Saccharomyces arboricola and Its Hybrids’ Propensity for Sake Production: Interspecific Hybrids Reveal Increased Fermentation Abilities and a Mosaic Metabolic Profile

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains
2.2. Yeast Isolation
2.3. Serial Spot Dilutions
2.4. Sporulation and Spore Staining
2.5. Interspecific Hybridization
2.6. Genomic DNA Extraction
2.7. Polymerase Chain Reaction
2.8. Sake Fermentations
2.9. Chromatography Analysis
2.9.1. Gas Chromatography-Flame Ionization Detection
2.9.2. Head Space-Solid Phase Microextraction
2.9.3. Gas Chromatography–Mass Spectroscopy
2.9.4. Data Analysis
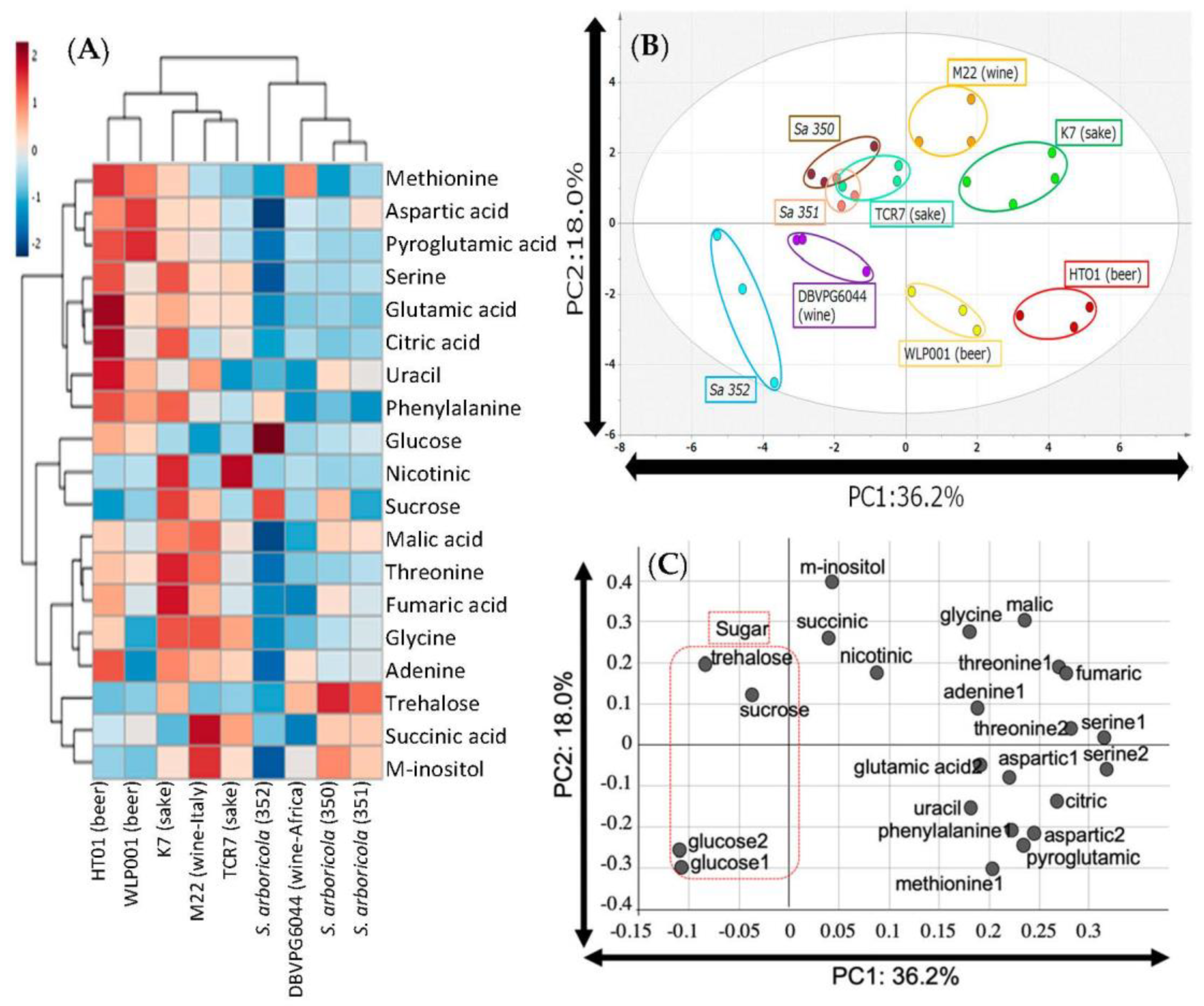
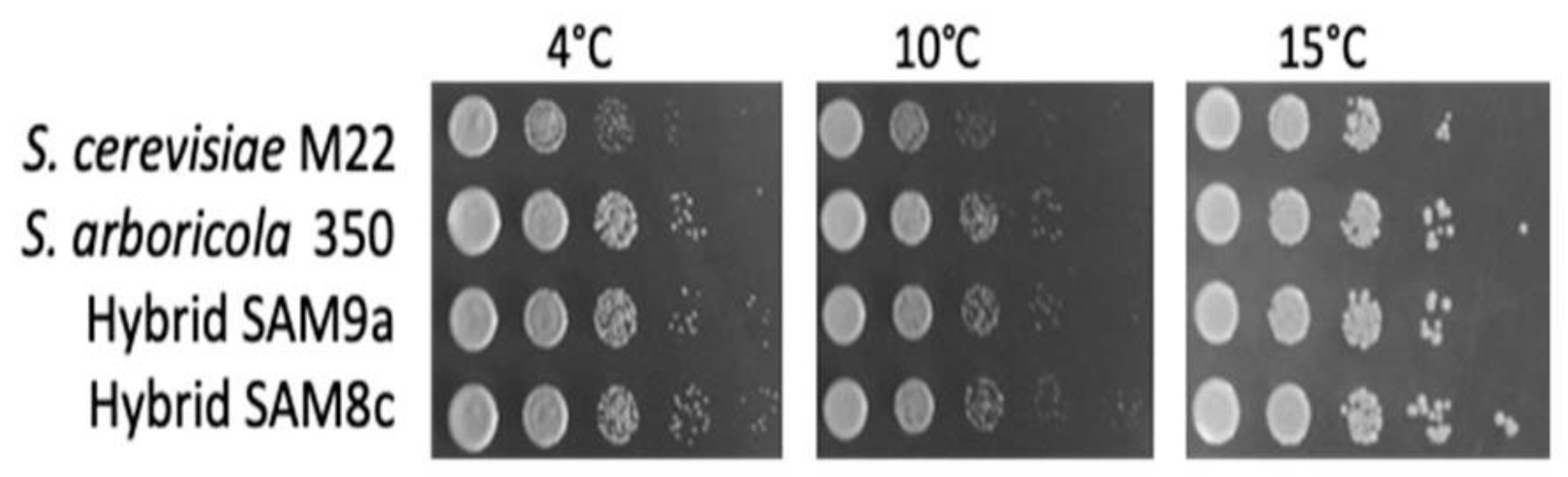
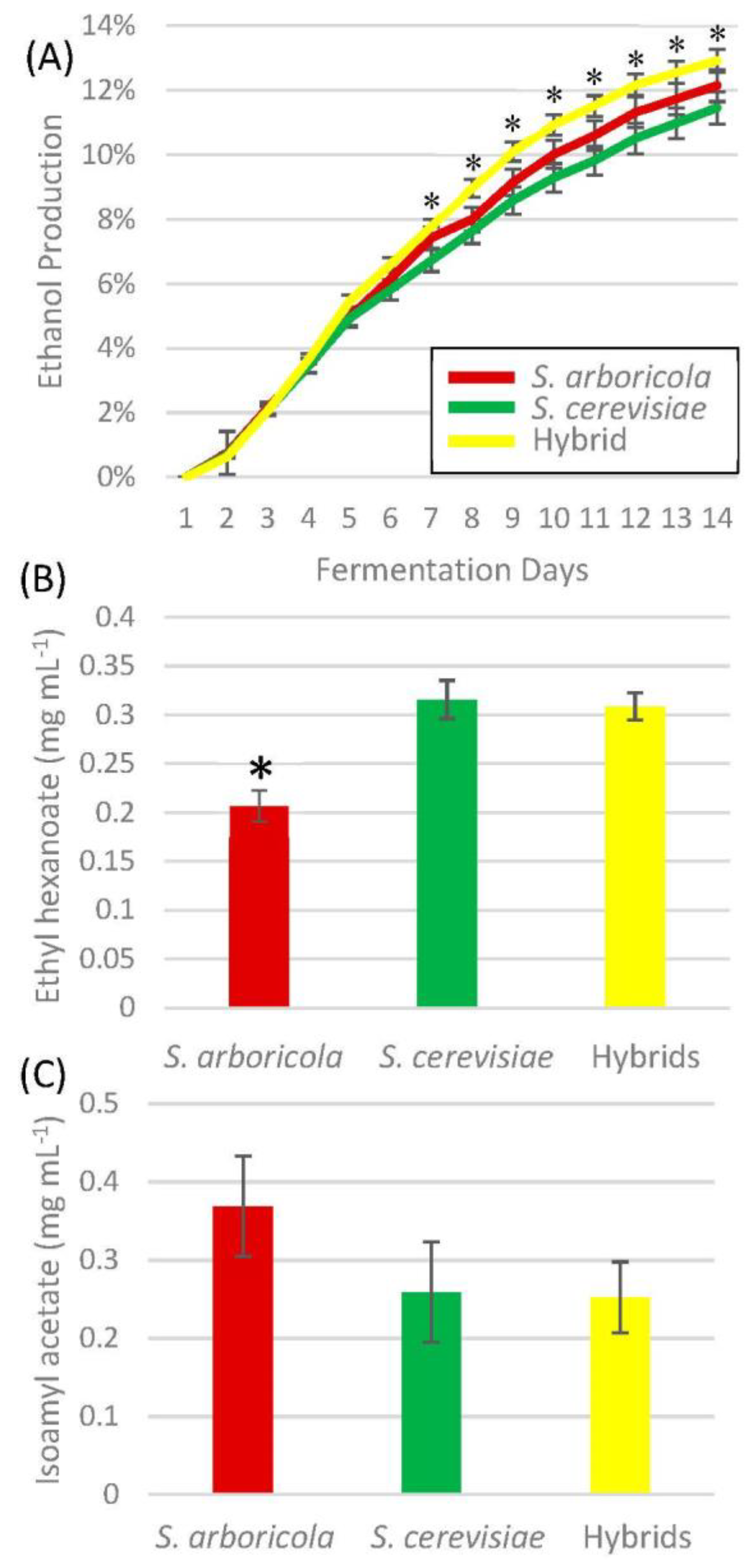
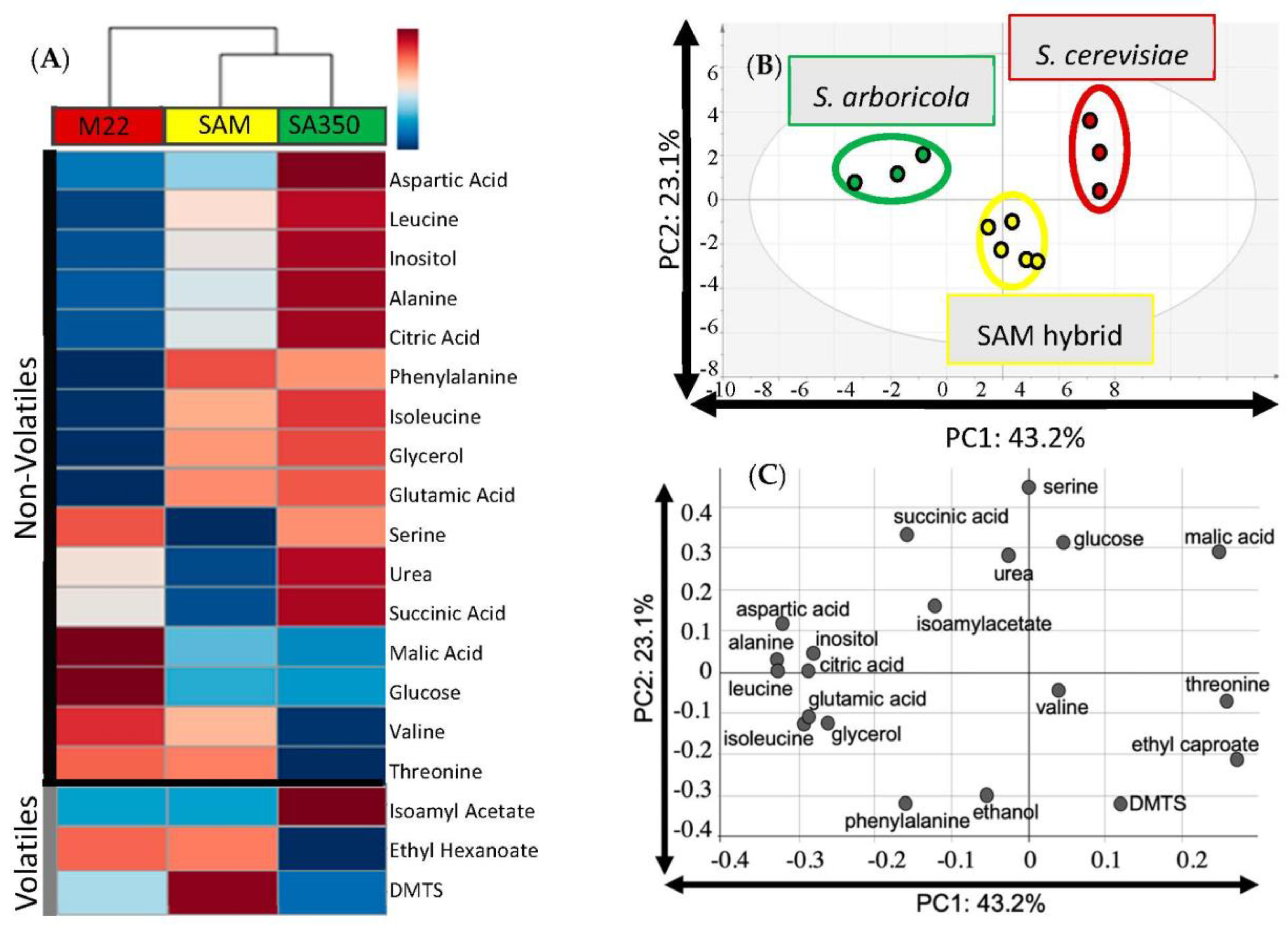
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Martini, A.V.; Martini, A. Three newly delimited species of Saccharomyces sensu stricto. Antonie Van Leeuwenhoek 1987, 53, 77–84. [Google Scholar] [CrossRef]
- Wang, S.A.; Bai, F.Y. Saccharomyces arboricolus sp. nov., a yeast species from tree bark. Int. J. Syst. Evol. Microbiol. 2008, 58, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Naumova, E.S.; Roberts, I.N.; James, S.A.; Naumov, G.I.; Louis, E.J. Three new species in the Saccharomyces sensu stricto complex: Saccharomyces cariocanus, Saccharomyces kudriavzevii and Saccharomyces mikatae. Int. J. Syst. Evol. Microbiol. 2015, 50, 1931–1942. [Google Scholar] [CrossRef]
- Naumov, G.I.; Naumova, E.S.; Hagler, A.N.; Mendonça-Hagler, L.C.; Louis, E.J. A new genetically isolated population of the Saccharomyces sensu stricto complex from Brazil. Antonie Van Leeuwenhoek 1995, 67, 351–355. [Google Scholar] [CrossRef]
- Naumov, G.I.; Lee, C.-F.; Naumova, E.S. Molecular genetic diversity of the Saccharomyces yeasts in Taiwan: Saccharomyces arboricola, Saccharomyces cerevisiae and Saccharomyces kudriavzevii. Antonie Van Leeuwenhoek 2013, 103, 217–228. [Google Scholar] [CrossRef]
- Naumov, G.I.; Naumova, E.S.; Louis, E.J. Two New Genetically Isolated Popoulations of the Saccharomyces Sensu Stricto Complex from Japan. J. Gen. Appl. Microbiol. 1995, 41, 499–505. [Google Scholar] [CrossRef]
- Libkind, D.; Hittinger, C.T.; Valério, E.; Gonçalves, C.; Dover, J.; Johnston, M.; Gonçalves, P.; Sampaio, J.P. Microbe domestication and the identification of the wild genetic stock of lager-brewing yeast. Proc. Natl. Acad. Sci. USA 2011, 108, 14539–14544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boynton, P.J.; Greig, D. The ecology and evolution of non-domesticated Saccharomyces species Primrose. Yeast 2014, 31, 449–462. [Google Scholar] [PubMed] [Green Version]
- Sipiczki, M. Interspecies hybridisation and genome chimerisation in Saccharomyces: Combining of gene pools of species and its biotechnological perspectives. Front. Microbiol. 2018, 9, 3071. [Google Scholar] [CrossRef] [PubMed]
- Masneuf, I.; Hansen, J.; Groth, C.; Piskur, J.; Dubourdieu, D. New hybrids between Saccharomyces sensu stricto yeast species found among wine and cider production strains. Appl. Environ. Microbiol. 1998, 64, 3887–3892. [Google Scholar] [CrossRef] [Green Version]
- Querol, A.; Bond, U. The complex and dynamic genomes of industrial yeasts: Minireview. FEMS Microbiol. Lett. 2009, 293, 1–10. [Google Scholar] [CrossRef]
- Nguyen, H.V.; Legras, J.L.; Neuvéglise, C.; Gaillardin, C. Deciphering the hybridisation history leading to the lager lineage based on the mosaic genomes of Saccharomyces bayanus strains NBRC1948 and CBS380 T. PLoS ONE 2011, 6, e25821. [Google Scholar] [CrossRef] [PubMed]
- Gayevskiy, V.; Goddard, M.R. Saccharomyces eubayanus and Saccharomyces arboricola reside in North Island native New Zealand forests. Environ. Microbiol. 2016, 18, 1137–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liti, G.; Nguyen Ba, A.N.; Blythe, M.; Müller, C.A.; Bergström, A.; Cubillos, F.A.; Dafhnis-Calas, F.; Khoshraftar, S.; Malla, S.; Mehta, N.; et al. High quality de novo sequencing and assembly of the Saccharomyces arboricolus genome. Bmc Genom. 2013, 14, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cliften, P.; Sudarsanam, P.; Desikan, A.; Fulton, L.; Fulton, B.; Majors, J.; Waterston, R.; Cohen, B.A.; Johnston, M. Finding functional features in Saccharomyces genomes by phylogenetic footprinting. Science 2003, 301, 71–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birren, B.; Lander, E.S.; Kellis, M.; Endrizzi, M.; Patterson, N. Sequencing and comparison of yeast species to identify genes and regulatory elements. Nature 2003, 423, 241–254. [Google Scholar]
- Liti, G.; Carter, D.M.; Moses, A.M.; Warringer, J.; Parts, L.; James, S.A.; Davey, R.P.; Roberts, I.N.; Burt, A.; Koufopanou, V.; et al. Population genomics of domestic and wild yeasts. Nature 2009, 458, 337–341. [Google Scholar] [CrossRef] [Green Version]
- Dujon, B. Yeasts illustrate the molecular mechanisms of eukaryotic genome evolution. Trends Genet. 2006, 22, 375–387. [Google Scholar] [CrossRef]
- Postma, E.; Verduyn, C.; Scheffers, A.; Van Dijken, J.P. Enzymic analysis of the Crabtree Effect in glucose-limited chemostat cultures of Saccharomyces cerevisiae. Appl. Environ. Microbiol. 1989, 55, 468–477. [Google Scholar] [CrossRef] [Green Version]
- Galdieri, L.; Mehrotra, S.; Yu, S.; Vancura, A. Transcriptional regulation in yeast during diauxic shift and stationary phase. OMI. J. Integr. Biol. 2010, 14, 629–638. [Google Scholar] [CrossRef] [Green Version]
- McGovern, P.E.; Zhang, J.; Tang, J.; Zhang, Z.; Hall, G.R.; Moreau, R.A.; Nuñez, A.; Butrym, E.D.; Richards, M.P.; Wang, C.-S.; et al. Fermented beverages of pre- and proto-historic China. Proc. Natl. Acad. Sci. USA 2004, 101, 17593–17598. [Google Scholar] [CrossRef] [Green Version]
- McGovern, P.E. Ancient Wine: The Search for the Origins of Viniculture; Princeton Publishing: Princeton, NJ, USA, 2003; pp. 1–15. ISBN 0691070806. [Google Scholar]
- Kitagaki, H.; Kitamoto, K. Breeding Researches of Sake Yeasts in Japan: History, Recent Technological Advances, and Future Perspectives. Food Sci. Technol. 2012, 4, 215–235. [Google Scholar]
- Charles, M.; Tang, H.; Belcram, H.; Paterson, A.; Gornicki, P.; Chalhoub, B. Sixty million years in evolution of soft grain trait in grasses: Emergence of the softness locus in the common ancestor of pooideae and ehrhartoideae, after their divergence from panicoideae. Mol. Biol. Evol. 2009, 26, 1651–1661. [Google Scholar] [CrossRef]
- Arendt, E.; Zannini, E. Rice. In Cereal Grains for the Food and Beverage Industries; Woodhead Publishing: Cambridge, UK, 2013; pp. 114–154. ISBN 9780857098924. [Google Scholar]
- Corran, H.S. A History of Brewing; David & Charles: London, UK, 1975. [Google Scholar]
- Sampaio, J.P. Microbe profile: Saccharomyces eubayanus, the missing link to lager beer yeasts. Microbiolog 2018, 164, 1069–1071. [Google Scholar] [CrossRef] [PubMed]
- Peris, D.; Langdon, Q.K.; Moriarty, R.V.; Sylvester, K.; Bontrager, M.; Charron, G.; Leducq, J.B.; Landry, C.R.; Libkind, D.; Hittinger, C.T. Complex Ancestries of Lager-Brewing Hybrids Were Shaped by Standing Variation in the Wild Yeast Saccharomyces eubayanus. PLoS Genet. 2016, 12, e1006155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nespolo, R.F.; Villarroel, C.A.; Oporto, C.I.; Tapia, S.M.; Vega, F.; Urbina, K.; De Chiara, M.; Mozzachiodi, S.; Mikhalev, E.; Thompson, D.; et al. An Out-of-Patagonia dispersal explains most of the worldwide genetic distribution in Saccharomyces eubayanus. bioRxiv 2019, 709253. [Google Scholar] [CrossRef]
- Langdon, Q.K.; Peris, D.; Eizaguirre, J.I.; Opulente, D.A.; Buh, K.V.; Sylvester, K.; Jarzyna, M.; Rodríguez, M.E.; Lopes, C.A.; Libkind, D.; et al. Genomic diversity and global distribution of Saccharomyces eubayanus, the wild ancestor of hybrid lager-brewing yeasts. bioRxiv 2019, 709535. [Google Scholar]
- Eizaguirre, J.I.; Peris, D.; Rodríguez, M.E.; Lopes, C.A.; De Los Ríos, P.; Hittinger, C.T.; Libkind, D. Phylogeography of the wild Lager-brewing ancestor (Saccharomyces eubayanus) in Patagonia. Environ. Microbiol. 2018, 20, 3732–3743. [Google Scholar] [CrossRef]
- Nagy, L.G.; Ohm, R.A.; Kovács, G.M.; Floudas, D.; Riley, R.; Gácser, A.; Sipiczki, M.; Davis, J.M.; Doty, S.L.; De Hoog, G.S.; et al. Latent homology and convergent regulatory evolution underlies the repeated emergence of yeasts. Nat. Commun. 2014, 5, 4471. [Google Scholar] [CrossRef] [Green Version]
- Bing, J.; Han, P.J.; Liu, W.Q.; Wang, Q.M.; Bai, F.Y. Evidence for a far east asian origin of lager beer yeast. Curr. Biol. 2014, 24, R380–R381. [Google Scholar] [CrossRef] [Green Version]
- Peter, J.; De Chiara, M.; Friedrich, A.; Yue, J.X.; Pflieger, D.; Bergström, A.; Sigwalt, A.; Barre, B.; Freel, K.; Llored, A.; et al. Genome evolution across 1,011 Saccharomyces cerevisiae isolates. Nature 2018, 556, 339–344. [Google Scholar] [CrossRef] [Green Version]
- Duan, S.F.; Han, P.J.; Wang, Q.M.; Liu, W.Q.; Shi, J.Y.; Li, K.; Zhang, X.L.; Bai, F.Y. The origin and adaptive evolution of domesticated populations of yeast from Far East Asia. Nat. Commun. 2018, 9, 2690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Dong, J.; Yin, H.; Zhao, Y.; Chen, R.; Wan, X.; Chen, P.; Hou, X.; Liu, J.; Chen, L. Wort composition and its impact on the flavour-active higher alcohol and ester formation of beer—A review. J. Inst. Brew. 2014, 120, 157–163. [Google Scholar] [CrossRef]
- Engan, S. Wort Composition and Beer Flavour. II. The Influence of Different Carbohydrates on the Formation of Some Flavour Components During Fermentation. J. Inst. Brew. 1971, 78, 169–173. [Google Scholar] [CrossRef]
- Younis, O.S.; Stewart, G.G. Sugar uptake and subsequent ester and higher alcohol production by Saccharomyces cerevisiae. J. Inst. Brew. 1998, 104, 255–264. [Google Scholar] [CrossRef]
- Mortimer, R.; Polsinelli, M. On the origins of wine yeast. Res. Microbiol. 1999, 150, 199–204. [Google Scholar] [CrossRef]
- Bisson, L.F. Gene Expression in Yeasts During Wine Fermentation. In Yeasts in The Production of Wine; Romano, P., Ciani, M., Fleet, G., Eds.; Springer: New York, NY, USA, 2019; pp. 165–209. [Google Scholar]
- Kühbeck, F.; Back, W.; Krottenthaler, M. Influence of lauter turbidity on wort composition, fermentation performance and beer quality—A review. J. Inst. Brew. 2006, 112, 215–221. [Google Scholar] [CrossRef]
- White, C.; Zainasheff, J. Yeast: The Practical Guide to Beer Fermentation (Brewing Elements); Brewers Publications: Boulder, CO, USA, 2010. [Google Scholar]
- Kodama, K. Sake Yeast. In The Yeasts; Rose, H., Harrison, J.S., Eds.; Academic Press: London, UK, 1993; pp. 129–168. [Google Scholar]
- Ferdouse, J.; Yamamoto, Y.; Taguchi, S.; Yoshizaki, Y.; Takamine, K.; Kitagaki, H. Glycosylceramide modifies the flavor and metabolic characteristics of sake yeast. PeerJ 2018, 10, e4768. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Guan, Z.B.; Cao, Y.; Xie, G.F.; Lu, J. Secretome of Aspergillus oryzae in Shaoxing rice wine koji. Int. J. Food Microbiol. 2012, 155, 113–119. [Google Scholar] [CrossRef]
- Song, S.H.; Vieille, C. Recent advances in the biological production of mannitol. Appl. Microbiol. Biotechnol. 2009, 84, 55–62. [Google Scholar] [CrossRef]
- Smiley, K.L.; Cadmus, M.C.; Liepins, P. Biosynthesis of D-mannitol from D-glucose by Aspergillus candidus. Biotechnol. Bioeng. 1967, 9, 365–374. [Google Scholar] [CrossRef]
- Bellon, J.R.; Schmid, F.; Capone, D.L.; Dunn, B.L.; Chambers, P.J. Introducing a New Breed of Wine Yeast: Interspecific Hybridisation between a Commercial Saccharomyces cerevisiae Wine Yeast and Saccharomyces mikatae. PLoS ONE 2013, 8, e62053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, S.S.; Barrio, E.; Gafner, J.; Querol, A. Natural hybrids from Saccharomyces cerevisiae, Saccharomyces bayanus and Saccharomyces kudriavzevii in wine fermentations. FEMS Yeast Res. 2006, 6, 1221–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capece, A.; Romaniello, R.; Siesto, G.; Romano, P. Diversity of Saccharomyces cerevisiae yeasts associated to spontaneously fermenting grapes from an Italian “heroic vine-growing area”. Food Microbiol. 2012, 31, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Dickins, B.J.A.; Demeter, J.; Engel, S.; Dunn, B.; Cherry, J.M. AGAPE (Automated Genome Analysis PipelinE) for pan-genome analysis of Saccharomyces cerevisiae. PLoS ONE 2015, 10, e0120671. [Google Scholar] [CrossRef] [Green Version]
- Katou, T.; Kitagaki, H.; Akao, T.; Shimoi, H. Brewing characteristics of haploid strains isolated from sake yeast Kyokai No. Yeast 2008, 25, 799–807. [Google Scholar] [CrossRef]
- Horie, K.; Oba, T.; Motomura, S.; Isogai, A.; Yoshimura, T.; Tsuge, K.; Koganemaru, K.; Kobayashi, G.; Kitagaki, H. Breeding of a Low Pyruvate-Producing Sake Yeast by Isolation of a Mutant Resistant to Ethyl α-Transcyanocinnamate, an Inhibitor of Mitochondrial Pyruvate Transport. Biosci. Biotechnol. Biochem. 2010, 74, 843–847. [Google Scholar] [CrossRef]
- Sniegowski, P.D.; Dombrowski, P.G.; Fingerman, E. Saccharomyces cerevisiae and Saccharomyces paradoxus coexist in a natural woodland site in North America and display different levels of reproductive isolation from European conspecifics. FEMS Yeast Res. 2002, 1, 299–306. [Google Scholar]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Consortium, F.B.; Bolchacova, E.; Voigt, K.; et al. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Rong-Mullins, X.; Winans, M.J.; Lee, J.B.; Lonergan, Z.R.; Pilolli, V.A.; Weatherly, L.M.; Carmenzind, T.W.; Jiang, L.; Cumming, J.R.; Oporto, G.S.; et al. Proteomic and genetic analysis of the response of S. cerevisiae to soluble copper leads to improvement of the antimicrobial function of cellulosic copper nanoparticles. Metallomics 2017, 9, 1304–1315. [Google Scholar] [CrossRef] [Green Version]
- Shimwell, J.L. A Simple Staining Method for the Detection of Ascospores in Yeasts. J. Inst. Brew. 1938, 44, 474. [Google Scholar] [CrossRef]
- Muir, A.; Harrison, E.; Wheals, A. A multiplex set of species-specific primers for rapid identification of members of the genus Saccharomyces. FEMS Yeast Res. 2011, 11, 552–563. [Google Scholar] [CrossRef] [PubMed]
- Korbie, D.J.; Mattick, J.S. Touchdown PCR for increased specificity and sensitivity in PCR amplification. Nat. Protoc. 2008, 3, 1452–1456. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.Y.; Costumbrado, J.; Hsu, C.-Y.; Kim, Y.H. Agarose Gel Electrophoresis for the Separation of DNA Fragments. J. Vis. Exp. 2012, 20, e3923. [Google Scholar] [CrossRef]
- Xia, J.; Sinelnikov, I.V.; Han, B.; Wishart, D.S. MetaboAnalyst 3.0-making metabolomics more meaningful. Nucleic Acids Res. 2015, 43, W251–W257. [Google Scholar] [CrossRef] [Green Version]
- Berthels, N.J.; Cordero Otero, R.R.; Bauer, F.F.; Thevelein, J.M.; Pretorius, I.S. Discrepancy in glucose and fructose utilisation during fermentation by Saccharomyces cerevisiae wine yeast strains. FEMS Yeast Res. 2004, 4, 683–689. [Google Scholar] [CrossRef] [Green Version]
- Berthels, N.J.; Cordero Otero, R.R.; Bauer, F.F.; Pretorius, I.S.; Thevelein, J.M. Correlation between glucose/fructose discrepancy and hexokinase kinetic properties in different Saccharomyces cerevisiae wine yeast strains. Appl. Microbiol. Biotechnol. 2008, 77, 1083–1091. [Google Scholar] [CrossRef]
- Melcher, K.; Entian, K.-D. Genetic analysis of serine biosynthesis and glucose repression in yeast. Curr. Genet. 1992, 21, 295–300. [Google Scholar] [CrossRef]
- Gallone, B.; Steensels, J.; Baele, G.; Maere, S.; Verstrepen, K.J.; Prahl, T.; Soriaga, L.; Saels, V.; Herrera-Malaver, B.; Merlevede, A.; et al. Domestication and Divergence of Saccharomyces cerevisiae Beer Yeasts. Cell 2016, 166, 1397–1410.e16. [Google Scholar] [CrossRef] [Green Version]
- Dunn, B.; Richter, C.; Kvitek, D.J.; Pugh, T.; Sherlock, G. Analysis of the Saccharomyces cerevisiae pan-genome reveals a pool of copy number variants distributed in diverse yeast strains from differing industrial environments. Genome Res. 2012, 22, 908–924. [Google Scholar] [CrossRef] [Green Version]
- Bergström, A.; Simpson, J.T.; Salinas, F.; Barré, B.; Parts, L.; Zia, A.; Nguyen Ba, A.N.; Moses, A.M.; Louis, E.J.; Mustonen, V.; et al. A high-definition view of functional genetic variation from natural yeast genomes. Mol. Biol. Evol. 2014, 31, 872–888. [Google Scholar] [CrossRef] [PubMed]
- Verstrepen, K.J.; Derdelinckx, G.; Dufour, J.P.; Winderickx, J.; Thevelein, J.M.; Pretorius, I.S.; Delvaux, F.R. Flavor-active esters: Adding fruitiness to beer. J. Biosci. Bioeng. 2003, 96, 110–118. [Google Scholar] [CrossRef]
- Pisarnitskii, A.F. Formation of wine aroma: Tones and imperfections caused by minor components (review). Appl. Biochem. Microbiol. 2001, 37, 552–560. [Google Scholar] [CrossRef]
- Pretorius, I.S.; Lambrechts, M.G. Yeast and its importance to wine aroma: A review. S. Afr. J. Enol. Vitic. 2000, 21, 97–129. [Google Scholar]
- Quilter, M.G.; Hurley, J.C.; Lynch, F.J.; Murphy, M.G. The Production of Isoamyl Acetate from Amyl Alcohol by Saccharomyces cerevisiae. J. Inst. Brew. 2003, 109, 34–40. [Google Scholar] [CrossRef]
- Verstrepen, K.J.; Van Laere, S.D.M.; Vanderhaegen, B.M.P.; Derdelinckx, G.; Dufour, J.-P.; Pretorius, I.S.; Winderickx, J.; Thevelein, J.M.; Delvaux, F.R. Expression Levels of the Yeast Alcohol Acetyltransferase Genes ATF1, Lg-ATF1, and ATF2 Control the Formation of a Broad Range of Volatile Esters. Appl. Environ. Microbiol. 2003, 69, 5228–5237. [Google Scholar] [CrossRef] [Green Version]
- Norstedt, C.; Bengtsson, A.; Bennet, P.; Lindström, I.; Äyräpää, T. Technical Measures to Control the Formation of Esters During Beer Fermentation. In Proceedings of the European Brewery Convention: Proceedings of the 15th Congress; Elsevier Scientific Publishing Company: Amsterdam, The Netherlands, 1975; pp. 581–600. [Google Scholar]
- Hammond, J.R.M. Yeast Technology. In Brewer’s Yeast; Rose, A.H., Harrison, J.S., Eds.; Academic Press: San Diego, CA, USA, 1993; Volumne 5, pp. 8–56. [Google Scholar]
- Takahashi, T.; Ohra, Y.; Sueno, K. Breeding of a sake yeast mutant with enhanced ethyl caproate productivity in sake brewing using rice milled at a high polishing ratio. J. Biosci. Bioeng. 2017, 123, 707–713. [Google Scholar] [CrossRef]
- Furukawa, K.; Yamada, T.; Mizoguchi, H.; Hara, S. Increased Ethyl Caproate Production by Inositol Limitation in Saccharomyces cerevisiae. J. Biosci. Bioeng. 2003, 95, 448–454. [Google Scholar] [CrossRef]
- Dittrich-Reed, D.R.; Fitzpatrick, B.M. Transgressive Hybrids as Hopeful Monsters. Evol. Biol. 2013, 40, 310–315. [Google Scholar] [CrossRef] [Green Version]
- Bisson, L.F. Yeast Hybrids in Winemaking. Catal. Discov. Pract. 2017, 1, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Morales, L.; Dujon, B. Evolutionary Role of Interspecies Hybridization and Genetic Exchanges in Yeasts. Microbiol. Mol. Biol. Rev. 2012, 76, 721–739. [Google Scholar] [CrossRef] [Green Version]
- Nikulin, J.; Krogerus, K.; Gibson, B. Alternative Saccharomyces interspecies hybrid combinations and their potential for low-temperature wort fermentation. Yeast 2018, 35, 113–127. [Google Scholar] [CrossRef] [Green Version]
- Peris, D.; Moriarty, R.V.; Alexander, W.G.; Baker, E.C.; Sylvester, K.; Sardi, M.; Langdon, Q.K.; Libkind, D.; Wang, Q.M.; Bai, F.Y.; et al. Hybridization and adaptive evolution of diverse Saccharomyces species for cellulosic biofuel production. Biotechnol. Biofuels 2017, 10, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvadó, Z.; Arroyo-López, F.N.; Guillamón, J.M.; Salazar, G.; Querol, A.; Barrio, E. Temperature adaptation Markedly Determines evolution within the genus Saccharomyces. Appl. Environ. Microbiol. 2011, 77, 2292–2302. [Google Scholar] [CrossRef] [Green Version]
- Krogerus, K.; Magalhes, F.; Vidgren, V.; Gibson, B. New lager yeast strains generated by interspecific hybridization. J. Ind. Microbiol. Biotechnol. 2015, 769–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magalhães, F.; Krogerus, K.; Vidgren, V.; Sandell, M.; Gibson, B. Improved cider fermentation performance and quality with newly generated Saccharomyces cerevisiae × Saccharomyces eubayanus hybrids. J. Ind. Microbiol. Biotechnol. 2017, 44, 1203–1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magalhães, F.; Krogerus, K.; Castillo, S.; Ortiz-Julien, A.; Dequin, S.; Gibson, B. Exploring the potential of Saccharomyces eubayanus as a parent for new interspecies hybrid strains in winemaking. FEMS Yeast Res. 2017, 17, 1–10. [Google Scholar] [CrossRef]
- Torija, M.J.; Beltran, G.; Novo, M.; Poblet, M.; Guillamón, J.M.; Mas, A.; Rozès, N. Effects of fermentation temperature and Saccharomyces species on the cell fatty acid composition and presence of volatile compounds in wine. Int. J. Food Microbiol. 2003, 85, 127–136. [Google Scholar] [CrossRef]
- Krogerus, K.; Magalhães, F.; Vidgren, V.; Gibson, B. Novel brewing yeast hybrids: Creation and application. Appl. Microbiol. Biotechnol. 2017, 101, 65–78. [Google Scholar] [CrossRef] [Green Version]
- Fay, J.C.; Hittinger, C.T.; Sia, E.A.; Peris, D.; Li, X.C. Mitochondria-encoded genes contribute to evolution of heat and cold tolerance in yeast. Sci. Adv. 2019, 5, eaav1848. [Google Scholar]
- Beltran, G.; Novo, M.; Guillamón, J.M.; Mas, A.; Rozès, N. Effect of fermentation temperature and culture media on the yeast lipid composition and wine volatile compounds. Int. J. Food Microbiol. 2008, 121, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Redón, M.; Guillamón, J.M.; Mas, A.; Rozès, N. Effect of lipid supplementation upon Saccharomyces cerevisiae lipid composition and fermentation performance at low temperature. Eur. Food Res. Technol. 2009, 228, 833–840. [Google Scholar] [CrossRef]
- Baker, E.P.; Peris, D.; Moriarty, R.V.; Li, X.C.; Fay, J.C.; Hittinger, C.T. Mitochondrial DNA and temperature tolerance in lager yeasts. Sci. Adv. 2019, 5, eaav1869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tronchoni, J.; Rozès, N.; Querol, A.; Guillamón, J.M. Lipid composition of wine strains of Saccharomyces kudriavzevii and Saccharomyces cerevisiae grown at low temperature. Int. J. Food Microbiol. 2012, 155, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Iwase, T.; Takada, A.; Watanade, T.; Fukuda, H.; Sasaki, K.; Yoshitake, M. Breeding of Sake Yeast by Protoplast Fusion of Sake Yeast and Wine Yeast. J. Brew. Soc. Jpn. 1995, 90, 137–142. [Google Scholar] [CrossRef] [Green Version]
- Motomura, S.; Horie, K.; Kitagaki, H. Mitochondrial activity of sake brewery yeast affects malic and succinic acid production during alcoholic fermentation. J. Inst. Brew. 2012, 118, 22–26. [Google Scholar] [CrossRef]
- Asano, T.; Kurose, N.; Hiraoka, N.; Kawakita, S. Effect of NAD+-dependent isocitrate dehydrogenase gene (IDH1, IDH2) disruption of sake yeast on organic acid composition in sake mash. J. Biosci. Bioeng. 1999, 88, 258–263. [Google Scholar] [CrossRef]
- Wakabayashi, K.; Isogai, A.; Watanabe, D.; Fujita, A.; Sudo, S. Involvemwnt of methionine salvage pathway genes of Saccharomyces cerevisiae in the production of precursor compounds of dimethyl trisulfide (DMTS). J. Biosci. Bioeng. 2013, 116, 475–479. [Google Scholar] [CrossRef]
- Schneider, B.L.; Kiupakis, A.K.; Reitzer, L.J. Arginine catabolism and the arginine succinyltransferase pathway in Escherichia coli. J. Bacteriol. 1998, 180, 4278–4286. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Species | Isolation Source | Reference |
|---|---|---|---|
| SA350 (syn. AS 2.3317) | S. arboricola | Tree bark-China | Liti, G., et al. (2013) |
| SA351 (syn. AS 2.3318) | S. arboricola | Tree bark-China | Liti, G., et al. (2013) |
| SA352 (syn. AS 2.3319) | S. arboricola | Tree bark-China | Liti, G., et al. (2013) |
| M22 | S. cerevisiae | Wine-Italy | Capece, A., et al. (2012) |
| DBVPG6044 | S. cerevisiae | Billi wine-West Africa | Song, G., et al. (2015) |
| Kyokai No. 7 (syn. 868K7/ K7) | S. cerevisiae | Sake-Japan | Katou, T., et. al. (2008) |
| TCR7 | S. cerevisiae | Sake-Japan | Horie, K., et al. (2010) |
| WLP001 | S. cerevisiae | Beer, American ale | Commercially available from White Labs, USA |
| HT01 | S. cerevisiae | Beer, American ale | Current study |
| SAM8c | Hybrid (S. arboricola x S. cerevisiae) | Lab, spore to spore mating | Current study |
| SAM9a | Hybrid (S. arboricola x S. cerevisiae) | Lab, spore to spore mating | Current study |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Winans, M.J.; Yamamoto, Y.; Fujimaru, Y.; Kusaba, Y.; Gallagher, J.E.G.; Kitagaki, H. Saccharomyces arboricola and Its Hybrids’ Propensity for Sake Production: Interspecific Hybrids Reveal Increased Fermentation Abilities and a Mosaic Metabolic Profile. Fermentation 2020, 6, 14. https://doi.org/10.3390/fermentation6010014
Winans MJ, Yamamoto Y, Fujimaru Y, Kusaba Y, Gallagher JEG, Kitagaki H. Saccharomyces arboricola and Its Hybrids’ Propensity for Sake Production: Interspecific Hybrids Reveal Increased Fermentation Abilities and a Mosaic Metabolic Profile. Fermentation. 2020; 6(1):14. https://doi.org/10.3390/fermentation6010014
Chicago/Turabian StyleWinans, Matthew J., Yuki Yamamoto, Yuki Fujimaru, Yuki Kusaba, Jennifer E. G. Gallagher, and Hiroshi Kitagaki. 2020. "Saccharomyces arboricola and Its Hybrids’ Propensity for Sake Production: Interspecific Hybrids Reveal Increased Fermentation Abilities and a Mosaic Metabolic Profile" Fermentation 6, no. 1: 14. https://doi.org/10.3390/fermentation6010014
APA StyleWinans, M. J., Yamamoto, Y., Fujimaru, Y., Kusaba, Y., Gallagher, J. E. G., & Kitagaki, H. (2020). Saccharomyces arboricola and Its Hybrids’ Propensity for Sake Production: Interspecific Hybrids Reveal Increased Fermentation Abilities and a Mosaic Metabolic Profile. Fermentation, 6(1), 14. https://doi.org/10.3390/fermentation6010014