Co-Existence of Inoculated Yeast and Lactic Acid Bacteria and Their Impact on the Aroma Profile and Sensory Traits of Tempranillo Red Wine

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. Vinifications
2.3. Microbiological Control
2.4. FML Progress and Physicochemical and Spectrophotometric Analysis
2.5. Volatile Compound Analysis
2.6. Sensory Analysis
2.7. Statistical Analysis
3. Results
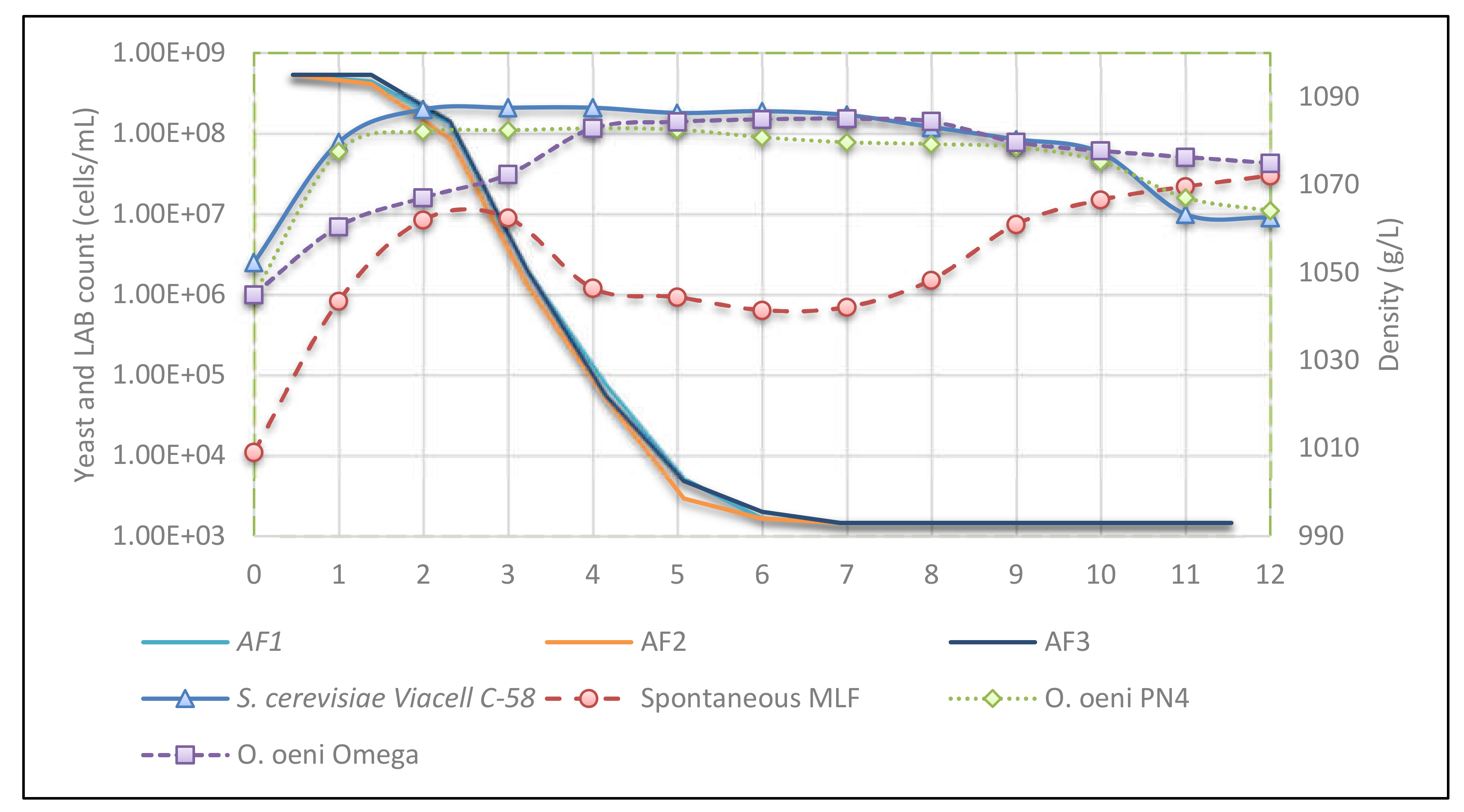
3.1. Evolution of Microbial Populations and Fermentation Kinetics
3.2. Malolactic Fermentation
3.3. Determination of Physic-Chemical Parameters of Fermentations and Color
3.4. Volatile Compound Analysis
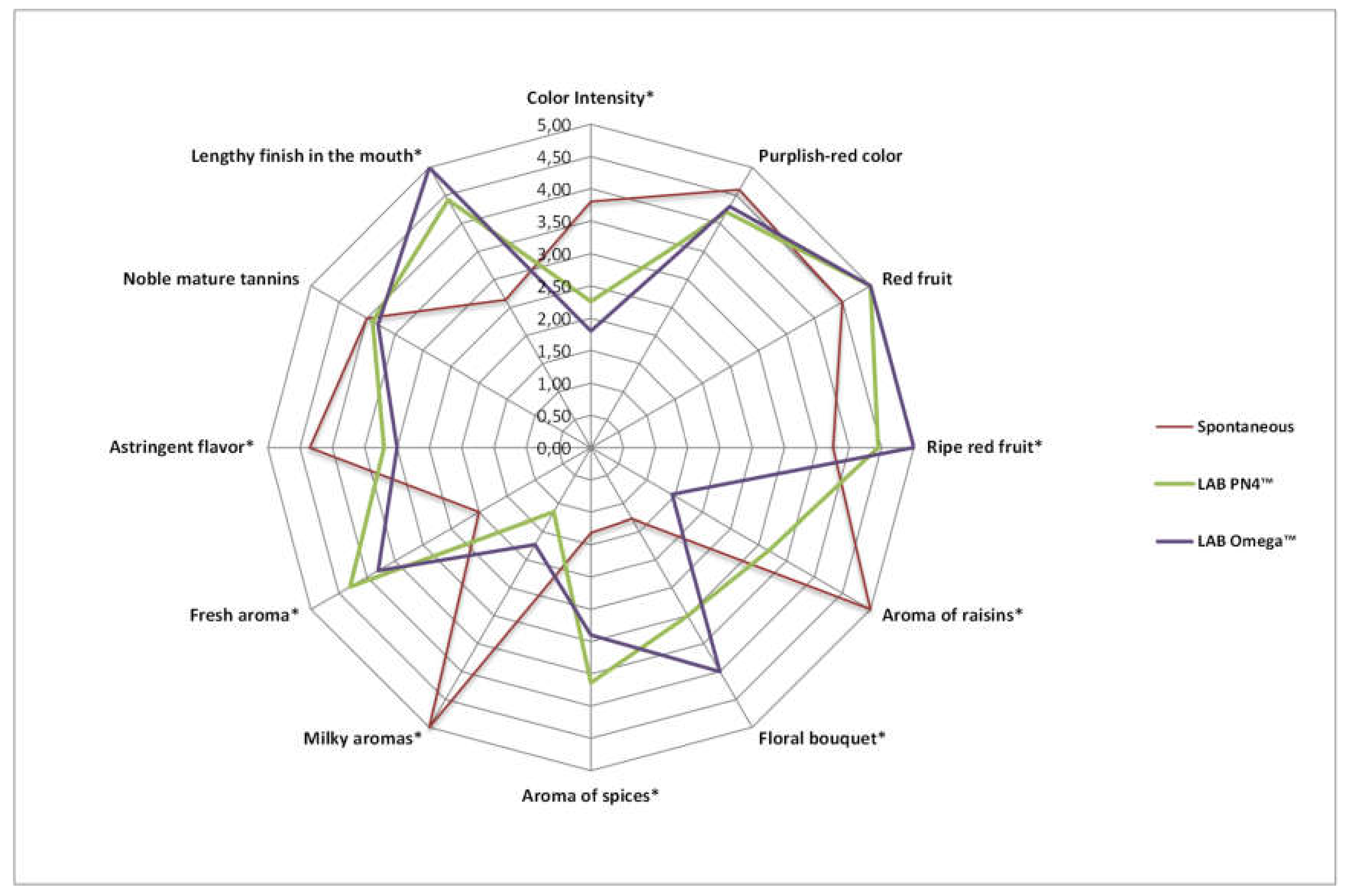
3.5. Sensory Analysis
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Davis, C.R.; Wibowo, D.; Eschenbruch, R.; Lee, T.H.; Fleet, G.H. Practical implications of malolactic fermentation: A review. Am. J. Enol. Viticult. 1985, 36, 290–301. [Google Scholar]
- Versari, A.; Patrizi, C.; Parpinello, G.P.; Mattioli, A.U.; Pasini, L.; Meglioli, M.; Longhini, G. Effect of co-inoculation with yeast and bacteria on chemical and sensory characteristics of commercial Cabernet Franc red wine from Switzerland. J. Chem. Technol. Biot. 2016. [Google Scholar] [CrossRef]
- Davis, C.R.; Wibowo, D.; Fleet, G.H.; Lee, T.H. Properties of wine lactic acid bacteria: Their potential enological significance. Am. Soc. Enol. Viticult. 1988, 39, 137–142. [Google Scholar]
- Styger, G.; Prior, B.; Bauer, F.F. Wine flavor and aroma. J. Ind. Microbiol. Biot. 2011. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Collins, T.S.; Masarweh, C.; Allen, G.; Heymann, H.; Ebeler, S.E.; Mills, D.A. Associations among wine grape microbiome, metabolome, and fermentation behavior suggest microbial contribution to regional wine characteristics. mBio 2016, 7, e00631-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belda, I.; Ruiz, J.; Esteban-Fernández, A.; Navascués, E.; Marquina, D.; Santos, A.; Moreno-Arribas, M.V. Microbial contribution to Wine aroma and its intended use for Wine quality improvement. Molecules 2017, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartowsky, E.J.; Borneman, A.R. Genomic variations of Oenococcus oeni strains and the potential to impact on malolactic fermentation and aroma compounds in wine. Appl. Microbiol. Biot. 2011, 92, 441–447. [Google Scholar] [CrossRef]
- Pozo-Bayón, M.; G.-Alegría, E.; Polo, M.C.; Tenorio, C.; Martín-Álvarez, P.J.; Calvo De La Banda, M.T.; Ruiz-Larrea, F.; Moreno-Arribas, M.V. Wine volatile and amino acid composition after malolactic fermentation: Effect of Oenococcus oeni and Lactobacillus plantarum starter cultures. J. Agr. Food Chem. 2005, 53, 8729–8735. [Google Scholar]
- Cappello, M.S.; Zapparoli, G.; Logrieco, A.; Bartowsky, E.J. Linking wine lactic acid bacteria diversity with wine aroma and flavour. Int. J. Food Microbiol. 2017. [Google Scholar] [CrossRef]
- De Revel, G.; Martin, N.; Pripis-Nicolau, L.; Lonvaud-Funel, A.; Bertrand, A. Contribution to the knowledge of malolactic fermentation influence on wine aroma. J. Agr. Food Chem. 1999, 47, 4003–4008. [Google Scholar] [CrossRef]
- Malherbe, S.; Menichelli, E.; du Toit, M.; Tredoux, A.; Muller, N.; Næs, T.; Nieuwoudt, H. The relationships between consumer liking, sensory and chemical attributes of Vitis vinifera L. cv. Pinotage wines elaborated with different Oenococcus oeni starter cultures. J. Sci. Food Agr. 2013. [Google Scholar] [CrossRef] [PubMed]
- Liu, S. Malolactic fermentation in wine—beyond deacidification. J. Appl. Microbiol. 2002, 92, 589–601. [Google Scholar] [CrossRef]
- Guo, Y.-Y.; Yang, Y.-P.; Peng, Q.; & Han, Y. Biogenic amines in wine: A review. Int. J. Food Sci. Tech. 2015, 50, 1523–1532. [Google Scholar] [CrossRef]
- Inês, A.; Falco, V. Lactic Acid Bacteria Contribution to Wine Quality and Safety. In Generation of Aromas and Flavours; Vilela, A., Ed.; InTechOpen: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.; Fleet, G.H. The degradation of malic acid by high-density cell suspensions of Leuconostoc oenos. J. Appl. Bacteriol. 1994. [Google Scholar] [CrossRef]
- Izquierdo-Cañas, P.M.I.; Pérez-Martín, F.; Romero, E.G.; Prieto, S.S.; de los Herreros, M.L.P. Influence of inoculation time of an autochthonous selected malolactic bacterium on volatile and sensory profile of Tempranillo and Merlot wines. Int. J. Food Microbiol. 2012. [Google Scholar] [CrossRef]
- Fugelsang, K.C.; Edwards, C.G. Wine microbiology. Practical Applications and Procedures, in Lactic Acid Bacteria, 2nd ed.; Springer: New York, NY, USA, 2007; pp. 29–41. [Google Scholar]
- Abrahamse, C.E.; Bartowsky, E.J. Timing of malolactic fermentation inoculation in Shiraz grape must and wine: Influence on chemical composition. W J. Microb Biot. 1007. [Google Scholar] [CrossRef] [PubMed]
- OIV. Compendium of international methods of analysis–OIV Chromatic Characteristics Method OIV-MA-AS2-07B: R2009; OIV: Paris, France, 2009; Available online: http://www.oiv.int/oiv/files/6-Domainesscientifiques/6-4Methodesdanalyses/6-4-1/EN/OIV-MA-AS2-07B.pdf.
- OIV. Compendium of international methods of wine and must analysis; International Organization of Vine and Wine: Paris, France.
- Ayala, F.; Echávarri, J.F.; Negueruela, A.I. A new simplified method for measuring the color of wines. I. Red and Rosé wines. Am. J. Enol. Viticult. 1997, 48, 357–363. [Google Scholar]
- Ribéreau-Gayon, P.; Stonestreet, E. Le dosage des anthocyanes dans les vins rouges. Bull. Soc. Chim. 1965, 9, 2649–2652. [Google Scholar]
- Sarneckis, C.J.; Dambergs, R.G.; Jones, P.; Mercurio, M.; Herderich, M.J.; Smith, P.A. Quantification of condensed tannins by precipitation with methyl-cellulose: Development and validation of an optimized tool for grape and wine analysis. Aust. J. Grape Wine Res. 2006, 12, 39–42. [Google Scholar] [CrossRef]
- Ibarz, M.; Ferreira, V.; Hernández-Orte, P.; Loscos, N.; Cacho, J. Optimization and evaluation of a procedure for the gas chromatographic-mass spectrometric analysis of the aromas generated by fast acid hydrolysis of flavors precursors extracted from grapes. J. Chromatogr. A. 2006, 1116, 217–229. [Google Scholar] [CrossRef]
- ISO Standard 11035. In Sensory analysis. Identification and selection of descriptors for stablishing a sensory profile; ISO: Geneva, Switzerland, 1994.
- Lucio, O.; Pardo, I.; Krieger-Weber, S.; Heras, J.M.; Ferrer, S. Selection of Lactobacillus strains to induce biological acidification in low acidity wines. LWT Food Sci Tech. 2016. [Google Scholar] [CrossRef]
- Pardo, I.; Ferrer, S. Yeast-Bacteria Co-inoculation. Red Wine Technol. 2019. [Google Scholar] [CrossRef]
- Lasik, M. The application of malolactic fermentation process to create good-quality grape wine produced in cool-climate countries: A review. Eur. Food Res. Technol. 2013. [Google Scholar] [CrossRef] [Green Version]
- Tristezza, M.; di Feo, L.; Tufariello, M.; Grieco, F.; Capozzi, V.; Spano, G.; Mita, G.; Grieco, F. Simultaneous inoculation of yeasts and lactic acid bacteria: Effects on fermentation dynamics and chemical composition of Negroamaro wine. LWT Food Sci. Tech. 2016. [Google Scholar] [CrossRef]
- Jussier, D.; Morneau, A.D.; De Orduña, R.M. Effect of simultaneous inoculation with yeast and bacteria on fermentation kinetics and key wine parameters of cool-climate Chardonnay. Appl. Environ. Microb. 2006. [Google Scholar] [CrossRef] [Green Version]
- Rosi, I.; Fia, G.; Canuti, V. Influence of different pH values and inoculation time on the growth and malolactic activity of a strain of Oenococcus oeni. Aust. J. Grape. Wine Res. 2003. [Google Scholar] [CrossRef]
- Massera, A.; Soria, A.; Catania, C.; Krieger, S.; Combina, M. Simultaneous inoculation of Malbec (Vitis Vinifera) musts with yeast and bacteria: Effects on fermentation performance, sensory and sanitary attributes of wines. Food Technol. Biotech. 2009. [Google Scholar]
- Zapparoli, G.; Tosi, E.; Azzolini, M.; Vagnoli, P.; Krieger, S. Bacterial inoculation strategies for the achievement of Malolactic fermentation in high-alcohol wines. S. Afr J. Enol. Vitic. 2009. [Google Scholar] [CrossRef]
- Antalick, G.; Perello, M.C.; De Revel, G. Co-inoculation with yeast and LAB under winery conditions: Modification of the aromatic profile of merlot wines. S. Afr. J. Enol. Vitic. 2013. [Google Scholar] [CrossRef] [Green Version]
- Alexandre, H.; Costello, P.J.; Remize, F.; Guzzo, J.; Guilloux-Benatier, M. Saccharomyces cerevisiae-Oenococcus oeni interactions in wine: Current knowledge and perspectives. Int. J. Food Microbiol. 2004. [Google Scholar] [CrossRef]
- Ribéreau-Gayon, P.; Glories, Y.; Maujean, A.; Dubourdieu, D. Handbook of Enology, The Chemistry of Wine: Stabilization and Treatments: Second Edition, 2nd ed.; Wiley: Hoboken, NJ, USA, 2006. [Google Scholar] [CrossRef]
- Tristezza, M.; Fantastico, L.; Vetrano, C.; Bleve, G.; Corallo, D.; Mita, G.; Grieco, F. Molecular and technological characterization of Saccharomyces cerevisiae strains isolated from natural fermentation of susumaniello grape must in Apulia, Southern Italy. Int. J. Microbiol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo-Cañas, P.M.; Romero, E.G.; Pérez-Martín, F.; Seseña, S.; Palop, M.L. Sequential inoculation versus co-inoculation in Cabernet Franc wine fermentation. Food Sci. Technol. Int. 2015. [Google Scholar] [CrossRef]
- Knoll, C.; Fritsch, S.; Schnell, S.; Grossmann, M.; Krieger-Weber, S.; du Toit, M.; Rauhut, D. Impact of different malolactic fermentation inoculation scenarios on Riesling wine aroma. World J. Microb. Biot. 2012. [Google Scholar] [CrossRef] [PubMed]
- Garofalo, C.; El Khoury, M.; Lucas, P.; Bely, M.; Russo, P.; Spano, G.; Capozzi, V. Autochthonous starter cultures and indigenous grape variety for regional wine production. J. Appl. Microbiol. 2015. [Google Scholar] [CrossRef]
- Du Plessis, H.; Du Toit, M.; Nieuwoudt, H.; Van Der Rijst, M.; Kidd, M.; Jolly, N. Effect of Saccharomyces, non-Saccharomyces yeasts and malolactic fermentation strategies on fermentation kinetics and flavor of Shiraz wines. Fermentation 2017, 64. [Google Scholar] [CrossRef] [Green Version]
- Burns, T.R.; Osborne, J.P. Impact of malolactic fermentation on the color and color stability of pinot noir and Merlot wine. Am. J. Enol. Viticult. 2013. [Google Scholar] [CrossRef]
- Fang, Y.; Qian, M. Aroma compounds in Oregon Pinot Noir wine determined by aroma extract dilution analysis (AEDA). Flavour Frag. J. 2005. [Google Scholar] [CrossRef]
- Bartowsky, E.J.; Henschke, P.A. The “buttery” attribute of wine - Diacetyl - Desirability, spoilage and beyond. Int J. Food Microbiol. 2004. [Google Scholar] [CrossRef]
- Gerbaux, V.; Briffox, C.; Dumont, A.; Krieger, S. Infuence of inoculation with Malolactic bacteria on volatile phenols in wines. Am. J. Enol. Viticult. 2009, 60, 233–235. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Spontaneous MLF | PN4TM | OmegaTM | |
|---|---|---|---|
| Alcoholic strength (% v/v) | 13.34 ± 0.66 a | 13.45 ± 0.15 a | 13.52 ± 0.23 a |
| Total acidity (g/L) | 3.49 ± 0.03 a | 3.62 ± 0.18 a | 3.63 ± 0.18 a |
| pH | 4.07 ± 0.08 a | 4.04 ± 0.07 a | 4.01 ± 0.03 a |
| Volatile acidity (g/L) | 0.35 ± 0.01 b | 0.28 ± 0.03 a | 0.26 ± 0.00 a |
| L-malic acid (g/L) | 0.07 ± 0.04 a | 0.05 ± 0.01 a | 0.08 ± 0.02 a |
| L-lactic acid (g/L) | 1.27 ± 0.00 c | 1.22 ± 0.02 b | 1.12 ± 0.06 a |
| Citric acid (g/L) | 0.07 ± 0.03 a | 0.20 ± 0.04 b | 0.19 ± 0.04 b |
| Glycerol (g/L) | 9.69 ± 0.93 a | 9.91 ± 0.26 a | 9.85 ± 0.28 a |
| L* | 17.13 ± 3.29 a | 15.34 ± 0.36 a | 19.26 ± 2.23 a |
| a* | 43.39 ± 1.31 a | 43.46 ± 0.68 a | 44.38 ± 0.09 a |
| b* | 22.13 ± 0.84 a | 21.28 ± 0.18 a | 20.99 ± 0.82 a |
| Color intensity | 6.44 ± 0.83 a | 6.79 ± 0.32 a | 5.89 ± 0.55 a |
| Color tone | 0.83 ± 0.00 b | 0.76 ± 0.04 a | 0.79 ± 0.04 ab |
| Total anthocyanins (mg/L) | 446.00 ± 39.60 a | 432.00 ± 33.94 a | 426.50 ± 27.58 a |
| Catechins (mg/L) | 694.05 ± 67.39 a | 730.10 ± 107.48 a | 614.60 ± 55.58 a |
| Tannins (mg/L) | 0.61 ± 0.12 a | 0.64 ± 0.15 a | 0.51 ± 0.30 a |
| Folin–Ciocalteu | 33.00 ± 4.24 a | 34.50 ± 3.54 a | 31.00 ± 1.41 a |
| Spontaneous MLF | PN4TM | OmegaTM | |
|---|---|---|---|
| Ethyl acetate (mg/L) | 80.00 ± 15.89 a | 95.18 ± 0.13 b | 108.65 ± 2.93 c |
| Isoamyl acetate (mg/L) | 5.85 ± 0.76 a | 6.13 ± 0.10 a | 8.85 ± 0.89 b |
| 2-Phenylethyl acetate (μg/L) | 182.61 ± 7.24 b | 141.89 ± 7.89 a | 170.63 ± 31.80 b |
| Hexyl acetate (μg/L) | 13.54 ± 1.77 a | 13.75 ± 0.84 a | 21.21 ± 7.18 b |
| Ethyl lactate (mg/L) | 145.86 ± 19.96 b | 137.78 ± 6.11 b | 117.54 ± 1.24 a |
| Ethyl butyrate (μg/L) | 632.50 ± 106.77 a | 687.50 ± 30.41 a | 896.00 ± 94.75 b |
| Ethyl hexanoate (μg/L) | 739.82 ± 26.96 a | 790.88 ± 57.99 ab | 866.78 ± 85.64 b |
| Ethyl octanoate (μg/L) | 716.64 ± 49.17 a | 677.01 ± 90.65 a | 785.66 ± 85.41 a |
| Ethyl decanoate (μg/L) | 97.26 ± 8.89 a | 90.29 ± 14.61 a | 100.97 ± 10.01 a |
| 1-propanol (mg/L) | 62.83 ± 8.25 a | 54.74 ± 5.25 a | 54.55 ± 0.51 a |
| Isobutanol (mg/L) | 30.48 ± 3.71 a | 28.96 ± 0.51 a | 30.65 ± 1.20 a |
| 1-butanol (mg/L) | 1.46 ± 0.30 a | 1.65 ± 0.12 a | 1.46 ± 0.23 a |
| Isoamylic alcohols (mg/L) | 342.50 ± 36.63 a | 332.19 ± 1.25 a | 327.92 ± 10.87 a |
| 1-octanol (μg/L) | 13.82 ± 2.45 a | 16.14 ± 2.27 a | 17.33 ± 1.77 a |
| 3-methyl-1-pentanol (μg/L) | 167.16 ± 42.26 a | 212.66 ± 6.96 c | 181.06 ± 1.16 ab |
| 3-ethoxy-1-propanol (μg/L) | 53.47 ± 13.10 a | 48.15 ± 3.02 a | 45.46 ± 2.95 a |
| 1-hexanol (mg/L) | 3.24 ± 0.12 a | 3.46 ± 0.16 b | 3.53 ± 0.08 b |
| t-3-hexenol (μg/L) | 74.70 ± 0.47 a | 75.24 ± 9.86 a | 74.74 ± 0.86 a |
| c-3-hexen-1-ol (μg/L) | 513.62 ± 88.94 | 516.16 ± 21.29 | 560.66 ± 4.26 |
| 2-phenylethanol (mg/L) | 23.68 ± 2.79 a | 26.07 ± 1.58 a | 22.54 ± 1.49 a |
| Isobutiric acid (μg/L) | 460.53 ± 49.70 a | 428.63 ± 51.80 a | 426.93 ± 10.52 a |
| Butyric acid (μg/L) | 570.86 ± 1.69 b | 436.89 ± 2.96 a | 513.28 ± 91.90 b |
| Isovaleric acid (mg/L) | 1.43 ± 0.05 c | 1.31 ± 0.08 b | 1.22 ± 0.06 a |
| Valeric acid (μg/L) | 13.68 ± 3.89 b | 7.71 ± 2.71 a | 6.68 ± 4.72 a |
| Hexanoic acid (mg/L) | 8.29 ± 0.46 b | 7.28 ± 0.57 a | 7.80 ± 0.34 ab |
| Octanoic acid (mg/L) | 7.25 ± 1.65 b | 5.02 ± 0.08 a | 5.41 ± 0.16 a |
| Decanoic acid (mg/L) | 3.78 ± 0.94 b | 2.08 ± 0.10 a | 1.92 ± 0.17 a |
| γ-nonalactone (μg/L) | 6.35 ± 0.53 a | 5.39 ± 0.56 a | 6.14 ± 0.55 a |
| δ-dodecalactone (μg/L) | 9.58 ± 0.67 a | 6.89 ± 1.05 a | 7.56 ± 3.70 a |
| δ-ethoxycarbonyl-δ-butyrolactone (μg/L) | 321.46 ± 39.16 ab | 275.65 ± 26.25 a | 355.95 ± 31.85 b |
| Damascenone (μg/L) | 6.03 ± 0.81 a | 6.27 ± 0.23 a | 5.74 ± 0.97 a |
| 3-oxo-α-ionol (μg/L) | 55.82 ± 0.60 a | 49.64 ± 7.01 a | 53.82 ± 4.76 a |
| Acetaldehyde (mg/L) | 9.28 ± 0.05 a | 10.87 ± 0.96 b | 14.00 ± 1.16 c |
| 2.3 Butanodione (mg/L) | 18.35 ± 1.67 c | 14.77 ± 1.61 b | 8.55 ± 0.48 a |
| 1-octen-3-ona (μg/L) | 14.25 ± 0.59 a | 14.02 ± 1.84 a | 14.19 ± 2.08 a |
| 3-methyl-thio-propanol (μg/L) | 277.33 ± 85.07 a | 311.44 ± 66.18 a | 211.11 ± 10.75 a |
| Furaneol (μg/L) | 12.32 ± 1.26 b | 5.94 ± 3.73 a | 10.39 ± 2.52 b |
| Guaiacol (μg/L) | 12.39 ± 1.65 a | 18.44 ± 0.37 b | 19.88 ± 5.18 b |
| Eugenol (μg/L) | 3.97 ± 0.36 b | 2.52 ± 0.79 a | 2.71 ± 0.13 a |
| Phenol (μg/L) | 8.33 ± 0.98 a | 9.41 ± 0.66 a | 10.24 ± 3.49 a |
| 4-metil-2, 6-ditercbutil-fenol (μg/L) | 95.91 ± 29.03 b | 66.28 ± 8.35 a | 63.06 ± 0.07 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Izquierdo-Cañas, P.M.; Ríos-Carrasco, M.; García-Romero, E.; Mena-Morales, A.; Heras-Manso, J.M.; Cordero-Bueso, G. Co-Existence of Inoculated Yeast and Lactic Acid Bacteria and Their Impact on the Aroma Profile and Sensory Traits of Tempranillo Red Wine. Fermentation 2020, 6, 17. https://doi.org/10.3390/fermentation6010017
Izquierdo-Cañas PM, Ríos-Carrasco M, García-Romero E, Mena-Morales A, Heras-Manso JM, Cordero-Bueso G. Co-Existence of Inoculated Yeast and Lactic Acid Bacteria and Their Impact on the Aroma Profile and Sensory Traits of Tempranillo Red Wine. Fermentation. 2020; 6(1):17. https://doi.org/10.3390/fermentation6010017
Chicago/Turabian StyleIzquierdo-Cañas, Pedro Miguel, María Ríos-Carrasco, Esteban García-Romero, Adela Mena-Morales, José María Heras-Manso, and Gustavo Cordero-Bueso. 2020. "Co-Existence of Inoculated Yeast and Lactic Acid Bacteria and Their Impact on the Aroma Profile and Sensory Traits of Tempranillo Red Wine" Fermentation 6, no. 1: 17. https://doi.org/10.3390/fermentation6010017
APA StyleIzquierdo-Cañas, P. M., Ríos-Carrasco, M., García-Romero, E., Mena-Morales, A., Heras-Manso, J. M., & Cordero-Bueso, G. (2020). Co-Existence of Inoculated Yeast and Lactic Acid Bacteria and Their Impact on the Aroma Profile and Sensory Traits of Tempranillo Red Wine. Fermentation, 6(1), 17. https://doi.org/10.3390/fermentation6010017