Non-Saccharomyces Yeasts and Organic Wines Fermentation: Implications on Human Health


Abstract
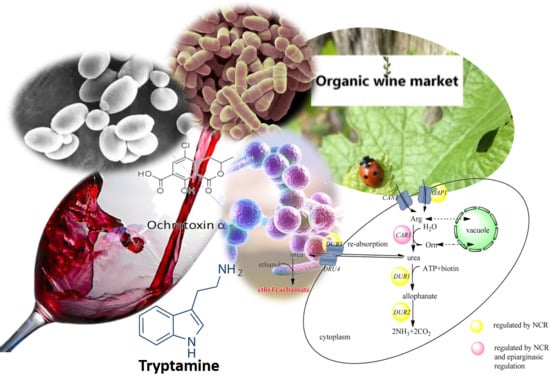
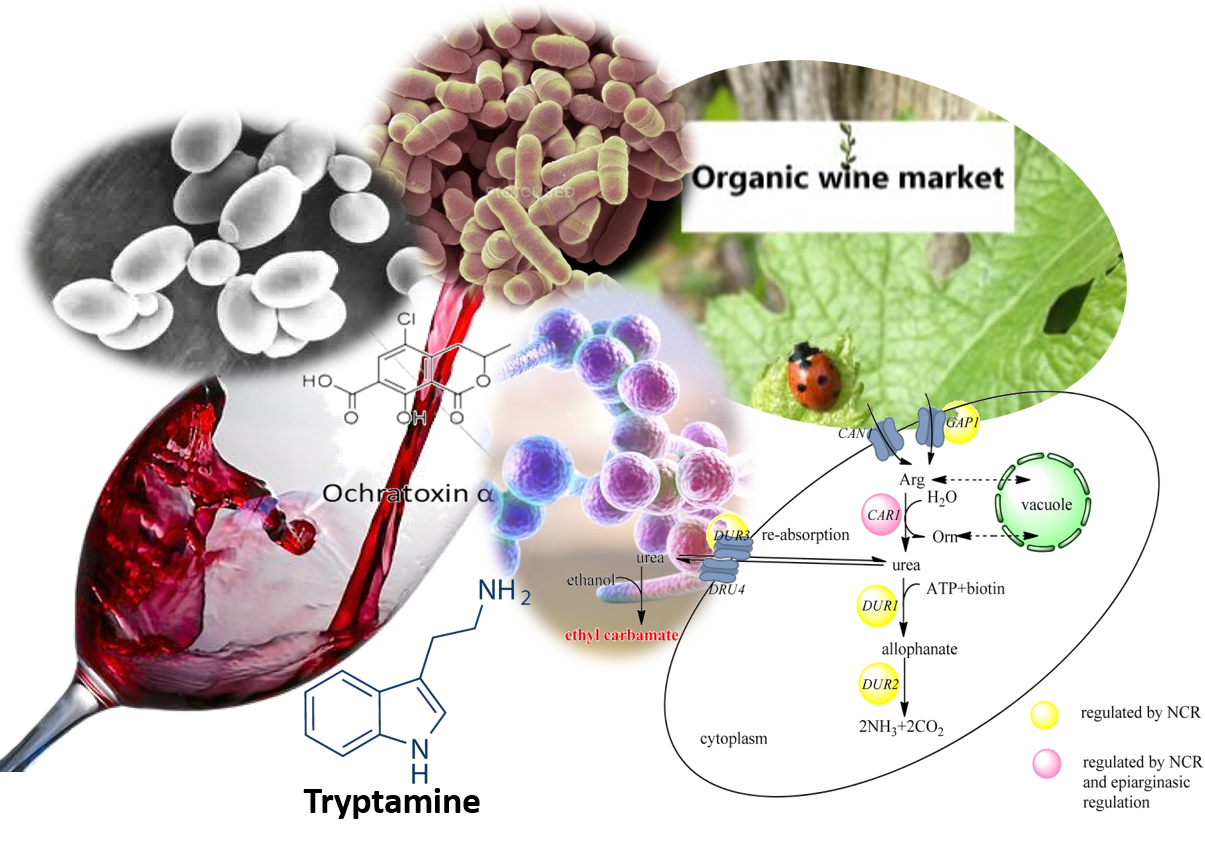
:
1. General Introduction
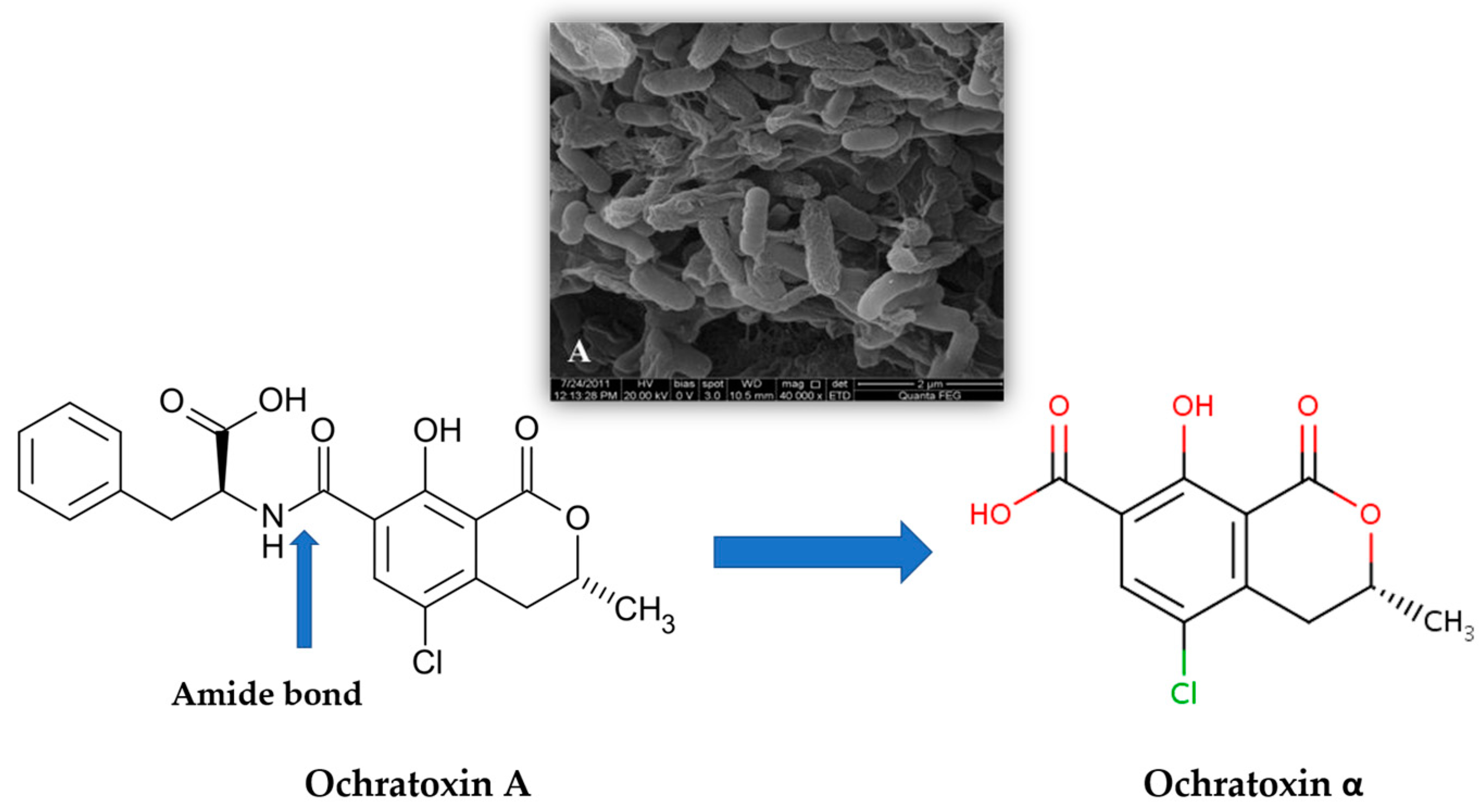
2. Wine Contamination by Ochratoxin A (OTA) and Other Mycotoxins
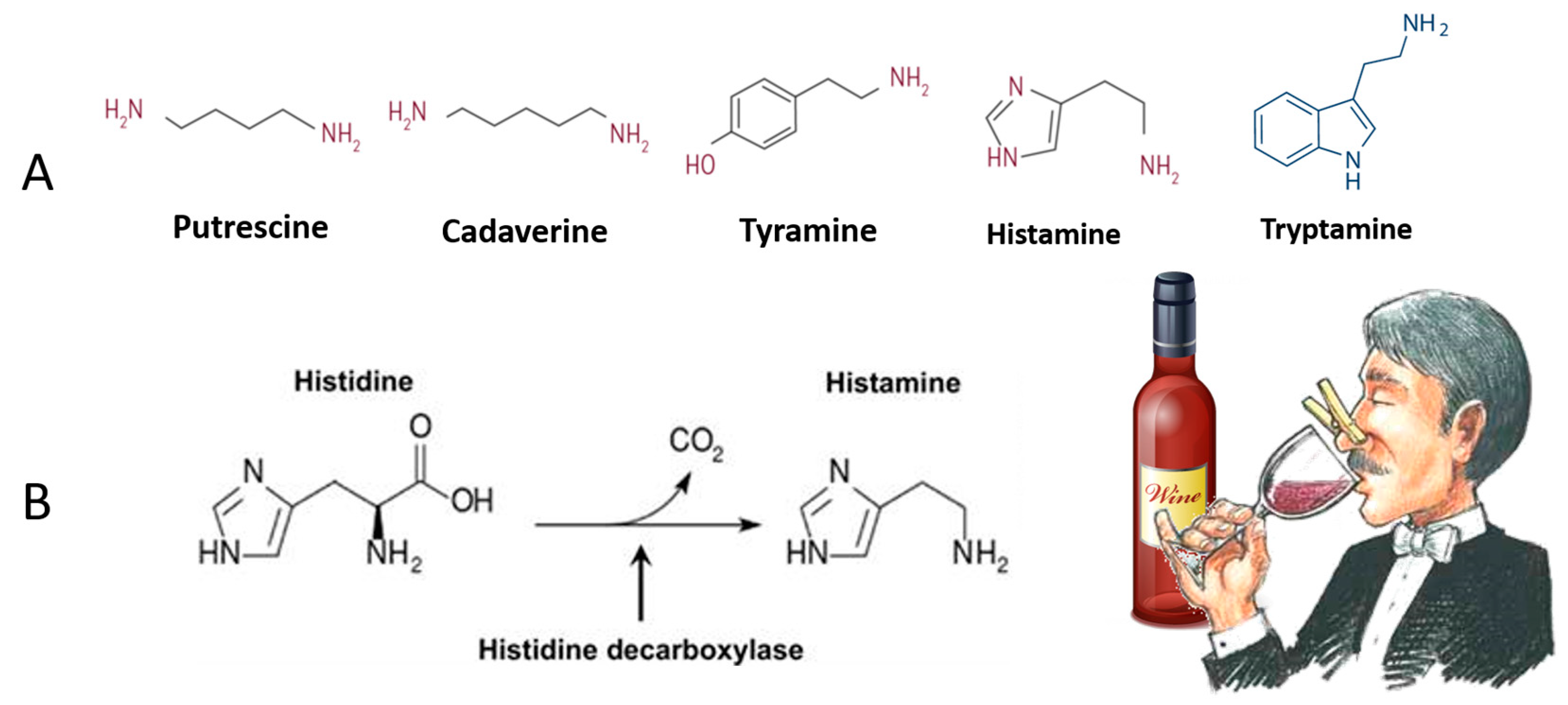
3. Organic Wines Contamination with Biogenic Amines
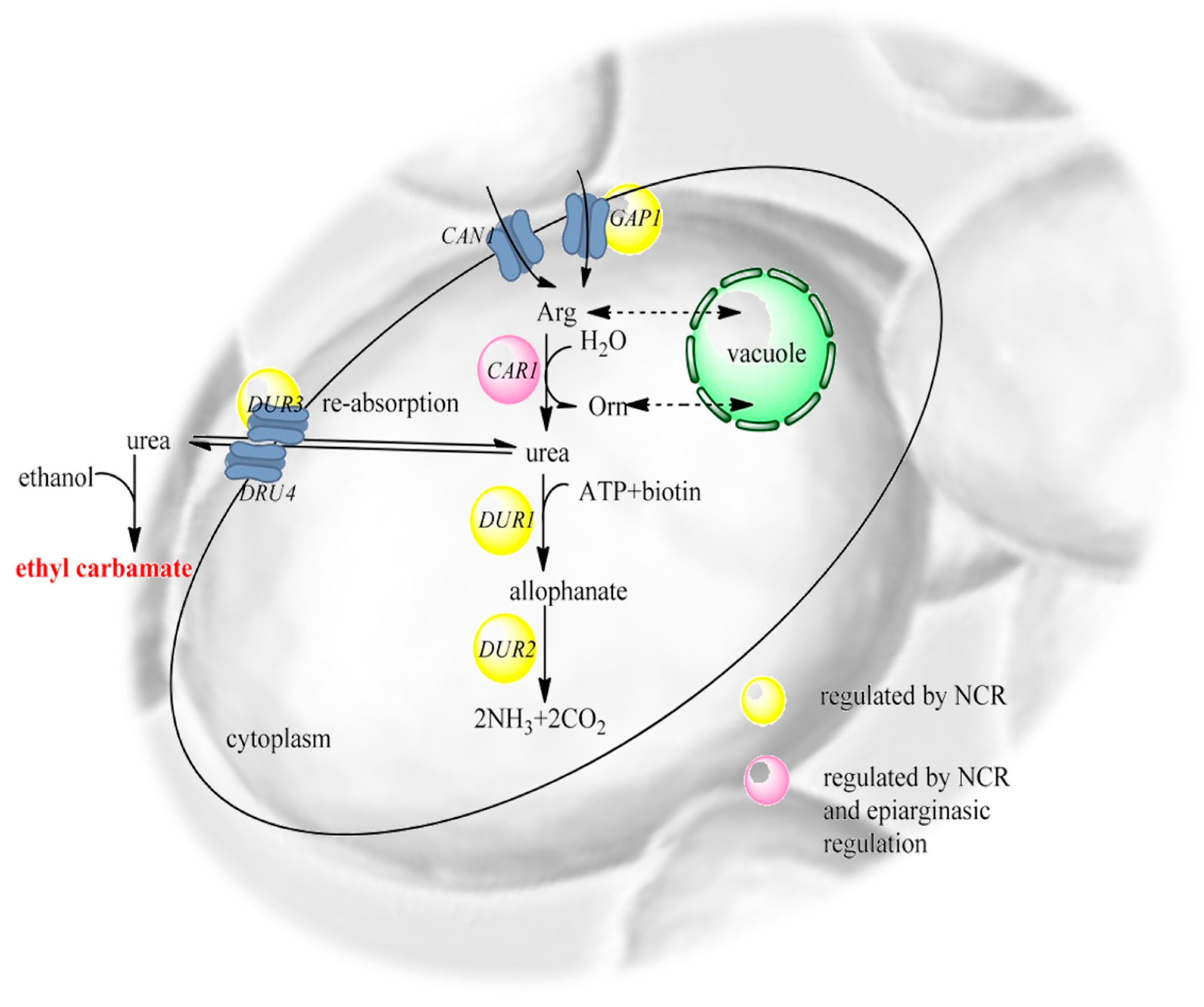
4. Wines and Ethyl Carbamate Contamination
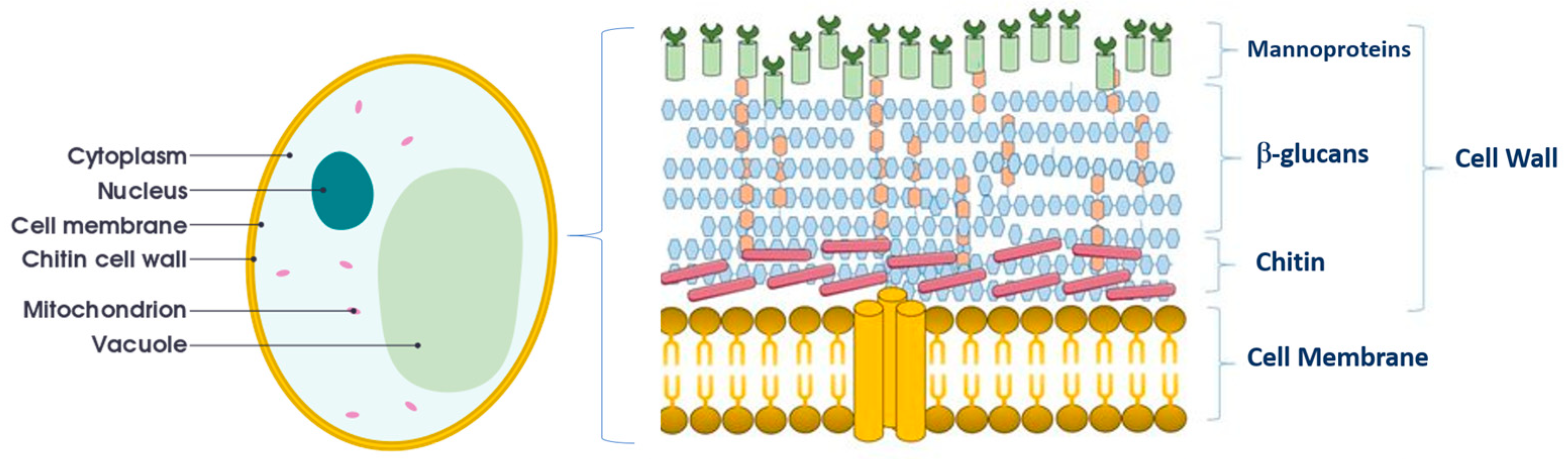
5. Other Benefits of the Use of Non-Saccharomyces in Organic Winemaking
6. Final Remarks
Funding
Acknowledgments
Conflicts of Interest
References
- IFOAM. Basic Standards for Organic Production and Processing. Bonn-Germany. 2014. Available online: http://www.ifoam.orghttps://www.ifoam.bio/sites/default/files/ifoam_norms_july_2014_t.pdf (accessed on 15 April 2020).
- Trioli, G.; Hofmann, U. ORWINE: Code of good organic viticulture and winemaking. In ECOVIN-Federal Association of Organic Wine-Producer; Ecovin: Oppenheim, Germany, 2009. [Google Scholar]
- IFAOM. EU Rules for Organic Wine Production: Background, Evaluation, and Further Sector Development. 2013. Available online: https://orgprints.org/29867/1/ifoameu_reg_wine_dossier_201307.pdf (accessed on 17 April 2020).
- Commission Implementing Regulation (EU) No 203/2012 of 8 March 2012 Amending Regulation (EC) No 889/2008 Laying Down Detailed Rules for the Implementation of Council Regulation (EC) No 834/2007, as Regards Detailed Rules on Organic Wine. OJ L 71. 9 March 2012, pp. 42–47. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex%3A32012R0203 (accessed on 10 May 2020).
- Schäufele, I.; Hamm, U. Consumers’ perceptions, preferences, and willingness-to-pay for wine with sustainability characteristics: A review. J. Clean. Prod. 2017, 147, 379–394. [Google Scholar] [CrossRef]
- Cravero, M.C. Organic, and biodynamic wines quality and characteristics: A review. Food Chem. 2019, 295, 334–340. [Google Scholar] [CrossRef] [PubMed]
- DIVA. Overview of the Organic Wine Market. 2007. Available online: https://divawine.com/overview-organic-market/ (accessed on 7 May 2020).
- Tofalo, R.; Schirone, M.; Telera, G.C.; Manetta, A.C.; Corsetti, A.; Suzzi, G. Influence of organic viticulture on non-Saccharomyces wine yeast populations. Ann. Microbiol. 2011, 61, 57–66. [Google Scholar] [CrossRef]
- Russo, P.; Capozzi, V.; Spano, G.; Corbo, M.R.; Sinigaglia, M.; Antonio, B. Metabolites of Microbial Origin with an Impact on Health: Ochratoxin A and Biogenic Amines. Front. Microbiol. 2016, 7, 482. [Google Scholar] [CrossRef]
- Ciani, M.; Comitini, F. Non-Saccharomyces wine yeasts have a promising role in biotechnological approaches to winemaking. Ann. Microbiol. 2011, 61, 25–32. [Google Scholar] [CrossRef]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [Green Version]
- Maurizio, C.; Francesca, C.; Ilaria, M.; Paola, D. Controlled mixed culture fermentation: A new perspective on the use of non-Saccharomyces yeasts in winemaking. FEMS Yeast Res. 2010, 10, 123–133. [Google Scholar] [CrossRef] [Green Version]
- Vilela, A. The Importance of Yeasts on Fermentation Quality and Human Health-Promoting Compounds. Fermentation 2019, 5, 46. [Google Scholar] [CrossRef] [Green Version]
- Vilela, A. Modulating Wine Pleasantness Throughout Wine-Yeast Co-Inoculation or Sequential Inoculation. Fermentation 2020, 6, 22. [Google Scholar] [CrossRef] [Green Version]
- Walker, R. Risk assessment of ochratoxin: Current views of the European Scientific Committee on Food, the JECFA and the Codex Committee on Food Additives and Contaminants. Adv. Exp. Med. Biol. 2002, 504, 249–255. [Google Scholar] [CrossRef]
- Ji, C.; Fan, Y.; Zhao, L. Review on biological degradation of mycotoxins. Anim. Nutr. 2016, 2, 127–133. [Google Scholar] [CrossRef]
- Barreira, M.J.; Alvito, P.C.; Almeida, C.M. Occurrence of patulin in apple-based foods in Portugal. Food Chem. 2010, 121, 653–658. [Google Scholar] [CrossRef]
- Peraica, M.; Radic, B.; Lucicć, P.; Pavlovic, M. Toxic effects of mycotoxins in humans. Int. J. Public Health 1999, 77, 754–766. [Google Scholar]
- Ferenczi, S.; Cserháti, M.; Krifaton, C.; Szoboszlay, S.; Kukolya, J.; Szőke, Z.; Kovács, K.J. A New Ochratoxin A Biodegradation Strategy Using Cupriavidus basilensis Őr16 Strain. PLoS ONE 2014, 9, e109817. [Google Scholar] [CrossRef] [PubMed]
- Spano, G.; Russo, P.; Lonvaud-Funel, A.; Lucas, P.; Alexandre, H.; Grandvalet, C.; Rattray, F. Biogenic amines in fermented foods. Eur. J. Clin. Nutr. 2010, 64, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Petruzzi, L.; Sinigaglia, M.; Corbo, M.R.; Campaniello, D.; Speranza, B.; Bevilacqua, A. Decontamination of Ochratoxin A by yeasts: Possible approaches and factor leading to toxin removal in wine. Appl. Microbiol. Biotechnol. 2014, 98, 6555–6567. [Google Scholar] [CrossRef]
- Battilani, P.; Giorni, P.; Bertuzzi, T.; Formenti, S.; Pietri, A. Black Aspergilli and Ochratoxin A in grapes in Italy. Int. J. Food Microbiol. 2006, 111, S53–S60. [Google Scholar] [CrossRef]
- Bellver Soto, J.; Fernández-Franzón, M.; Ruiz, M.J.; García, A.J. Presence of Ochratoxin A (OTA) mycotoxin in alcoholic drinks from southern European countries: Wine and beer. J. Agric. Food Chem. 2014, 62, 7643–7651. [Google Scholar] [CrossRef]
- IARC. Mycotoxins and Human Health (Chapter 6). 2020; pp. 87–104. Available online: file:///C:/Users/avimo/Downloads/IARC_SP158_Chapter%206.pdf (accessed on 17 April 2020).
- EFSA (European Food Safety Authority). Opinion of the scientific panel on contaminants in the food chain on a request. Commission related to Ochratoxin A in food. EFSA J. 2006, 365, 1–56. [Google Scholar]
- Quintela, S.; Villarán, M.C.; Armentia, I.L.; Elejalde, E. Ochratoxin A removal in wine: A review. Food Control 2013, 30, 439–445. [Google Scholar] [CrossRef]
- Pitout, M.J. The hydrolysis of ochratoxin A by some proteolytic enzymes. Biochem. Pharmacol. 1969, 18, 485–491. [Google Scholar] [CrossRef]
- Abrunhosa, L.; Inês, A.; Rodrigues, A.I.; Guimarães, A.; Pereira, V.L.; Paropt, P.; Venâncio, A. Biodegradation of Ochratoxin A by Pediococcus parvulus isolated from Douro wines. Int. J. Food Microbiol. 2014, 188, 45–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Bellis, P.; Tristezza, M.; Haidukowski, M.; Fanelli, F.; Sisto, A.; Mulè, G.; Grieco, F. Biodegradation of Ochratoxin A by Bacterial Strains Isolated from Vineyard Soils. Toxins (Basel) 2015, 7, 5079–5093. [Google Scholar] [CrossRef] [PubMed]
- Friman, H.; Schechter, A.; Ioffe, Y.; Nitzan, Y.; Cahan, R. Electricity formation in a microbial fuel cell. Microb. Biotechnol. 2013, 6, 425–434. [Google Scholar] [CrossRef]
- Caridi, A.; Cufari, A.; Lovino, R.; Palumbo, R.; Tedesco, I. Influence of yeast on polyphenol composition of wine. Food Technol. Biotechnol. 2004, 42, 37–40. [Google Scholar]
- Petruzzi, L.; Baiano, A.; De Gianni, A.; Sinigaglia, M.; Corbo, M.R.; Bevilacqua, A. Differential Adsorption of Ochratoxin A and Anthocyanins by Inactivated Yeasts and Yeast Cell Walls during Simulation of Wine Aging. Toxins (Basel) 2015, 7, 4350–4365. [Google Scholar] [CrossRef] [Green Version]
- Bevilacqua, A.; Petruzzi, L.; Corbo, M.R.; Baiano, A.; Garofalo, C.; Sinigaglia, M. Ochratoxin A released back into the medium by Saccharomyces cerevisiae as a function of the strain, washing medium, and fermentative conditions. J. Sci. Food Agric. 2014, 94, 3291–3295. [Google Scholar] [CrossRef]
- Mazauric, J.P.; Salmon, J.M. Interactions between yeast lees and wine polyphenols during simulation of wine aging: II. Analysis of desorbed polyphenol compounds from yeast lees. J. Agric. Food Chem. 2006, 54, 3876–3881. [Google Scholar] [CrossRef]
- Palacios, S.; Vasserot, Y.; Maujean, A. Evidence for sulfur volatile products adsorption by yeast lees. Am. J. Enol. Vitic. 1997, 48, 525–526. [Google Scholar]
- Alexandre, H.; Lubbers, S.; Charpentier, C. Interactions between toxic fatty acids for yeasts and colloids, cellulose and yeast ghost using the equilibrium dialysis method in a model wine system. Food Biotechnol. 1997, 11, 89–99. [Google Scholar] [CrossRef]
- Navarro, S.; Barba, A.; Oliva, J.; Navarro, G.; Pardo, F. Evolution of residual levels of six pesticides during elaboration of red wines. Effect of wine-making procedures in their disappearance. J. Agric. Food Chem. 1999, 47, 264–270. [Google Scholar] [CrossRef]
- Pradelles, R.; Chassagne, D.; Vichi, S.; Gougeon, R.; Alexandre, H. (−) Geosmin sorption by enological yeasts in model wine and FTIR spectroscopy characterization of the sorbent. Food Chem. 2010, 120, 531–538. [Google Scholar] [CrossRef]
- Palomero, F.; Ntanos, K.; Morata, A.; Benito, S.; Suárez-Lepe, J.A. Reduction of wine 4-ethylphenol concentration using lyophilized yeast as a bioadsorbent: Influence on anthocyanin content and chromatic variables. Eur. Food Res. Technol. 2011, 232, 971–977. [Google Scholar] [CrossRef]
- Anwar, M.I.; Muhammad, F.; Awais, M.M.; Akhtar, M. A review of β-glucans as a growth promoter and antibiotic alternative against enteric pathogens in poultry. World’s Poult. Sci. J. 2017, 73, 651–661. [Google Scholar] [CrossRef]
- Huwig, A.; Freimund, S.; Käppeli, O.; Dutler, H. Mycotoxin detoxication of animal feed by different adsorbents. Toxicol Lett. 2001, 122, 179–188. [Google Scholar] [CrossRef]
- Domizio, P.; Liu, Y.; Bisson, L.F.; Barile, D. Cell wall polysaccharides released during the alcoholic fermentation by Schizosaccharomyces pombe and S. japonicus: Quantification and characterization. Food Microbiol. 2017, 61, 136–149. [Google Scholar] [CrossRef] [Green Version]
- Marx, H.; Sauer, M.; Resina, D.; Vai, M.; Porro, D.; Valero, F.; Ferrer, P.; Mattanovich, D. Cloning, disruption and protein secretory phenotype of the GAS1 homologue of Pichia pastoris. FEMS Microbiol. Lett. 2006, 264, 40–47. [Google Scholar] [CrossRef] [Green Version]
- Jigami, Y.; Odani, T. Mannosylphosphate transfer to yeast mannan. Molecular and Cell Biology of Lipids. Biochim. Biophys. Acta 1999, 1426, 335–345. [Google Scholar] [CrossRef]
- Esti, M.; Benucci, I.; Liburdi, K.; Acciaro, G. Monitoring of ochratoxin A fate during alcoholic fermentation of wine must. Food Control. 2012, 27, 53–56. [Google Scholar] [CrossRef]
- Luo, Y.; Liu, X.; Yuan, L.; Li, J. Complicated interactions between bio-adsorbents and mycotoxins during mycotoxin adsorption: Current research and future prospects. Trends Food Sci. Technol. 2020, 96, 127–134. [Google Scholar] [CrossRef]
- Bejaoui, H.; Mathieu, F.; Taillandier, P.; Lebrihi, A. Ochratoxin A removal in synthetic and natural grape juices by selected oenological Saccharomyces strains. J. Appl. Microbiol. 2004, 97, 1038–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ringot, D.; Lerzy, B.; Bonhoure, J.P.; Auclair, E.; Oriol, E.; Larondelle, Y. Effect of temperature on in vitro ochratoxin A biosorption onto yeast cell wall derivatives. Proc. Biochem. 2005, 40, 3008–3016. [Google Scholar] [CrossRef]
- Caridi, A. New perspectives in safety and quality enhancement of wine through selection of yeasts based on the parietal adsorption activity. Int. J. Food Microbiol. 2007, 120, 167–172. [Google Scholar] [CrossRef]
- Piotrowska, M.; Masek, A. Saccharomyces Cerevisiae Cell Wall Components as Tools for Ochratoxin A Decontamination. Toxins 2015, 7, 1151–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farbo, M.G.; Urgeghe, P.P.; Fiori, S.; Marceddu, S.; Jaoua, S.; Migheli, Q. Adsorption of Ochratoxin A from grape juice by yeast cells immobilised in calcium alginate beads. Int. J. Food Microbiol. 2016, 217, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Pfliegler, W.P.; Pusztahelyi, T.; Pócsi, I. Mycotoxins-prevention and decontamination by yeasts. J. Basic Microbiol. 2015, 55, 805–818. [Google Scholar] [CrossRef]
- Cecchini, F.; Morassut, M.; Saiz, J.C.; Moruno, E.G. Anthocyanins enhance yeast’s adsorption of Ochratoxin A during the alcoholic fermentation. Eur. Food Res. Technol. 2019, 245, 309–314. [Google Scholar] [CrossRef]
- Luo, Y.; Wang, Z.L.; Yuan, Y.H.; Zhou, Z.K.; Yue, T.L. Patulin adsorption of a superior microorganism strain with low flavor-affection of kiwi fruit juice. World Mycotoxin J. 2016, 9, 195–203. [Google Scholar] [CrossRef]
- Nunez, Y.P.; Pueyo, E.; Carrascosa, A.V.; Martínez-Rodríguez, A.J. Effects of Aging and Heat Treatment on Whole Yeast Cells and Yeast Cell Walls and on Adsorption of Ochratoxin A in a Wine Model System. J. Food Prot. 2008, 71, 1496–1499. [Google Scholar] [CrossRef]
- Armando, M.R.; Pizzolitto, R.P.; Dogi, C.A.; Cristofolini, A.; Merkis, C.; Poloni, V.; Cavaglieri, L.R. Adsorption of ochratoxin A and zearalenone by potential probiotic Saccharomyces cerevisiae strains and its relation with cell wall thickness. J. Appl. Microbiol. 2012, 113, 256–264. [Google Scholar] [CrossRef]
- Luo, Y.; Wang, J.G.; Liu, B.; Wang, Z.L.; Yuan, Y.H.; Yue, T.L. Effect of yeast cell morphology, cell wall physical structure and chemical composition on patulin adsorption. PLoS ONE 2015, 21, e0136045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolken, W.A.M.; Lucas, P.M.; Lonvaud-Funel, A.; Lolkema, J.S. The mechanism of the tyrosine transporter TyrP supports a proton motive tyrosine decarboxylation pathway in Lactobacillus brevis. J. Bacteriol. 2006, 188, 2198–2206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smit, A.Y.; du Toit, W.J.; du Toit, M. Biogenic amines in wine: Understanding the headache. South Afr. J. Enol. Vitic. 2008, 29, 109–127. [Google Scholar] [CrossRef] [Green Version]
- Cappello, M.S.; Zapparoli, G.; Logrieco, A.; Bartowsky, E.J. Linking wine lactic acid bacteria diversity with wine aroma and flavour. Review article. Int. J. Food Microbiol. 2017, 243, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.-Y.; Yang, Y.-P.; Peng, Q.; Han, Y. Biogenic amines in wine: A review. Int. J. Food Sci. Technol. 2015, 50, 1523–1532. [Google Scholar] [CrossRef]
- Martuscelli, M.; Mastrocola, D. Biogenic Amines: A Claim for Wines, Biogenic Amines, Charalampos Proestos; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Beneduce, L.; Romano, A.; Capozzi, V.; Lucas, P.; Barnavon, L.; Bach, B.; Spano, G. Biogenic amines in regional wines. Ann. Microbiol. 2010, 60, 573–578. [Google Scholar] [CrossRef]
- Ancín-Azpilicueta, C.; González-Marco, A.; Jiménez-Moreno, N. Current knowledge about the presence of amines in wine. Crit. Rev. Food Sci. Nutr. 2008, 48, 257–275. [Google Scholar] [CrossRef]
- Tassoni, A.; Tango, N.; Ferri, M. Comparison of biogenic amine and polyphenol profiles of grape berries and wines obtained following conventional, organic and biodynamic agricultural and oenological practices. Food Chem. 2013, 139, 405–413. [Google Scholar] [CrossRef]
- Yildirim, H.K.; Üren, A.; Yücel, U. Evaluation of biogenic amines in organic and non-organic wines by HPLC OPA derivatization. Food Technol. Biotechnol. 2007, 45, 62–68. [Google Scholar]
- Garcia-Marino, M.; Trigueros, Á.; Escribano-Bailón, T. Influence of oenological practices on the formation of biogenic amines in quality red wines. J. Food Comp. Anal. 2010, 23, 455–462. [Google Scholar] [CrossRef] [Green Version]
- Tassoni, A.; Tango, N.; Ferri, M. Polyphenol and Biogenic Amine Profiles of Albana and Lambrusco Grape Berries and Wines Obtained Following Different Agricultural and Oenological Practices. Food Nutr. Sci. 2014, 5, 8–16. [Google Scholar] [CrossRef] [Green Version]
- Caruso, M.; Fiore, C.; Contursi, M.; Salzano, G.; Paparella, A.; Romano, P. Formation of biogenic amines as criteria for the selection of wine yeasts. World J. Microbiol. Biotechnol. 2002, 18, 159–163. [Google Scholar] [CrossRef]
- Benito, Á.; Calderón, F.; Palomero, F.; Benito, S. Quality and Composition of Airén Wines Fermented by Sequential Inoculation of Lachancea thermotolerans and Saccharomyces cerevisiae. Food Technol. Biotechnol. 2016, 54, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Medina, K.; Boido, E.; Fariña, L.; Gioia, O.; Gomez, M.E.; Barquet, M.; Gaggero, C.; Dellacassa, E.; Carrau, F. Increased flavour diversity of Chardonnay wines by spontaneous fermentation and co-fermentation with Hanseniaspora vineae. Food Chem. 2013, 141, 2513–2521. [Google Scholar] [CrossRef]
- Thibon, C.; Marullo, P.; Claisse, O.; Cullin, C.; Dubourdieu, D.; Tominaga, T. Nitrogen catabolic repression controls the release of volatile thiols by Saccharomyces cerevisiae during wine fermentation. Fems Yeast Res. 2008, 8, 1076–1086. [Google Scholar] [CrossRef] [Green Version]
- Thorgeirsson, U.P.; Dalgard, D.W.; Reeves, J.; Adamson, R.H. Tumor incidence in a chemical carcinogenesis study of nonhuman primates. Regul. Toxicol. Pharm. 1994, 19, 130–151. [Google Scholar] [CrossRef]
- Jiao, Z.; Dong, Y.; Chen, Q. Ethyl Carbamate in Fermented Beverages: Presence, Analytical Chemistry, Formation Mechanism, and Mitigation Proposals. Compr. Rev. Food Sci. Food Saf. 2014, 13, 611–626. [Google Scholar] [CrossRef]
- Salmon, A.G.; Zeise, L. Risks of Carcinogenesis from Urethane Exposure; CRC Press: Boca Raton, FL, USA, 1991; p. 115. [Google Scholar]
- Beland, F.A.; Benson, R.W.; Mellick, P.W.; Kovatch, R.M.; Roberts, D.W.; Fang, J.-L.; Doerge, D.R. Effect of ethanol on the tumorigenicity of urethane (ethyl carbamate) in B6C3F1 mice. Food Chem. Toxicol. 2005, 43, 1–19. [Google Scholar] [CrossRef]
- Dahabieh, M.; Husnik, J.; Van Vuuren, H. Functional enhancement of sake yeast strains to minimize the production of ethyl carbamate in sake wine. J. Appl. Microbiol. 2010, 109, 963–973. [Google Scholar] [CrossRef]
- Arena, M.; Saguir, F.; Manca de Nadra, M. Arginine, citrulline and ornithine metabolism by lactic acid bacteria from wine. Int. J. Food Microbiol. 1999, 52, 155–161. [Google Scholar] [CrossRef]
- Polychroniadou, E.; Kanellaki, M.; Iconomopoulou, M.; Koutinas, A.; Marchant, R.; Banat, I. Grape and apple wines volatile fermentation products and possible relation to spoilage. Bioresour. Technol. 2003, 87, 337–339. [Google Scholar] [CrossRef]
- Zhao, X.; Zou, H.; Fu, J.; Chen, J.; Zhou, J.; Du, G. Nitrogen regulation involved in the accumulation of urea in Saccharomyces cerevisiae. Yeast 2013, 30, 437–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fidaleo, M.; Esti, M.; Moresi, M. Assessment of urea degradation rate in model wine solutions by acid urease from Lactobacillus fermentum. J. Agric. Food Chem. 2006, 54, 6226–6235. [Google Scholar] [CrossRef] [PubMed]
- Benito, S. The Management of Compounds that Influence Human Health in Modern Winemaking from an HACCP Point of View. Fermentation 2019, 5, 33. [Google Scholar] [CrossRef] [Green Version]
- Andrich, L.; Esti, M.; Moresi, M. Urea removal in model wine solutions by immobilized acid urease in a stirred bioreactor. Chem. Eng. Trans. 2009, 17, 915–920. [Google Scholar] [CrossRef]
- Pflaum, T.; Hausler, T.; Baumung, C.; Ackermann, S.; Kuballa, T.; Rehm, J.; Lachenmeier, D.W. Carcinogenic compounds in alcoholic beverages: An update. Arch. Toxicol. 2016, 90, 2349–2367. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Ramón-Portugal, F.; Andrade, P.; Abreu, S.; de Fatima, T.M.; Strehaiano, P. Malic acid consumption by dry immobilized cells of Schizosaccharomyces pombe. Am. J. Enol. Vitic. 2003, 54, 50–55. [Google Scholar]
- Cecchini, F.; Morassut, M.; Moruno, E.G.; Di Stefano, R. Influence of yeast strain on ochratoxin A content during fermentation of white and red must. Food Microbiol. 2006, 23, 411–417. [Google Scholar] [CrossRef]
- Benito, Á.; Calderón, F.; Benito, S. Combined use of S. pombe and L. thermotolerans in winemaking. Beneficial effects determined through the study of wines’ analytical characteristics. Molecules 2016, 21, 1744. [Google Scholar] [CrossRef] [Green Version]
- Comitini, F.; Capece, A.; Ciani, M.; Romano, P. New insights on the use of wine yeasts. Curr. Opin. Food Sci. 2017, 13, 44–49. [Google Scholar] [CrossRef]
- Cordero-Bueso, G.; Arroyo, T.; Serrano, A.; Tello, J.; Aporta, I.; Vélez, M.D.; Valero, E. Influence of the farming system and vine variety on yeast communities associated with grape berries. Int. J. Food Microbiol. 2011, 145, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Grangeteau, C.; Gerhards, D.; von Wallbrunn, C.; Alexandre, N.; Rousseaux, S.; Guilloux-Benatier, M. Persistence of two non-Saccharomyces yeasts (Hanseniaspora and Starmerella) in the cellar. Front. Microbiol. 2016, 7, 11. [Google Scholar] [CrossRef]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Morata, A.; Benito, S.; Loira, I.; Palomero, F.; González, M.C.; Suárez-Lepe, J.A. Formation of pyranoanthocyanins by Schizosaccharomyces pombe during the fermentation of red must. Int. J. Food Microbiol. 2012, 159, 47–53. [Google Scholar] [CrossRef]
- Lleixà, J.; Martín, V.; Portillo, M.C.; Carrau, F.; Beltran, G.; Mas, A. Comparison of fermentation and wines produced by inoculation of Hanseniaspora vineae and Saccharomyces cerevisiae. Front. Microbiol. 2016, 7, 338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renault, P.; Coulon, J.; Moine, V.; Thibon, C.; Bely, M. Enhanced 3-sulfanylhexan-1-ol production in sequential mixed fermentation with Torulaspora delbrueckii/Saccharomyces cerevisiae reveals a situation of synergistic interaction between two industrial strains. Front. Microbiol. 2016, 7, 22. [Google Scholar] [CrossRef] [Green Version]
- Englezos, V.; Torchio, F.; Cravero, F.; Marengo, F.; Giacosa, S.; Gerbi, V.; Rantsiou, K.; Rolle, L.; Cocolin, L. Aroma profile and composition of Barbera wines obtained by mixed fermentations of Starmerella bacillaris (synonym Candida zemplinina) and Saccharomyces cerevisiae. LWT—Food Sci. Technol. 2016, 73, 567–575. [Google Scholar] [CrossRef]
- Lencioni, L.; Romani, C.; Gobbi, M.; Comitini, F.; Ciani, M.; Domizio, P. Controlled mixed fermentation at winery scale using Zygotorulaspora florentina and Saccharomyces cerevisiae. Int. J. Food Microbiol. 2016, 234, 36–44. [Google Scholar] [CrossRef]
- Tristezza, M.; Tufariello, M.; Capozzi, V.; Spano, G.; Mita, G.; Grieco, F. The oenological potential of Hanseniaspora uvarum in simultaneous and sequential co-fermentation with Saccharomyces cerevisiae for industrial wine production. Front. Microbiol. 2016, 7, 670. [Google Scholar] [CrossRef] [Green Version]
- Oro, L.; Ciani, M.; Bizzaro, D.; Comitini, F. Evaluation of damage induced by Kwkt and Pikt zymocins against Brettanomyces/Dekkera spoilage yeast, as compared to sulphur dioxide. J. Appl. Microbiol. 2016, 121, 207–214. [Google Scholar] [CrossRef]
- Mehlomakulu, N.N.; Setati, M.E.; Divol, B. Characterization of novel killer toxins secreted by wine- related non-Saccharomyces yeasts and their action on Brettanomyces spp. Int. J. Food Microbiol. 2014, 8, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Villalba, M.L.; Susana- Sáez, J.; Del Monaco, S.; Lopes, C.A.; Sangorrín, M.P. TdKT, a new killer toxin produced by Torulaspora delbrueckii effective against wine spoilage yeasts. Int. J. Food Microbiol. 2016, 217, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Zupan, J.; Avbelj, M.; Butinar, B.; Kosel, J.; Šergan, M.; Raspor, P. Monitoring of quorum-sensing molecules during minifermentation studies in wine yeast. J. Agric. Food Chem. 2013, 61, 2496–2505. [Google Scholar] [CrossRef] [PubMed]
- Avbelj, M.; Zupan, J.; Raspor, P. Quorum-sensing in yeast and its potential in wine making. Appl. Microbiol. Biotechnol. 2016, 100, 7841–7852. [Google Scholar] [CrossRef]
- Contreras, A.; Hidalgo, C.; Schmidt, S.; Henschke, P.A.; Curtin, C.; Varela, C. The application of non- Saccharomyces yeast in fermentations with limited aeration as a strategy for the production of wine with reduced alcohol content. Int. J. Food Microbiol. 2015, 205, 7–15. [Google Scholar] [CrossRef]
- Ciani, M.; Morales, P.; Comitini, F.; Tronchoni, J.; Canonico, L.; Curiel, J.A.; Oro, L.; Rodrigues, A.J.; Gonzalez, R. Non-conventional yeast species for lowering ethanol content of wines. Front. Microbiol. 2016, 7, 642. [Google Scholar] [CrossRef] [Green Version]
- Balboa-Lagunero, T.; Arroyo, T.; Cabellos, J.M.; Aznar, M. Yeast selection as a tool for reducing key oxidation notes in organic wines. Food Res. Int. 2013, 53, 252–259. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country | Maxim SO2 during Vinification | % of Organic Vineyard | Label: Organic or Sustainable |
|---|---|---|---|
| Chile | Red: 75 White: 100 | 3 |  Sustainable b Sustainable b |
| Argentina | Red: 70 White: 80 a | 2 |  |
| USA | Use is forbidden | 4.1 |  |
| Europe | Red: 100 White: 150 | 8.5 |  |
| Australia | Red: 100 White: 100 | No data available |  |
| New Zealand | Red: 100 White: 150 | 7 |  |
| South Africa | Red: 90 White: 100 | 2 |  Sustainable b Sustainable b |
| Substance | Application |
|---|---|
| Sorbic acid and sorbates | Microbiological stabilization |
| Lysozyme | |
| Chitosan | |
| L-malic acid, D, L-malic acid | Acidification |
| Ammonium bisulfite | Protection of harvesting |
| Ammonium sulfate | Management of alcoholic fermentation |
| Chitin-glucan | Wine finning |
| Chitosan | |
| Calcium alginate | |
| Co-polymer of vinylimidazole and vinylpyrrolidone (PVI-PVP) | |
| Carboxymethylcellulose (CMC) | Tartrate/Color stabilization |
| Yeast mannoproteins | |
| Polyvinylpolypyrrolidone (PVPP) | Correction of color |
| Beta-glucanase enzymes | Glucan elimination |
| Chitin-glucan | Clarification elimination of heavy metals (iron, copper) |
| Chitosan | |
| Calcium phytate | |
| Potassium ferrocyanide | |
| Urease | Treatment, elimination of ochratoxin A and urea |
| Caramel | Various |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vilela, A. Non-Saccharomyces Yeasts and Organic Wines Fermentation: Implications on Human Health. Fermentation 2020, 6, 54. https://doi.org/10.3390/fermentation6020054
Vilela A. Non-Saccharomyces Yeasts and Organic Wines Fermentation: Implications on Human Health. Fermentation. 2020; 6(2):54. https://doi.org/10.3390/fermentation6020054
Chicago/Turabian StyleVilela, Alice. 2020. "Non-Saccharomyces Yeasts and Organic Wines Fermentation: Implications on Human Health" Fermentation 6, no. 2: 54. https://doi.org/10.3390/fermentation6020054
APA StyleVilela, A. (2020). Non-Saccharomyces Yeasts and Organic Wines Fermentation: Implications on Human Health. Fermentation, 6(2), 54. https://doi.org/10.3390/fermentation6020054