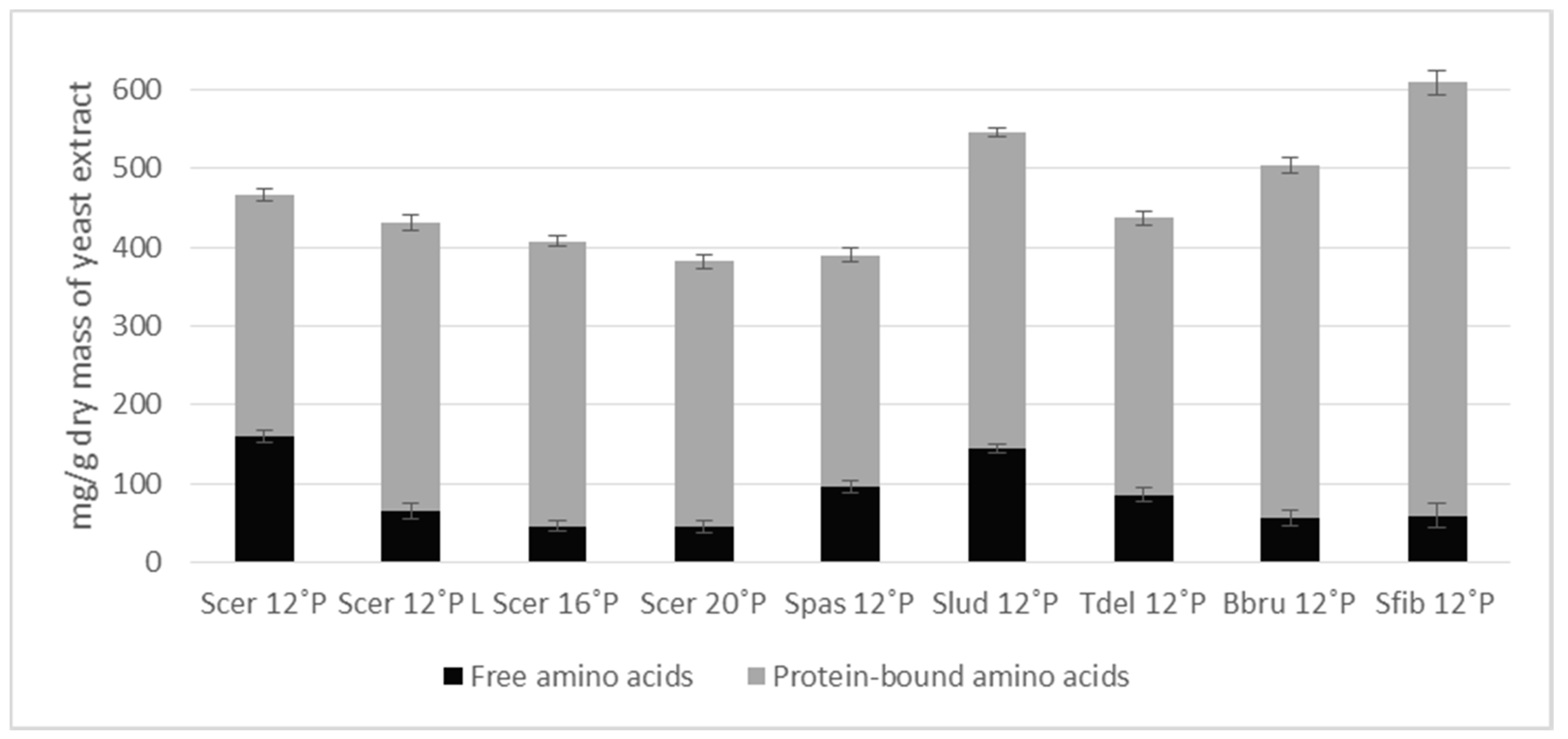
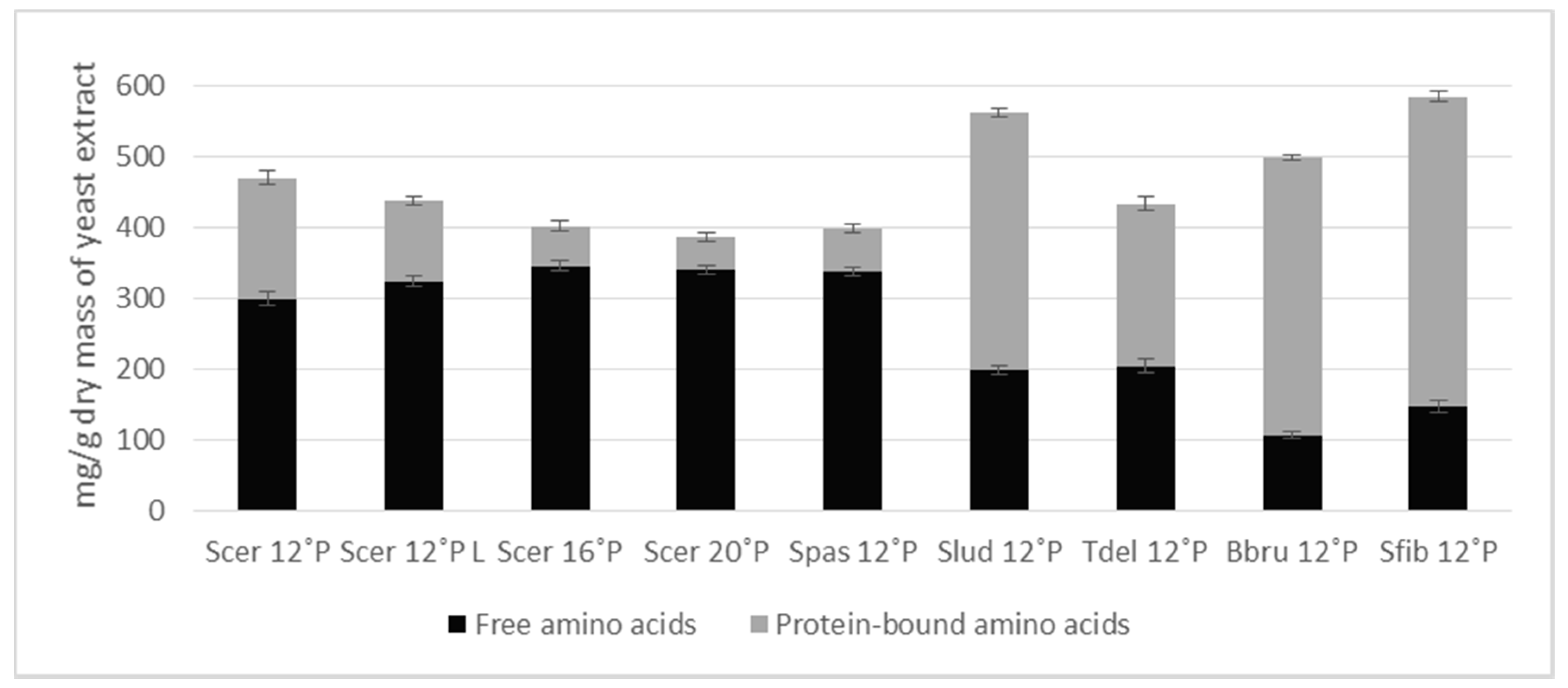
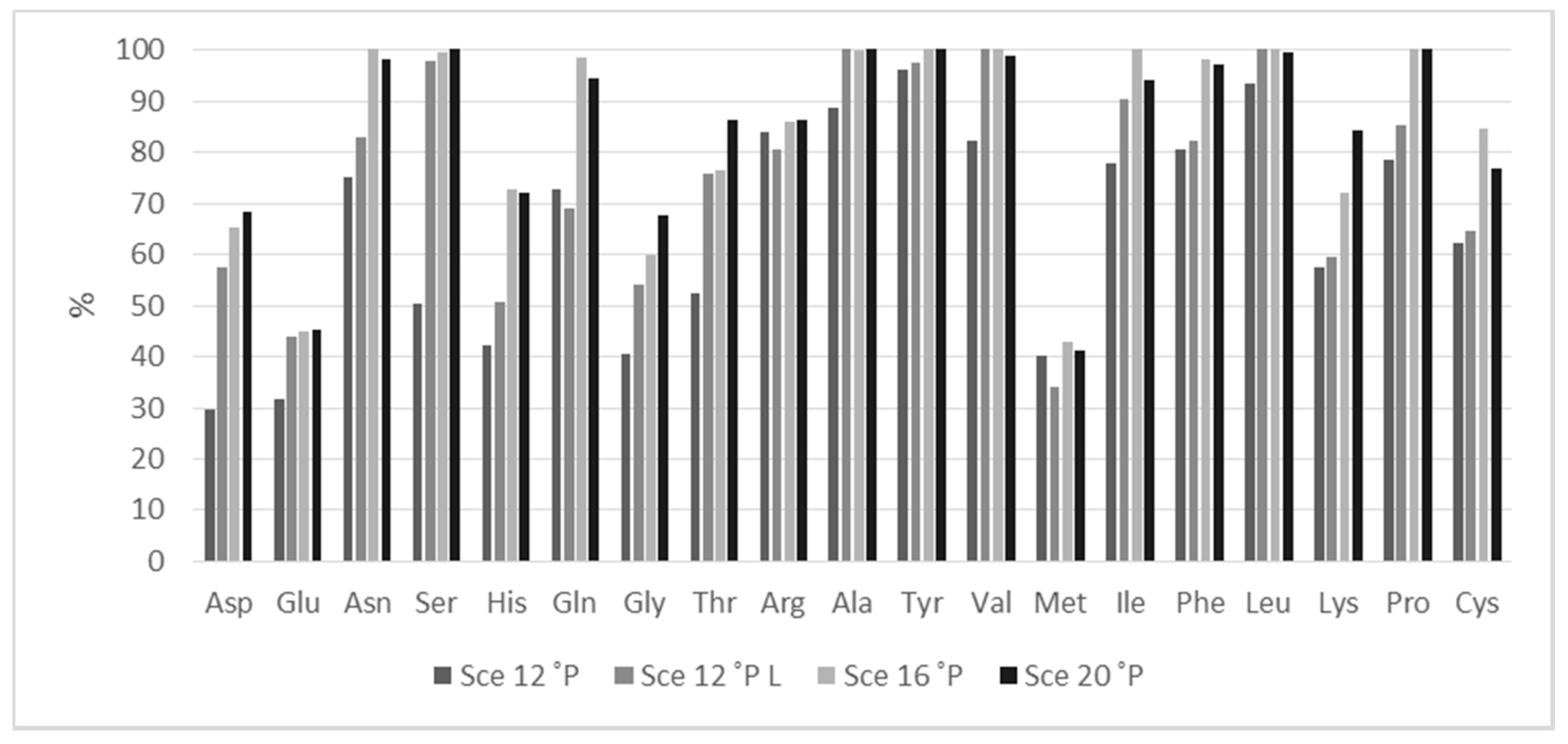
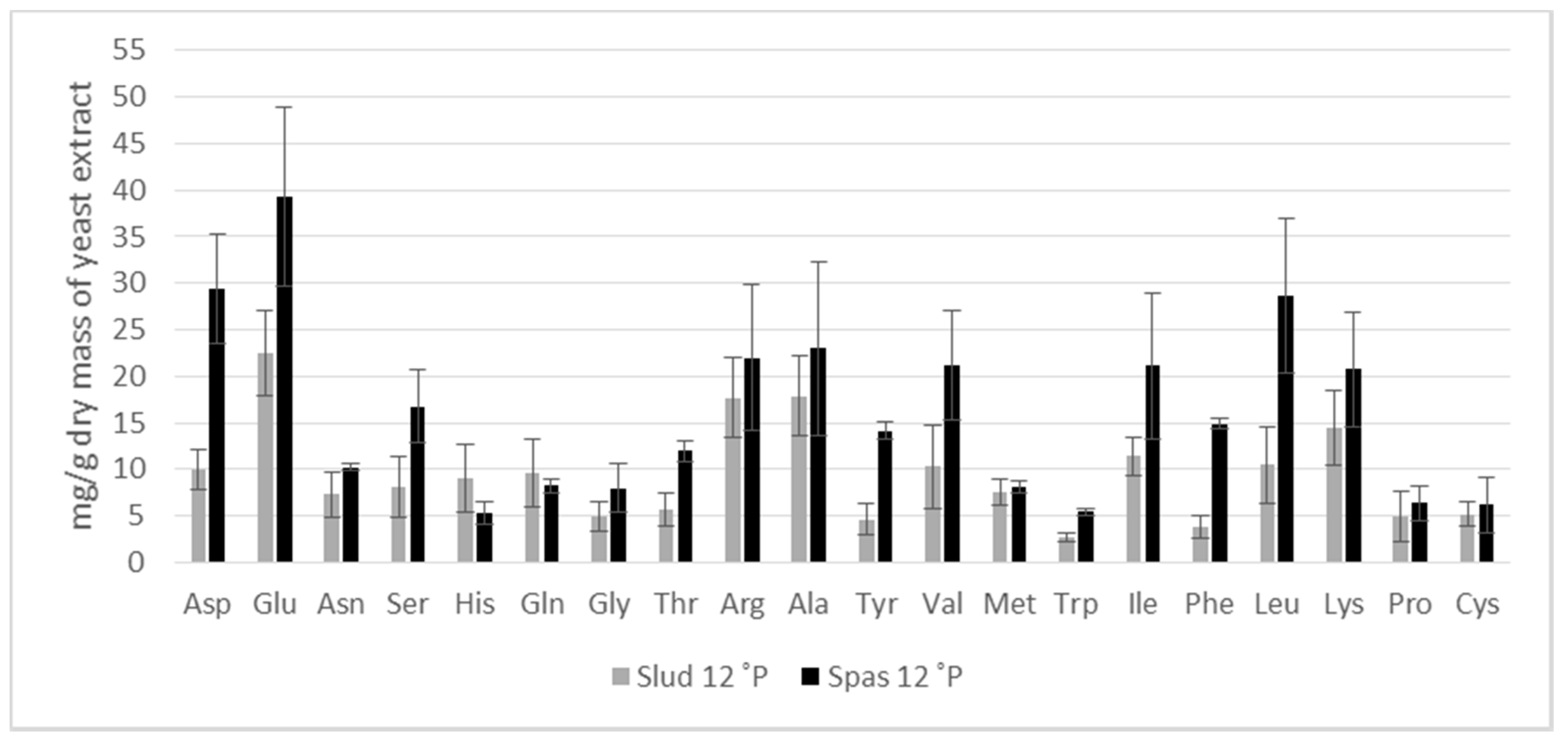
3.2. Amino Acid Composition
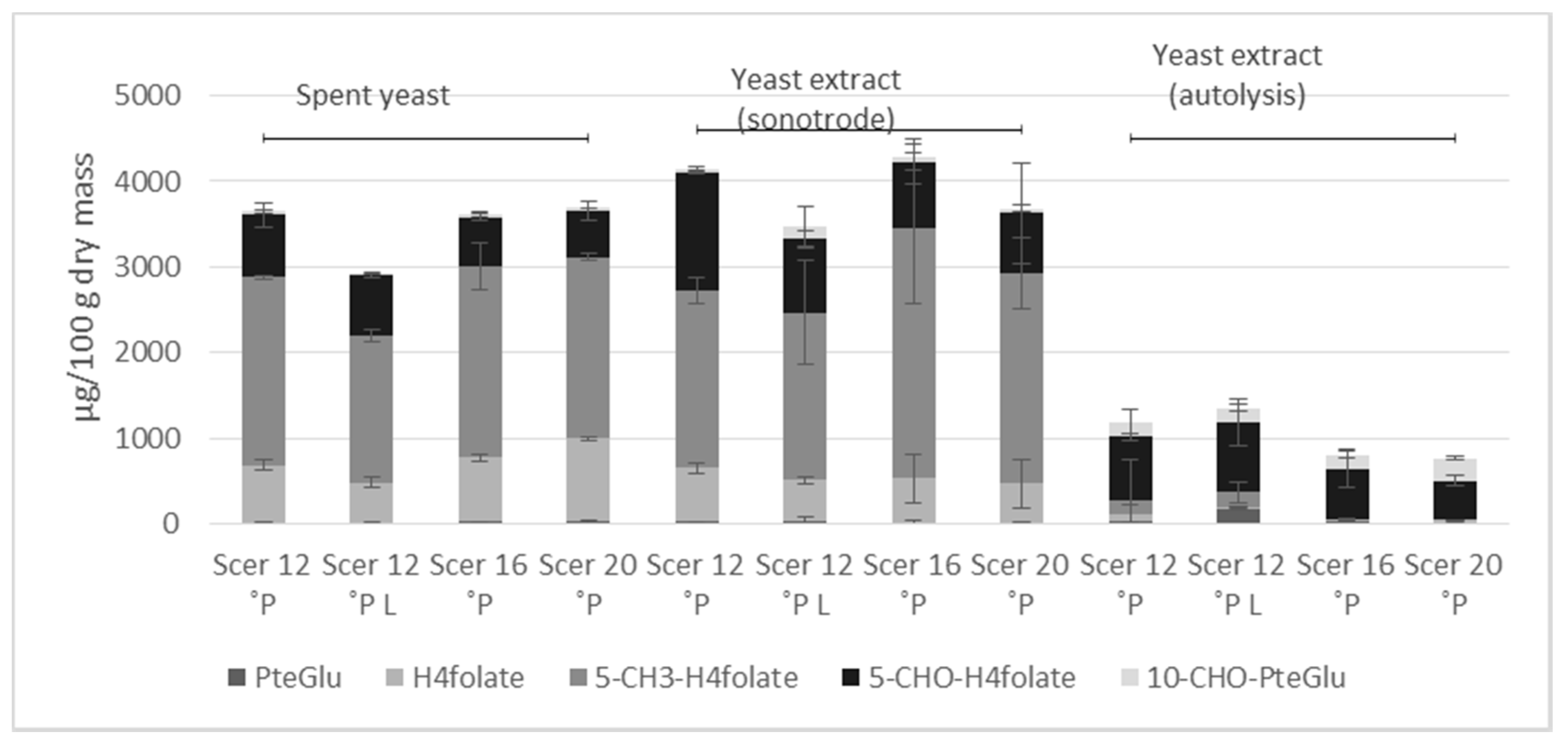
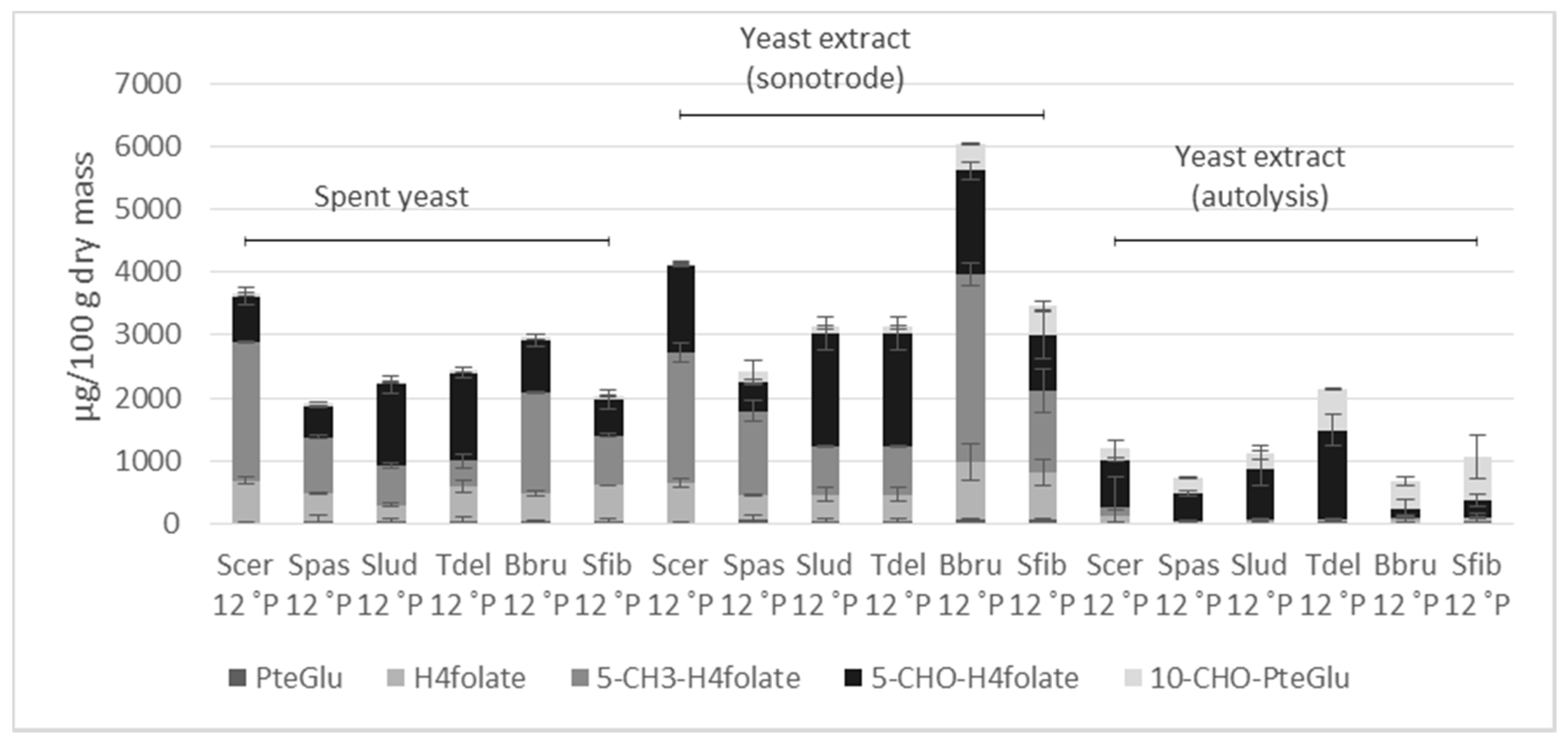
3.3. Folate Vitamer Distribution
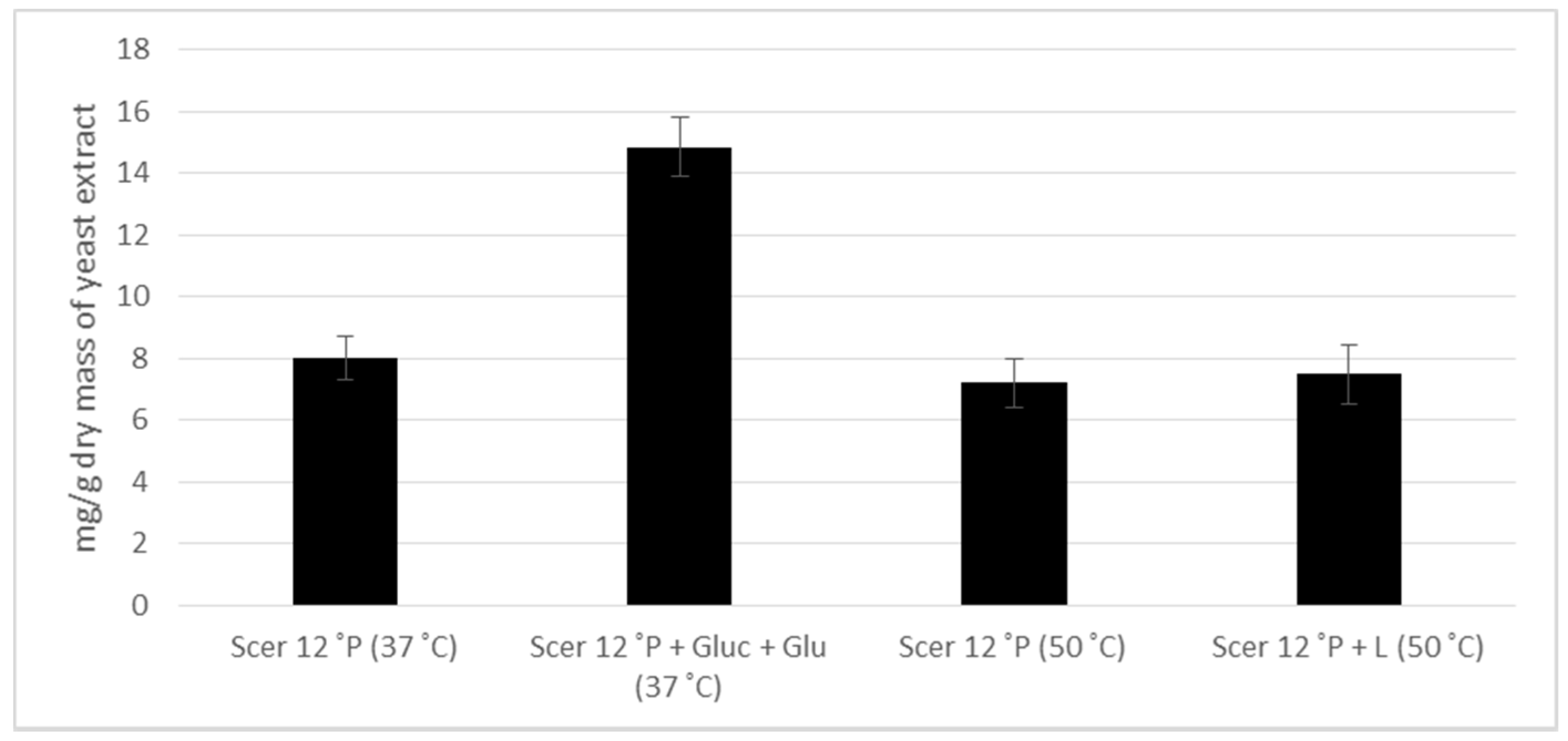
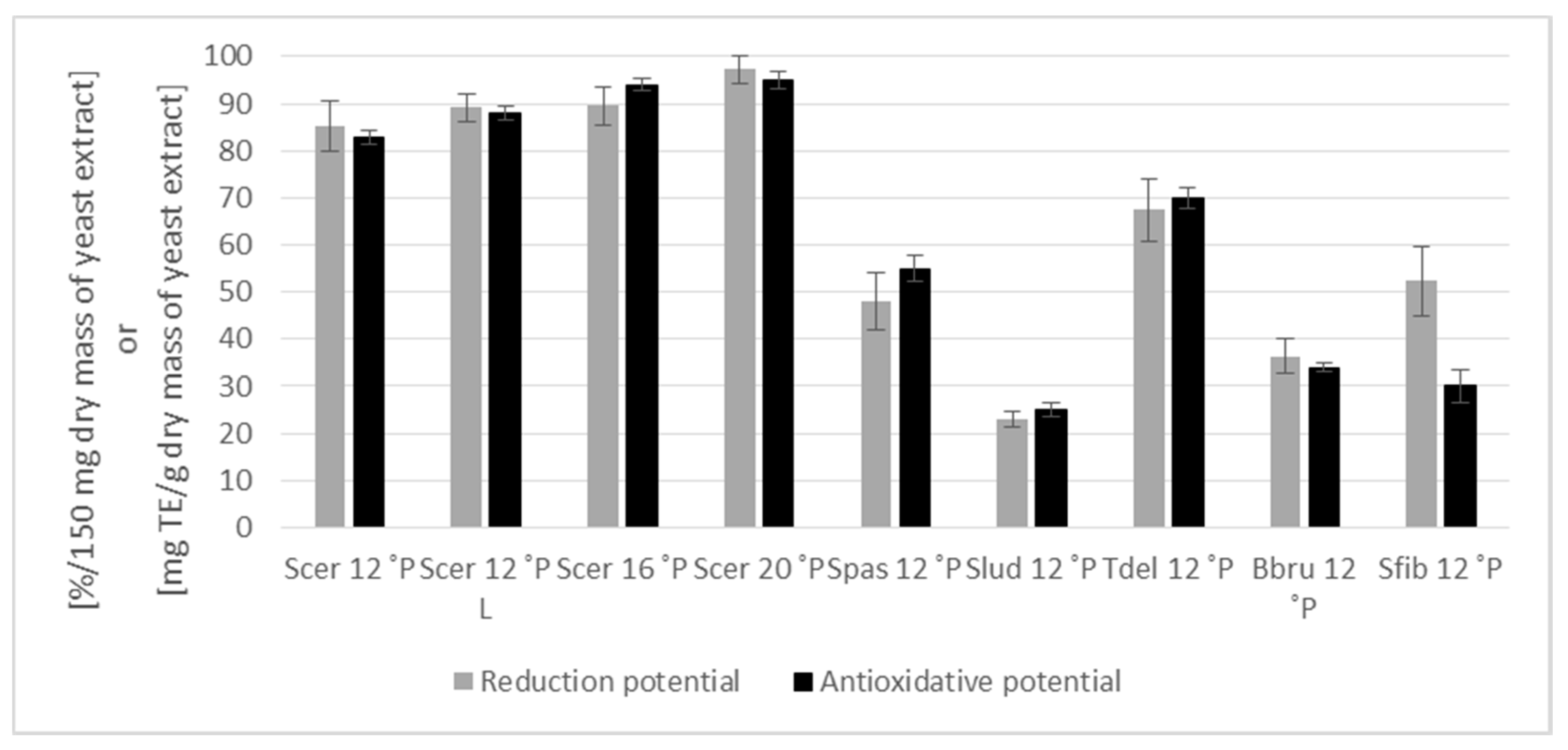
3.4. Biological Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Annemüller, G.; Manger, H.J.; Lietz, P. The Yeast in the Brewery, 1st ed.; VLB Berlin: Berlin, Germany, 2011. [Google Scholar]
- Narziss, L.; Back, W. Die Bierbrauerei Band 2: Die Technologie der Würzebereitung; WILEY-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2009. [Google Scholar]
- Ferreira, I.M.P.L.V.O.; Pinho, O.; Vieira, E.; Tavarela, J.G. Brewer’s Saccharomyces yeast biomass: Characteristics and potential applications. Trends Food Sci. Technol. 2010, 21, 77–84. [Google Scholar] [CrossRef]
- Gibson, B.R. 125th anniversary review: Improvement of higher gravity brewery fermentation via wort enrichment and supplementation. J. Inst. Brew. 2011, 117, 268–284. [Google Scholar] [CrossRef]
- Jacob, F.F.; Hutzler, M.; Methner, F.-J. Comparison of various industrially applicable disruption methods to produce yeast extract using spent yeast from top-fermenting beer production: Influence on amino acid and protein content. Eur. Food Res. Technol. 2019, 245, 95–109. [Google Scholar] [CrossRef]
- Jacob, F.F.; Striegel, L.; Rychlik, M.; Hutzler, M.; Methner, F.J. Yeast extract production using spent yeast from beer manufacture: Influence of industrially applicable disruption methods on selected substance groups with biotechnological relevance. Eur. Food Res. Technol. 2019, 245, 1169–1182. [Google Scholar] [CrossRef]
- Podpora, B.; Świderski, F.; Sadowska, A.; Piotrowska, A.; Rakowska, R. Spent brewer’s yeast autolysates as a new and valuable component of functional food and dietary supplements. J. Food Process. Technol. 2015, 6, 1. [Google Scholar]
- Le Van, V.M.; Strehaiano, P.; Nguyen, D.L.; Taillandier, P. Microbial protease or yeast extract—Alternative additions for improvement of fermentation performance and quality of beer brewed with a high rice content. J. Am. Soc. Brew. Chem. 2001, 59, 10–16. [Google Scholar]
- Casey, G.P.; Magnus, C.A.; Ingledew, W.M. High gravity brewing—nutrient enhanced production of high-concentrations of ethanol by brewing yeast. Biotechnol. Lett. 1983, 5, 429–434. [Google Scholar] [CrossRef]
- Casey, G.P.; Magnus, C.A.; Ingledew, W.M. High-gravity brewing—Effects of nutrition on yeast composition, fermentative ability, and alcohol production. Appl. Environ. Microbiol. 1984, 48, 639–646. [Google Scholar]
- Van Zandycke, S.M.; Fischborn, T. The impact of yeast nutrients on fermentation performance and beer quality. Tech. Q. Master Brew. Assoc. Am. 2008, 45, 290–293. [Google Scholar] [CrossRef]
- Ingledew, W.M.; Sosulski, F.W.; Magnus, C.A. An assessment of yeast foods and their utility in brewing and enology. J. Am. Soc. Brew. Chem. 1986, 44, 166–170. [Google Scholar] [CrossRef]
- Lei, H.J.; Zheng, L.Y.; Wang, C.X.; Zhao, H.F.; Zhao, M.M. Effects of worts treated with proteases on the assimilation of free amino acids and fermentation performance of lager yeast. Int. J. Food Microbiol. 2013, 161, 76–83. [Google Scholar] [CrossRef]
- Procopio, S.; Krause, D.; Hofmann, T.; Becker, T. Significant amino acids in aroma compound profiling during yeast fermentation analyzed by PLS regression. LWT Food Sci. Technol. 2013, 51, 423–432. [Google Scholar] [CrossRef]
- Vieira, F.; Carvalho, J.; Pinto, E.; Cunha, S.; Almeida, A.; Ferreira, I. Nutritive value, antioxidant activity and phenolic compounds profile of brewer’s spent yeast extract. J. Food Compos. Anal. 2016, 52, 44–51. [Google Scholar] [CrossRef]
- Pentieva, K.; McNulty, H.; Reichert, R.; Ward, M.; Strain, J.J.; McKillop, D.J.; McPartlin, J.M.; Connolly, E.; Molloy, A.; Kramer, K.; et al. The short-term bioavailabilities of 6S-5-methyltetrahydrofolate and folic acid are equivalent in men. J. Nutr. 2004, 134, 580–585. [Google Scholar] [CrossRef] [PubMed]
- Jacob, F.F.; Michel, M.; Zarnkow, M.; Hutzler, M.; Methner, F.-J. The complexity of yeast extracts and its consequences on the utility in brewing: A review. BrewingScience 2019, 72, 50–62. [Google Scholar] [CrossRef]
- Piddocke, M.P.; Kreisz, S.; Heldt-Hansen, H.P.; Nielsen, K.F.; Olsson, L. Physiological characterization of brewer’s yeast in high-gravity beer fermentations with glucose or maltose syrups as adjuncts. Appl. Microbiol. Biotechnol. 2009, 84, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Callejo, M.J.; García Navas, J.J.; Alba, R.; Escott, C.; Loira, I.; González, M.C.; Morata, A. Wort fermentation and beer conditioning with selected non-Saccharomyces yeasts in craft beers. Eur. Food Res. Technol. 2018. [Google Scholar] [CrossRef]
- Michel, M.; Meier-Dornberg, T.; Jacob, F.; Methner, F.J.; Wagner, R.S.; Hutzler, M. Review: Pure non-Saccharomyces starter cultures for beer fermentation with a focus on secondary metabolites and practical applications. J. Inst. Brew. 2016, 122, 569–587. [Google Scholar] [CrossRef]
- Barrette, J.; Champagne, C.P.; Goulet, J. Development of bacterial contamination during production of yeast extracts. Appl. Environ. Microbiol. 1999, 65, 3261–3263. [Google Scholar]
- Berlowska, J.; Dudkiewicz-Kołodziejska, M.; Pawlikowska, E.; Pielech-Przybylska, K.; Balcerek, M.; Czysowska, A.; Kregiel, D. Utilization of post-fermentation yeasts for yeast extract production by autolysis: The effect of yeast strain and saponin from Quillaja saponaria. J. Inst. Brew. 2017, 123, 396–401. [Google Scholar] [CrossRef]
- Meier-Dörnberg, T.; Michel, M.; Wagner, R.S.; Jacob, F.; Hutzler, M. Genetic and phenotypic characterization of different top-fermenting Saccharomyces cerevisiae ale yeast isolates. BrewingScience 2017, 70, 9–25. [Google Scholar]
- Masuda, K.; Guo, X.F.; Uryu, N.; Hagiwara, T.; Watabe, S. Isolation of marine yeasts collected from the pacific ocean showing a high production of gamma-aminobutyric acid. Biosci. Biotechnol. Biochem. 2008, 72, 3265–3272. [Google Scholar] [CrossRef] [PubMed]
- Naumann, C.; Bassler, R.; Seibold, R.; Barth, C. Methodenbuch Band 3, die Chemische Untersuchung von Futtermitteln; VDLUFA-Verlag: Darmstadt, Germany, 1976. [Google Scholar]
- Reed, G.; Nagodawithana, T.W. Yeast Technology, 2nd ed.; Van Nostrand Reinhold: New York, NY, USA, 1991. [Google Scholar]
- Jacob, F. MEBAK Brautechnische Analysemethoden Würze Bier Biermischgetränke; Selbstverlag der MEBAK: Freising/Weihenstephan, Germany, 2012. [Google Scholar]
- Striegel, L.; Chebib, S.; Netzel, M.E.; Rychlik, M. Improved stable isotope dilution assay for dietary folates using LC-MS/MS and its application to strawberries. Front. Chem. 2018, 6, 11. [Google Scholar] [CrossRef] [PubMed]
- Caballero-Cordoba, G.M.; Sgarbieri, V.C. Nutritional and toxicological evaluation of yeast (Saccharomyces cerevisiae) biomass and a yeast protein concentrate. J. Sci. Food Agric. 2000, 80, 341–351. [Google Scholar] [CrossRef]
- Pratt, P.L.; Bryce, J.H.; Stewart, G.G. The effects of osmotic pressure and ethanol on yeast viability and morphology. J. Inst. Brew. 2003, 109, 218–228. [Google Scholar] [CrossRef]
- Casey, G.P.; Chen, E.C.H.; Ingledew, W.M. High-gravity brewing: Production of high levels of ethanol without excessive concentrations of esters and fusel alcohols. J. Am. Soc. Brew. Chem. 1985, 43, 179–182. [Google Scholar] [CrossRef]
- Powell, C.D.; Quain, D.E.; Smart, K.A. The impact of brewing yeast cell age on fermentation performance, attenuation and flocculation. FEMS Yeast Res. 2003, 3, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Steckley, J.D.; Grieve, D.G.; Macleod, G.K.; Moran, E.T. Brewers-yeast slurry, 1. Composition as affected by length of storage, temperature, and chemical treatment. J. Dairy Sci. 1979, 62, 941–946. [Google Scholar] [CrossRef]
- Vieira, E.; Cunha, S.C.; Ferreira, I.M.P.L.V.O. Characterization of a potential bioactive food ingredient from inner cellular content of brewer’s spent yeast. Waste Biomass Valori. 2018, 1–8. [Google Scholar] [CrossRef]
- Podpora, B.; Swiderski, F.; Sadowska, A.; Rakowska, R.; Wasiak-Zys, G. Spent brewer’s yeast extracts as a new component of functional food. Czech. J. Food Sci. 2016, 34, 554–563. [Google Scholar] [CrossRef]
- Reeds, P.J. Dispensable and indispensable amino acids for humans. J. Nutr. 2000, 130, 1835–1840. [Google Scholar] [CrossRef] [PubMed]
- Hans, M.A.; Heinzle, E.; Wittmann, C. Quantification of intracellular amino acids in batch cultures of Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2001, 56, 776–779. [Google Scholar] [CrossRef] [PubMed]
- Cooper, D.J.; Stewart, G.G.; Bryce, J.H. Yeast proteolytic activity during high and low gravity wort fermentations and its effect on head retention. J. Inst. Brew. 2000, 106, 197–201. [Google Scholar] [CrossRef]
- Ormrod, I.H.L.; Lalor, E.F.; Sharpe, F.R. The release of yeast proteolytic-enzymes into beer. J. Inst. Brew. 1991, 97, 441–443. [Google Scholar] [CrossRef]
- Fukal, L.; Kas, J.; Rauch, P. Properties of yeast proteinases. J. Inst. Brew. 1986, 92, 357–359. [Google Scholar] [CrossRef]
- Maddox, I.S.; Hough, J.S. Proteolytic enzymes of Saccharomyces carlsbergensis. Biochem. J. 1970, 117, 843–852. [Google Scholar] [CrossRef]
- Lenney, J.F.; Dalbec, J.M. Purification and properties of 2 proteinases from Saccharomyces cerevisiae. Arch. Biochem. Biophys. 1967, 120, 42–48. [Google Scholar] [CrossRef]
- Schneiderbanger, J.; Grammer, M.; Jacob, F.; Hutzler, M. Statistical evaluation of beer spoilage bacteria by real-time PCR analyses from 2010 to 2016. J. Inst. Brew. 2018, 124, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Ohtomo, M.; Kimura, K.; Watanabe, S.; Toeda, K. Production of components containing gamma-aminobutyric acid from rice bran by Lactobacillus brevis IFO12005. Seibutsu-Kogaku Kaishi 2006, 84, 479–483. [Google Scholar]
- Champagne, C.P.; Barrette, J.; Goulet, J. Interaction between pH, autolysis promoters and bacterial contamination on the production of yeast extracts. Food Res. Int. 1999, 32, 575–583. [Google Scholar] [CrossRef]
- Hjortmo, S.; Patring, J.; Jastrebova, J.; Andlid, T. Inherent biodiversity of folate content and composition in yeasts. Trends Food Sci. Technol. 2005, 16, 311–316. [Google Scholar] [CrossRef]
- Hjortmo, S.; Patring, J.; Andlid, T. Growth rate and medium composition strongly affect folate content in Saccharomyces cerevisiae. Int. J. Food Microbiol. 2008, 123, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Kanauchi, O.; Igarashi, K.; Ogata, R.; Mitsuyama, K.; Andoh, A. A yeast extract high in bioactive peptides has a blood-pressure lowering effect in hypertensive model. Curr. Med. Chem. 2005, 12, 3085–3090. [Google Scholar] [CrossRef] [PubMed]
- Vieira, E.F.; Melo, A.; Ferreira, I.M.P.L.V.O. Autolysis of intracellular content of brewer’s spent yeast to maximize ACE-inhibitory and antioxidant activities. LWT Food Sci. Technol. 2017, 82, 255–259. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Amount |
|---|---|
| Original gravity (°P) | 12.00 |
| pH | 5.17 |
| Spec. weight SL 20/20 ˚C | 1.04 |
| Zinc (mg/L) | 0.15 |
| FAN (mg/100 mL) | 25.00 |
| Total AS (mg/100 mL) | 201.38 |
| Total sugar (g/L) | 80.03 |
| EBC-Bittering units (EBU) | 20.00 |
| Glucose (g/L) | 10.46 |
| Fructose (g/L) | 2.17 |
| Sucrose (g/L) | 1.02 |
| Maltose (g/L) | 49.34 |
| Maltotriose (g/L) | 13.79 |
| Sample Name | Yeast Species | Yeast Strain | Original Gravity (˚P) | Apparent Attenuation (%) | Yeast Crop After |
|---|---|---|---|---|---|
| Scer 12˚P | Saccharomyces cerevisiae | TUM 68 | 12 | 79 | F. |
| Scer 12˚P L | Saccharomyces cerevisiae | TUM 68 | 12 | 79 | L. |
| Scer 16˚P | Saccharomyces cerevisiae | TUM 68 | 16 | 85 | F. |
| Scer 20˚P | Saccharomyces cerevisiae | TUM 68 | 20 | 89 | F. |
| Spas 12˚P | Saccharomyces pastorianus | TUM 34/70 | 12 | 80 | F. |
| Slud 12˚P | Saccharomycodes ludwigii | TUM SL 17 | 12 | 11 | F. |
| Tdel 12˚P | Torulaspora delbrueckii | TUM T 90 | 12 | 45 | F. |
| Bbru 12˚P | Brettanomyces bruxellensis | TUM Bretta 1 | 12 | 40 | F. |
| Sfib 12˚P | Saccharomycopsis fibuligera | TUM 525 | 12 | 60 | F. |
| Sample Name | Scer 12˚P | Scer 12˚P L | Scer 16˚P | Scer 20˚P | Spas 12˚P | Slud 12˚P | Tdel 12˚P | Bbru 12˚P | Sfib 12˚P |
|---|---|---|---|---|---|---|---|---|---|
| Protein (mg/g dw) (N × 5.5) | 480.08 ± 10.25 | 448.53 ± 4.93 | 411.80 ± 9.61 | 395,26 ± 5.67 | 411.54 ± 8.62 | 574.28 ± 11.23 | 446.81 ± 8.57 | 508.34 ± 8.75 | 598.01 ± 13.88 |
| Fat (mg/g dw) | 10.6 ± 0.32 | 10.5 ± 0.31 | 10.3 ± 0.36 | 10.4 ± 0,52 | 11.8 ± 0.72 | 15.5 ± 0.43 | 9.88 ± 0.53 | 18.2 ± 0.37 | 9. 68 ± 0.69 |
| Ash (mg/g dw) | 130.55 ± 2.49 | 120.11 ± 3.45 | 84.20 ± 2.73 | 87.10 ± 3.02 | 82.10 ± 1.98 | 110.40 ± 2.57 | 79.12 ± 1.94 | 130.64 ± 2.47 | 89.70 ± 3.24 |
| Cal. (mg/g dw) | 378.77 | 420.86 | 493.70 | 507.24 | 494.56 | 299.82 | 464.19 | 342.82 | 302.61 |
| Moisture (%) | 9.30 ± 0.11 | 9.35 ± 0.13 | 9.41 ± 0.10 | 9.46 ± 0.12 | 9.34 ± 0.13 | 9.43 ± 0.12 | 9.31 ± 0.13 | 9.32 ± 0.11 | 9.29 ± 0.14 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jacob, F.F.; Striegel, L.; Rychlik, M.; Hutzler, M.; Methner, F.-J. Spent Yeast from Brewing Processes: A Biodiverse Starting Material for Yeast Extract Production. Fermentation 2019, 5, 51. https://doi.org/10.3390/fermentation5020051
Jacob FF, Striegel L, Rychlik M, Hutzler M, Methner F-J. Spent Yeast from Brewing Processes: A Biodiverse Starting Material for Yeast Extract Production. Fermentation. 2019; 5(2):51. https://doi.org/10.3390/fermentation5020051
Chicago/Turabian StyleJacob, Friedrich Felix, Lisa Striegel, Michael Rychlik, Mathias Hutzler, and Frank-Jürgen Methner. 2019. "Spent Yeast from Brewing Processes: A Biodiverse Starting Material for Yeast Extract Production" Fermentation 5, no. 2: 51. https://doi.org/10.3390/fermentation5020051
APA StyleJacob, F. F., Striegel, L., Rychlik, M., Hutzler, M., & Methner, F. -J. (2019). Spent Yeast from Brewing Processes: A Biodiverse Starting Material for Yeast Extract Production. Fermentation, 5(2), 51. https://doi.org/10.3390/fermentation5020051