Nitrogen Sources Added to Must: Effect on the Fermentations and on the Tempranillo Red Wine Quality

 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Experimental Design
2.2. General Analytical Parameters
2.3. Aromatic Compound Analysis
2.4. Analysis of Nitrogenous Compounds
2.5. Microorganism Count and Identification
2.6. Sensory Analysis
2.7. Statistical Analysis
3. Results and Discussion
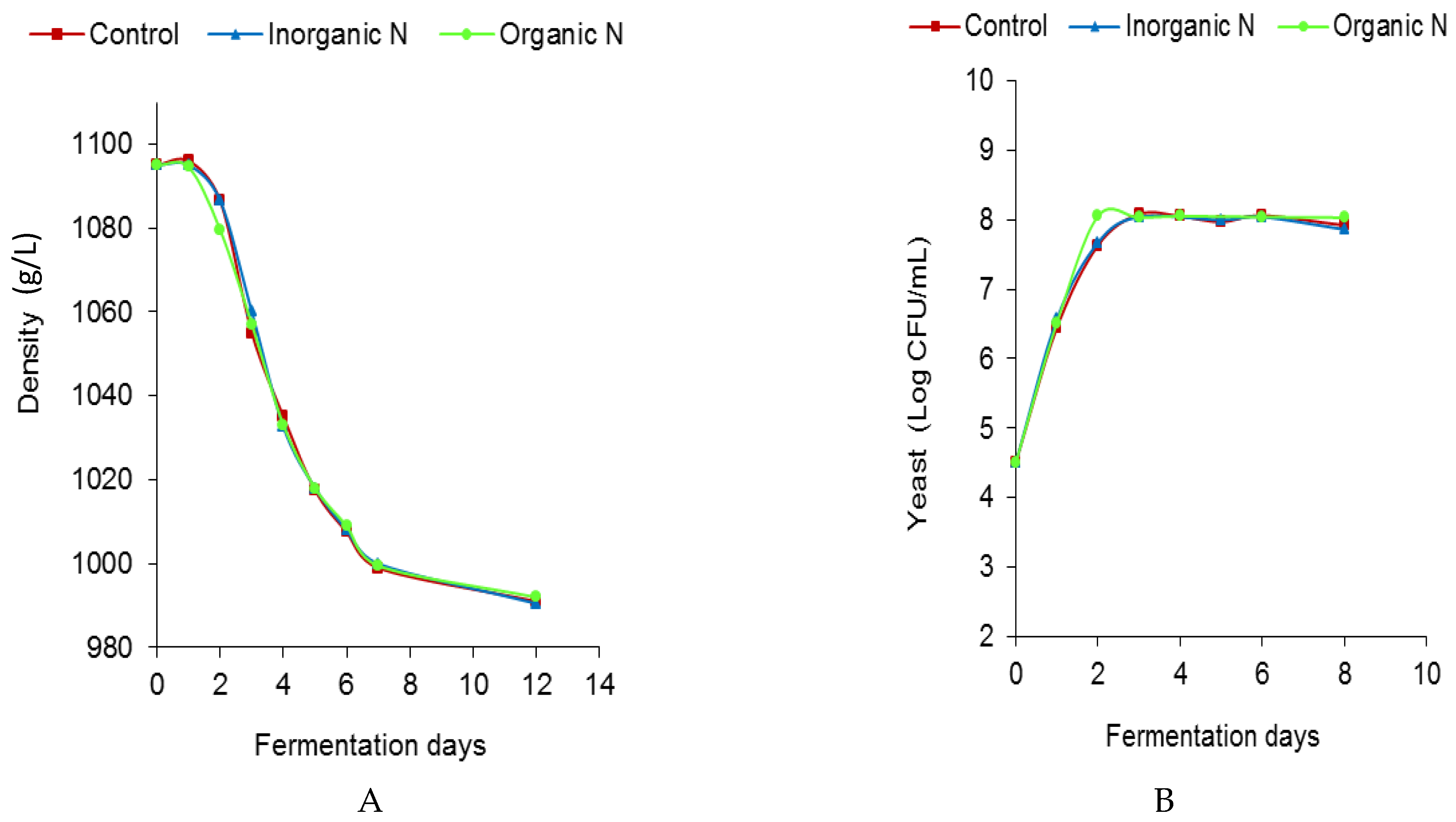
3.1. Influence of Nitrogen Addition on the Development of AF and MLF
3.2. Physicochemical Characteristics of Wines
3.3. Aromatic Composition of Wines
3.4. Nitrogen Composition of Wines
3.4.1. Amino Acids
3.4.2. Biogenic Amines
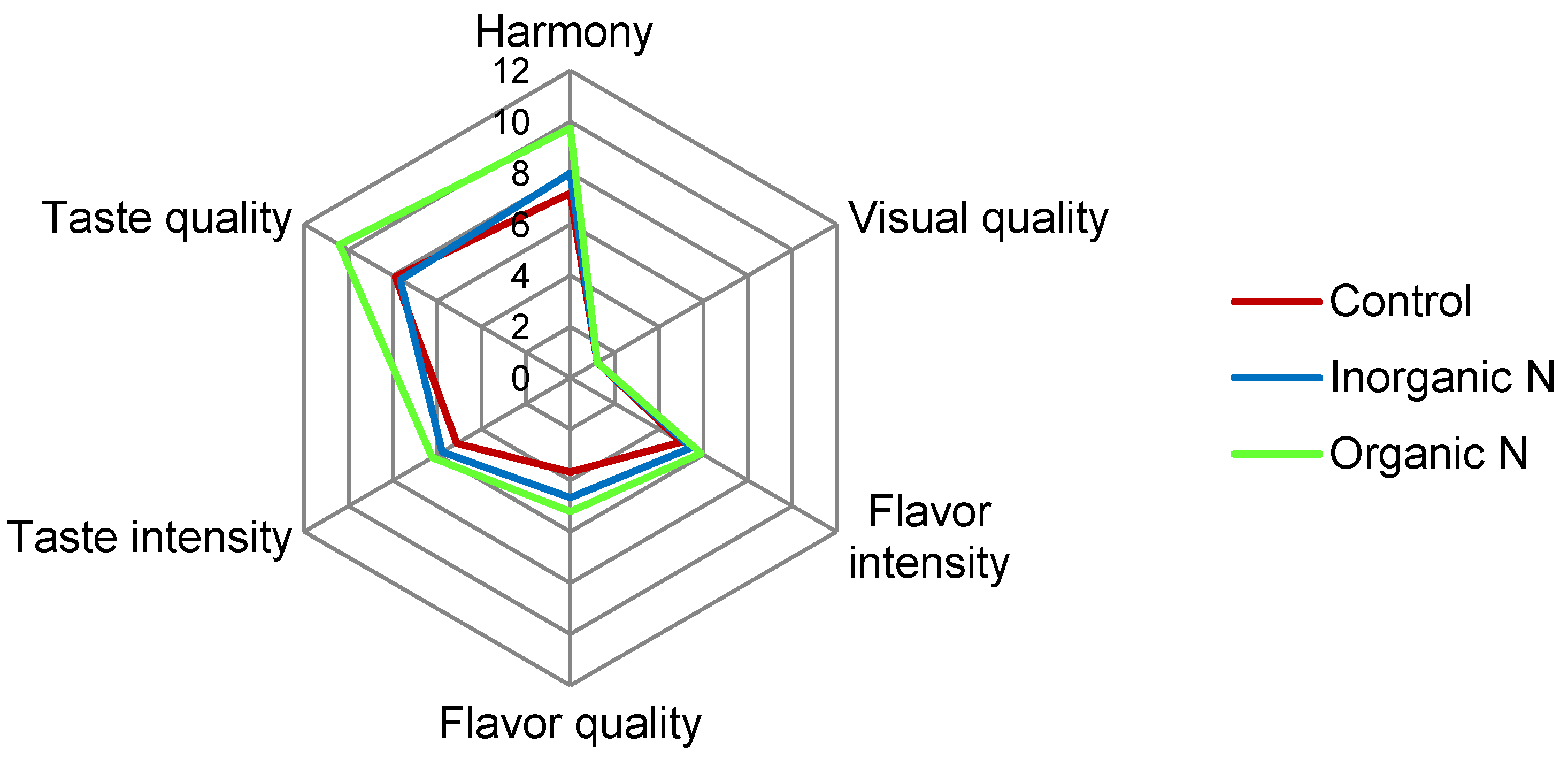
3.5. Organoleptic Analysis of Wines
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bell, S.; Henschke, P. Implications on nitrogen nutrition for grapes, fermentation and wine. Austr. J. Grape Wine Res. 2005, 11, 242–295. [Google Scholar] [CrossRef]
- Beltrán, G.; Esteve-Zarzoso, B.; Rozès, N.; Mas, A.; Guillamón, J.M. Influence of the timing of nitrogen additions during synthetic grape must fermentations on fermentation kinetics and nitrogen consumption. J. Agric. Food Chem. 2005, 53, 996–1002. [Google Scholar] [CrossRef]
- Bisson, L.F. Stuck and sluggish fermentations. Am. J. Enol. Vitic. 1999, 50, 107–119. [Google Scholar]
- Kemsawasd, V.; Viana, T.; Ardö, Y.; Arneborg, N. Influence of nitrogen sources on growth and fermentation performance of different wine yeast species during alcoholic fermentation. Appl. Microbiol. Biotechnol. 2015, 99, 10191–10207. [Google Scholar] [CrossRef] [PubMed]
- Gobert, A.; Tourdot-Maréchal, R.; Sparrow, C.; Morge, C.; Alexandre, H. Influence of nitrogen status in wine alcoholic fermentation. Food Microbiol. 2019, 83, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Jin, G.-J.; Xu, Y.-H.; Xue, S.-J.; Qiao, S.-J.; Teng, Y.-X.; Tao, Y.-S. Enhancing wine ester biosynthesis in mixed Hanseniaspora uvarum/Saccharomyces cerevisiae fermentation by nitrogen nutrient addition. Food Res. Int. 2019, 123, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Kessi-Pérez, E.I.; Molinet, J.; Martínez, C. Disentangling the genetic bases of Saccharomyces cerevisiae nitrogen consumption and adaptation to low nitrogen environments in wine fermentation. Biol. Res. 2020, 53. [Google Scholar]
- Prior, K.J.; Bauer, F.F.; Divol, B. The utilisation of nitrogenous compounds by commercial non-Saccharomyces yeasts associated with wine. Food Microbiol. 2019, 79, 75–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seguinot, P.; Bloem, A.; Brial, P.; Meudec, E.; Ortiz-Julien, A.; Camarasa, C. Analysing the impact of the nature of the nitrogen source on the formation of volatile compounds to unravel the aroma metabolism of two non-Saccharomyces strains. Int. J. Food Microbiol. 2020, 316, 108441. [Google Scholar] [CrossRef]
- Gutiérrez, A.; Chiva, R.; Sancho, M.; Beltran, G.; Arroyo-López, F.N.; Guillamon, J.M. Nitrogen requirements of commercial wine yeast strains during fermentation of a synthetic grape must. Food Microbiol. 2012, 31, 25–32. [Google Scholar] [CrossRef]
- Su, Y.; Origone, A.C.; Rodríguez, M.E.; Querol, A.; Guillamón, J.M.; Lopes, C.A. Fermentative behaviour and competition capacity of cryotolerant Saccharomyces species in different nitrogen conditions. Int. J. Food Microbiol. 2019, 291, 111–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bely, M.; Sablayrolles, J.; Barre, P. Automatic detection of assimilable nitrogen deficiencies during alcoholic fermentation in oenological conditions. J. Ferment. Bioeng. 1990, 70, 246–252. [Google Scholar] [CrossRef]
- Chen, D.; Toussaint, S.; Huanga, W.; Zhan, J.; Liu, S.-Q. Effects of diammonia phosphate addition on the chemical constituents in lychee wine fermented with Saccharomyces cerevisiae. LWT-Food Sci. Technol. 2019, 105, 224–232. [Google Scholar] [CrossRef]
- Wade, M.E.; Osborne, J.P.; Edwards, C.G. Influence of must supplementation on growth of Pediococcus spp. after alcoholic fermentation. S. Afr. J. Enol. Vitic. 2019, 40, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Arribas, M.V.; Polo, M.C.; Jorganes, F.; Muñoz, R. Screening of biogenic amine production by lactic acid bacteria isolated from grape must and wine. Int. J. Food Microbiol. 2003, 84, 117–123. [Google Scholar] [CrossRef] [Green Version]
- Fairbairn, S.; McKinnon, A.; Musarurwa, H.T.; Ferreira, A.C.; Bauer, F.F. The impact of single amino acids on growth and volatile aroma production by Saccharomyces cerevisiae strains. Front. Microbiol. 2017, 8, 2554. [Google Scholar] [CrossRef] [Green Version]
- Kevvai, K.; Kütt, M.L.; Nisamedtinov, I.; Paalme, T. Simultaneous utilization of ammonia, free amino acids and peptides during fermentative growth of Saccharomyces cerevisiae. J. Inst. Brew. 2016, 122, 110–115. [Google Scholar]
- King, E.S.; Osidacz, P.; Curtin, C.; Bastian, S.E.P.; Francis, I.L. Assessing desirable levels of sensory properties in Sauvignon Blanc wines-consumer preferences and contribution of key aroma compounds. Aust. J. Grape Wine Res. 2011, 17, 169–180. [Google Scholar] [CrossRef]
- Lambrechts, M.G.; Pretoris, I.S. Yeast and its importance to wine aroma-a review. S. Afr. J. Enol. Vitic. 2000, 21, 97–129. [Google Scholar] [CrossRef] [Green Version]
- Perez, D.; Assof, M.; Bolcato, E.; Sari, S.; Fanzone, M. Combined effect of temperature and ammonium addition on fermentation profile and volatile aroma composition of Torrontes Riojano wines. LWT-Food Sci. Technol. 2018, 87, 488–497. [Google Scholar] [CrossRef]
- Rollero, S.; Mouret, J.R.; Bloem, A.; Sanchez, I.; Camarasa, C.; Ortiz-Julien, A.; Sablayrolles, J.M.; Dequin, S. Key role of lipid management in nitrogen and aroma metabolism in an evolved wine yeast strain. Microb. Cell Fact. 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- Vilanova, M.; Siebert, T.E.; Varela, C.; Pretorius, I.S.; Henschke, P.A. Effect of ammonium nitrogen supplementation of grape juice on wine volatiles and non-volatiles composition of the aromatic grape variety Albariño. Food Chem. 2012, 133, 124–131. [Google Scholar] [CrossRef]
- Seguinot, P.; Rollero, S.; Sanchez, I.; Sablayrolles, J.M.; Ortiz-Julien, A.; Camarasa, C.; Mouret, J.R. Impacto of timing and the nature of nitrogen additions on the production kinetics of fermentative aromas by Saccharomyces cerevisiae during winemaking fermentation in synthetic media. Food Microbiol. 2018, 76, 29–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipka, Z.; Tanner, H. Une nouvelle méthode de dosage du acide tartarique dans les moûts, les vins et autres bissons (selon Rebelein). Rev. Suisse Vitic. Arboric. Hortic. 1974, 6, 5–10. [Google Scholar]
- Ribéreau-Gayon, P.; Stonestreet, E. Le dosage des anthocyanes dans le vin rouge. Bull. Soc. Chim. Fr. 1965, 9, 2649–2652. [Google Scholar]
- Ortega, C.; López, R.; Cacho, J.; Ferreira, V. Fasta analysis of important wine volatile compounds. Development and validation of a new method based on gas chromatographic-flame ionisation detection analysis of dichloromethane microextracts. J. Chromatogr. A 2001, 923, 205–214. [Google Scholar] [CrossRef]
- Aerny, J. Composés azotés des moûts et des vins. Rev. Suisse Vitic. Arboric. Hortic. 1996, 28, 161–165. [Google Scholar]
- López, R.; Tenorio, C.; Gutiérrez, A.R.; Garde-Cerdán, T.; Garijo, P.; González-Arenzana, L.; López-Alfaro, I.; Santamaria, P. Elaboration of Tempranillo wines at two different pHs. Influence on biogenic amine contents. Food Control. 2012, 25, 583–590. [Google Scholar] [CrossRef]
- Garijo, P.; Santamaría, P.; López, R.; Sanz, S.; Olarte, C.; Gutiérrez, A.R. The occurrence of fungi, yeasts and bacteria in the air of a Spanish winery during vintage. Int. J. Food. Microbiol. 2008, 141–145. [Google Scholar] [CrossRef]
- Zapparoli, G.; Torriani, S.; Presente, P.; Dellaglio, F. Design and evaluation of malolactic enzyme gene targeted primers for rapid identification and detection of Oenococcus oeni in wine. Lett. Appl. Microbiol. 1998, 27, 243–246. [Google Scholar] [CrossRef]
- López, I.; Ruiz-Larrea, F. Genetic typification by pulsed field electrophoresis (PFGE) and randomly amplified polymorphic DNA (RAPD) of wild Lactobacillus plantarum and Oenococcus oeni wine strains. Eur. Food Res. Technol. 2008, 227, 547–555. [Google Scholar] [CrossRef]
- Ruiz, M. La Cata y el Conocimiento de los Vinos; Mundi-Prensa: Madrid, Spain, 1999. [Google Scholar]
- Varela, C.; Pizarro, F.; Agosin, E. Biomass content governs fermentation rate in nitrogen-deficient wine musts. Appl. Environ. Microbiol. 2004, 70, 3392–3400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López, R.; Santamaría, P.; Epifanio, S.; Garijo, P.; Gutiérrez, A.R. La adición de materia nitrogenada al mosto de uva. In Zubía monográfico; Instituto de Estudios Riojanos: Logroño, Spain, 2004; pp. 83–92. [Google Scholar]
- Agenbach, W. A study of must content in relation to incomplete fermentations, yeast production and fermentation activity. In Proceedings of the South African Society for Enology and Viticulture, Cape Town, South Africa, 21–22 November 1977; pp. 66–87. [Google Scholar]
- Bisson, L. Relación entre grados Brix y Nitrógeno fácilmente asimilable. Infowine: Herramientas útiles. 2000. Available online: https://www.infowine.com/es/strumenti_utili/realcin_entre_grados_brix_y_nitrgeno_fcilmente_asimilable_sc_14380.htm (accessed on 3 August 2020).
- Burin, V.M.; Caliari, V.; Bordignon-Luiz, M.T. Nitrogen compounds in must and volatile profile of white wine: Influence of clarification process before alcoholic fermentation. Food Chem. 2016, 202, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Brice, C.; Cubillos, F.A.; Dequin, S.; Camarasa, C.; Martínez, C. Adaptability of the Saccharomyces cerevisiae yeasts to wine fermentation conditions relies on their strong ability to consume nitrogen. PLoS ONE 2018, 13, e0192383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crépin, L.; Nidelet, T.; Sanchez, I.; Dequin, S.; Camarasa, C. Sequential use of nitrogen compounds by Saccharomyces cerevisiae during wine fermentation: A model based on kinetic and regulation characteristics of nitrogen permeases. Appl. Environ. Microbiol. 2012, 78, 8102–8111. [Google Scholar] [CrossRef] [Green Version]
- Vendramini, C.; Beltran, G.; Nadai, C.; Giacomini, A.; Mas, A.; Corich, V. The role of nitrogen uptake on the competition ability of three vineyard Saccharomyces cerevisiae strains. Int. J. Food Microbiol. 2017, 258, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Long, D.; Wilkinson, K.L.; Taylor, D.K.; Jiranek, V. Novel wine yeast for improved utilisation of proline during fermentation. Fermentation 2018, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez, A.R. Selección de Levaduras Vínicas en la D.O.C. Rioja. Ph.D. Thesis, Universidad del Pais Vasco, Leioa, Spain, December 1994. [Google Scholar]
- Torrea, D.; Henschke, P. Ammonium supplementation of grape juice-effect on the aroma profile of Chardonnay wine. Tech. Rev. 2004, 150, 59–63. [Google Scholar]
- Hernández-Orte, P.; Bely, M.; Cacho, J.; Ferreira, V. Impact of ammonium additions on volatile acidity, ethanol, and aromatic compound production by different Saccharomyces cerevisiae strains during fermentation in controlled synthetic media. Aust. J. Grape Wine Res. 2006, 12, 150–160. [Google Scholar] [CrossRef]
- Garde-Cerdán, T.; Ancín-Azpilicueta, C. Effect of the addition of different quantities of amino acids to nitrogen-deficient must on the formation of esters, alcohols, and acids during wine alcoholic fermentation. LWT-Food Sci. Technol. 2008, 41, 501–510. [Google Scholar] [CrossRef]
- Ugliano, M.; Henschke, P.A. Yeasts and wine flavour. In Wine Chemistry and Biochemistry; Moreno-Arribas, M.V., Polo, C., Eds.; Springer: New York, NY, USA, 2008; pp. 328–348. [Google Scholar]
- Epifanio, S. Influencia de la Tecnología de Vinificación en la Microbiología y el Desarrollo de la Fermentación Alcohólica. Ph.D. Thesis, Universidad de La Rioja, Logroño, Spain, July 2005. [Google Scholar]
- Wang, S.Y.; Zhu, H.Z.; Lan, Y.B.; Liu, R.J.; Liu, Y.R.; Zhang, B.L.; Zhu, B.-Q. Modifications of phenolic compounds, biogenic amines, and volatile compounds in Cabernet Gernishct wine through malolactic fermentation by Lactobacillus plantarum and Oenococcus oeni. Fermentation 2020, 6, 15. [Google Scholar] [CrossRef] [Green Version]
- Rapp, A.; Versini, G. Influence of nitrogen compounds in grapes on aroma compounds in wine. In Proceedings of the International Symposium on Nitrogen in Grapes and Wine, Seattle, WA, USA, 18–19 June 1991; American Society for Enology and Viticulture: Seattle, WA, USA, 1991; pp. 156–164. [Google Scholar]
- Barbosa, C.; Mendes-Faia, A.; Mendes-Ferreira, A. The nitrogen source impacts major volatile compounds released by Saccharomyces cerevisiae during alcoholic fermentation. Int. J. Food Microbiol. 2012, 160, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Moreno, R.; Quirós, M.; Morales, P.; González, R. New insights into the advantages of ammonium as a winemaking nutrient. Int. J. Food Microbiol. 2014, 177, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Mouret, J.R.; Perez, M.; Angenieux, M.; Nicolle, P.; Farines, V.; Sablayrolles, J.M. Online-based kinetic analysis of higher alcohol and ester synthesis during winemaking fermentations. Food Bioprocess. Technol. 2014, 7, 1235–1245. [Google Scholar] [CrossRef]
- Dukes, B.; Goldspink, B.; Elliot, J.; Frayne, R. Time of nitrogen fertilisation can reduce fermentation time and improve quality. In Proceedings of the International Symposium on Nitrogen in Grapes and Wine, Seattle, WA, USA, 18–19 June 1991; pp. 249–254. [Google Scholar]
- Bartowsky, E. Oenococcus oeni and malolactic fermentation-moving into molecular arena. Austr. J. Grape Wine Res. 2005, 11, 174–187. [Google Scholar] [CrossRef]
- Sumby, K.M.; Grbin, P.R.; Jiranek, V. Implications of new research and technologies for malolactic fermentation in wine. Appl. Microbiol. Biotechnol. 2014, 98, 8111–8132. [Google Scholar] [CrossRef]
- Ferreira-Monteiro, F.; Bisson, L. Amino acid utilization and urea formation during vinification fermentations. Am. J. Enol. Vitic. 1991, 42, 199–208. [Google Scholar]
- Pozo-Bayon, M.; G.-Alegría, E.; Polo, M.C.; Tenorio, C.; Martín-Alvarez, P.; Calvo de la Banda, M.; Ruiz-Larrea, F.; Moreno-Arribas, M.V. Wine volatile and amino acid composition after Malolactic Fermentation: Effect of Oenococcus oeni and Lactobacillus plantarum starter cultures. J. Agric. Food Chem. 2005, 53, 8729–8736. [Google Scholar] [CrossRef]
- Feuillat, M.; Guilloux-Benatier, M.; Gerbaux, V. Essais d’activation de la fermentation malolactique dans les vins. Sci. Alim. 1985, 5, 103–122. [Google Scholar]
- Lorenzo, C.; Bordiga, M.; Pérez-Álvarez, E.P.; Travaglia, F.; Arlorio, M.; Salinas, M.R.; Coïsson, J.D.; Garde-Cerdán, T. The impacts of temperature, alcoholic degree and amino acids content on biogenic amines and their precursor amino acids content in red wine. Food Res. Int. 2017, 99, 328–335. [Google Scholar] [CrossRef]
- Pramateftaki, P.V.; Metafa, M.; Kallithraka, S.; Lanaridis, P. Evolution of malolactic bacteria and biogenic amines during spontaneous malolactic fermentation in a Greek winery. Lett. Appl. Microbiol. 2006, 43, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Arribas, M.V.; Polo, M.C.; Martín-Alvarez, P. Las Aminas Biógenas del Vino; Informe Presentado en la Reunión de la Comisión Española de la O.I.V.; del Ministerio de Agricultura, Pesca y Alimentación de España: Madrid, Spain, 2005.
- Restuccia, D.; Loizzo, M.R.; Spizzirri, U.G. Accumulation of biogenic amines in wine: Role of alcoholic and malolactic fermentation. Fermentation 2018, 4, 6. [Google Scholar] [CrossRef] [Green Version]
- Beneduce, L.; Romano, A.; Capozzi, V.; Lucas, P.; Barnavon, L.; Bach, B.; Vuchot, P.; Grieco, F.; Spano, G. Biogenic amine in wines. Ann. Microbiol. 2010, 60, 573–578. [Google Scholar] [CrossRef]
- Landete, J.M.; Ferrer, S.; Polo, L.; Pardo, I. Biogenic amines in wines from three Spanish regions. J. Agric. Food Chem. 2005, 53, 1119–1124. [Google Scholar] [CrossRef] [PubMed]
- Smit, A.Y.; Engelbrecht, L.; du Toit, M. 2012. Managing your wine fermentation to reduce the risk of biogenic amine formation. Front. Microbiol. 2012. [Google Scholar] [CrossRef] [Green Version]
- Bordiga, M.; Guzzon, R.; Larcher, R.; Travaglia, F.; Arlorio, M.; Coïsson, J.D. Variation in content of tyramine, histamine, 2-phenylethylamine, tryptamine and their precursor amino acids in a Chardonnay wine by using different commercial active dry yeasts and nitrogen sources. Int. J. Food Sci. Tech. 2020, 55, 559–568. [Google Scholar] [CrossRef]
- Martínez-Pinilla, O.; Guadalupe, Z.; Hernández, Z.; Ayestarán, B. Amino acids and biogenic amines in red varietal wines: The role of grape variety, malolactic fermentation and vintage. Eur. Food Res. Technol. 2013, 237, 887–895. [Google Scholar] [CrossRef]
- Berbegal, C.; Benavent-Gil, Y.; Navascués, E.; Calvo, A.; Albors, C.; Pardo, I.; Ferrer, S. Lowering histamine formation in a red Ribera de Duero wine (Spain) by using an indigenous O. oeni strain as a malolactic starter. Int. J. Food Microbiol. 2017, 244, 11–18. [Google Scholar] [CrossRef]
- Ferrer, S.; Landete, J.; Polo, L.; Pardo, I. Las bacterias y su repercusión sobre las aminas biógenas. In Proceedings of the International Symposium Microsafetywine, Vilafranca del Penedés, Spain, 20–21 November 2007; pp. 914–916. [Google Scholar]
- Landete, J.M.; Ferrer, S.; Pardo, I. Biogenic amine production by lactic acid bacteria, acetic bacteria and yeast isolated from wine. Food Control. 2007, 18, 1569–1574. [Google Scholar] [CrossRef]
- Torrea, D.; Varela, C.; Ugliano, M.; Ancin-Azpilicueta, C.; Leigh Francis, I.; Henschke, P.A. Comparison of inorganic and organic nitrogen supplementation of grape juice-Effect on volatile composition and aroma profile of a Chardonnay wine fermented with Saccharomyces cerevisiae yeast. Food Chem. 2011, 127, 1072–1083. [Google Scholar] [CrossRef]
- Ferrari, G. Influence de la composition azotée des moûts sur la qualité des vins et eaux-de-vie. Realtions vec les arômes et les defaults. J. Int. Sci. Vigne. Vin. 2002, 36, 1–10. [Google Scholar]
- Ugliano, M.; Travis, B.; Francis, I.L.; Henschke, P.A. Volatile composition and sensory properties of Shiraz wines as affected by nitrogen supplementation and yeast species: Rationalizing nitrogen modulation of wine aroma. J. Agric. Food Chem. 2010, 58, 12417–12425. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Orte, P.; Ibarz, M.J.; Cacho, J.; Ferreira, V. Effect of the addition of ammonium and amino acids to musts of Airen variety on aromatic composition and sensory properties of the obtained wine. Food Chem. 2004, 89, 163–174. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Physicochemical Parameters | After AF | After MLF | ||||
|---|---|---|---|---|---|---|
| Control | Inorganic N | Organic N | Control | Inorganic N | Organic N | |
| Ethanol (% v/v) | 13.8 | 13.7 | 13.8 | 13.8 | 13.7 | 13.8 |
| Reducing sugars (g/L) | 2.15 | 2.16 | 2.11 | 2.15 | 2.16 | 2.11 |
| pH | 3.53 | 3.53 | 3.53 | 3.59 | 3.59 | 3.60 |
| Total acidity (tartaric acid g/L) | 7.36 | 7.25 | 7.40 | 6.50 | 6.03 | 6.15 |
| Tartaric acid (g/L) | 2.73 | 2.61 | 2.89 | 3.11 | 3.00 | 3.15 |
| Malic acid (g/L) | 2.34 | 2.22 | 2.36 | nd * | nd | nd |
| Lactic acid (g/L) | nd | nd | nd | 1.48 | 1.48 | 1.58 |
| Volatile acidity (acetic acid g/L) | 0.16 a | 0.15 a | 0.20 b | 0.21 | 0.23 | 0.25 |
| Citric acid (mg/L) | 228 | 231 | 238 | 103 | 92.7 | 99 |
| Glycerol (g/L) | 9.79 | 11.9 | 8.50 | 8.58 | 8.93 | 8.35 |
| Color intensity | 22.1 | 22.8 | 24.0 | 15.2 | 15.7 | 15.7 |
| Hue | 0.394 | 0.393 | 0.408 | 0.462 | 0.466 | 0.474 |
| TPI | 63.2 | 66.1 | 66.9 | 61.6 | 63.1 | 62.1 |
| Anthocyanins (mg/L) | 1251 a | 1232 a | 1299 b | 1032 | 1049 | 1064 |
| Urea (mg/L) | 3.76 | 2.32 | 3.55 | 2.73 | 4.49 | 4.93 |
| Aromatic Compounds (mg/L). | After AF | After MLF | ||||
|---|---|---|---|---|---|---|
| Control | Inorganic N | Organic N | Control | Inorganic N | Organic N | |
| Alcohols | ||||||
| 1-Propanol | 20.0 a | 26.8 b | 20.6 a | 19.0 a | 24.4 b | 18.4 a |
| Isobutanol | 34.4 a | 42.6 b | 34.2 a | 32.6 a | 38.9 b | 32.4 a |
| 1-Hexanol | 2.12 | 2.35 | 2.17 | 1.99 | 1.92 | 1.99 |
| 2 + 3-Methyl-1-butanol | 230 a | 291 b | 237 a | 226 a | 268 b | 228 a |
| 2-Phenyl ethanol | 51.2 a | 61.2 b | 53.6 a | 46.1 | 48.4 | 48.9 |
| cis-3-hexenol | 0.40 | 0.47 | 0.37 | 0.37 | 0.38 | 0.34 |
| Methionol | 0.15 | 0.15 | 0.16 | 0.26 | 0.31 | 0.30 |
| Benzyl alcohol | 0.10 b | 0.05 a | 0.05 a | 0.09 b | 0.06 a | 0.05 a |
| Esters | ||||||
| 2-Phenylethyl acetate | 0.27 | 0.33 | 0.2 | 30.21 | 0.26 | 0.21 |
| Diethyl succinate | 2.51 | 2.08 | 2.39 | 2.17 | 2.01 | 2.60 |
| Ethyl-3 hidroxybutyrate | 0.87 a | 1.00 b | 0.82 a | 0.80 a | 0.92 b | 0.84 a |
| Ethyl acetate | 48.5 | 50.4 | 45.5 | 51.6 | 48.6 | 53.1 |
| Ethyl butyrate | 0.32 | 0.37 | 0.28 | 0.33 | 0.39 | 0.32 |
| Ethyl hexanoate | 0.25 | 0.27 | 0.15 | 0.18 | 0.20 | 0.20 |
| Ethyl isobutyrate | 0.01 | 0.02 | 0.01 | 0.02 | 0.04 | 0.03 |
| Ethyl lactacte | 1.15 | 1.20 | 1.23 | 25.2 | 25.5 | 26.2 |
| Ethyl octanoate | 0.07 | 0.05 | 0.02 | 0.06 | 0.12 | 0.14 |
| Ethyl propionate | 0.10 | 0.15 | 0.09 | 0.12 | 0.15 | 0.12 |
| Hexyl acetate | 0.06 | 0.19 | 0.13 | nd | nd | nd |
| Isoamyl acetate | 3.78 a | 4.69 b | 3.26 a | 3.41 a | 3.98 b | 3.55 a |
| Acids | ||||||
| Butyric acid | 2.02 | 2.14 | 1.89 | 1.79 | 1.98 | 1.90 |
| Isobutiric acid | 2.47 a | 3.09 b | 2.72 ab | 2.82 a | 3.42 b | 2.83 a |
| Isovaleric acid | 1.94 a | 2.38 b | 1.97 a | 1.74 a | 2.09 b | 1.87 a |
| Hexanoic acid | 3.42 | 3.42 | 3.62 | 3.39 | 3.12 | 3.63 |
| Octanoic acid | 1.79 | 1.56 | 1.76 | 2.11 | 2.22 | 2.21 |
| Decanoic acid | 0.18 | 0.21 | 0.13 | 0.21 | 0.22 | 0.20 |
| Other compounds | ||||||
| Acetaldehyde | 7.04 | 7.91 | 7.34 | 2.79 | 1.79 | 1.61 |
| Acetoin | 0.75 | 1.01 | 0.90 | 4.82 | 4.62 | 4.38 |
| Diacetyl | 1.15 | 1.31 | 1.49 | 3.02 | 2.64 | 2.45 |
| Butyrolactone | 1.57 | 1.59 | 1.40 | 2.85 | 3.31 | 3.18 |
| Amino Acids(mg/L) | Elaboration Stage | ||||
|---|---|---|---|---|---|
| Treatment | Must | After FA | After MLF | After 5 Months | |
| Aspartic acid | Control | 9.62 c | nd * a | nd a | 2.42 b |
| Inorganic N | 9.62 c | nd a | nd a | 2.40 b | |
| Organic N | 9.62 c | nd a | nd a | 3.19 b | |
| Glutamic acid | Control | 31.4 c | 11.4 b | 6.28 a | 9.22 b |
| Inorganic N | 31.4 c | 9.65 b | 5.77 a | 7.98 b | |
| Organic N | 31.4 c | 10.1 b | 8.01 a | 11.5 b | |
| Asparagine | Control | 1.98 a | 2.61 ab | 2.77 ab | 3.37 b |
| Inorganic N | 1.98 | 2.94 | 3.02 | 3.76 | |
| Organic N | 1.98 a | 2.80 ab | 3.14 bc | 4.18 c | |
| Serine | Control | 11.4 c | 0.76 a | 1.00 a | 1.67 b |
| Inorganic N | 11.4 b | 1.05 a | 1.33 a | 1.71 a | |
| Organic N | 11.4 c | 1.03 a | 1.37 ab | 2.09 b | |
| Glutamine | Control | 25.3 b | 2.56 a | 2.47 a | 1.18 a |
| Inorganic N | 25.3 b | 2.60 a | 1.99 a | 1.12 a | |
| Organic N | 25.3 b | 2.52 a | 1.98 a | 1.18 a | |
| Histidine | Control | 5.41 b | 1.30 a | 0.86 a | 2.42 a |
| Inorganic N | 5.41 b | 1.30 a | 1.41 a | 2.53 a | |
| Organic N | 5.41 c | 1.21 a | 1.75 ab | 3.14 b | |
| Glycine + Threonine | Control | 8.04 b | 4.17 a | 4.79 ab | 7.61 ab |
| Inorganic N | 8.04 b | 4.00 a | 5.59 ab | 8.52 b | |
| Organic N | 8.04 c | 3.82 a | 6.65 b | 9.91 c | |
| Citruline | Control | 1.76 | 1.72 | 1.44 | 1.15 |
| Inorganic N | 1.76 | 0.97 | 1.15 | 1.15 | |
| Organic N | 1.76 | 0.96 | 1.47 | 1.19 | |
| Arginine | Control | 30.5 b | 1.18 a | 1.18 a | 2.06 a |
| Inorganic N | 30.5 b | 1.51 a | 1.01 a | 2.22 a | |
| Organic N | 30.5 b | 1.07 a | 1.95 a | 3.60 a | |
| Alanine | Control | 13.7 c | 4.18 a | 5.43 ab | 7.26 b |
| Inorganic N | 13.7 c | 4.53 a | 5.90 ab | 7.84 b | |
| Organic N | 13.7 d | 4.48 a | 7.07 b | 9.76 c | |
| Tirosine | Control | 1.80 b | 0.56 a | 1.37 b | 1.78 b |
| Inorganic N | 1.80 b | 0.83 a | 1.44 b | 1.95 b | |
| Organic N | 1.80 b | 0.65 a | 1.63 b | 2.39 b | |
| Cystine | Control | nd a | 1.48 b | 1.47 b | nd a |
| Inorganic N | nd a | 1.32 b | 1.51 b | nd a | |
| Organic N | nd a | 0.91 b | 1.55 b | nd a | |
| Valine | Control | 12.1 c | nd a | nd a | 1.63 b |
| Inorganic N | 12.1 c | nd a | nd a | 1.71 b | |
| Organic N | 12.1 c | nd a | nd a | 2.18 b | |
| Methionine | Control | 1.79 c | nd a | nd a | 0.93 b |
| Inorganic N | 1.79 c | nd a | nd a | 0.98 b | |
| Organic N | 1.79 c | nd a | nd a | 1.24 b | |
| Tryptophan | Control | 11.4 b | 0.99 a | 1.43 a | 1.10 a |
| Inorganic N | 11.4 b | 1.29 a | 1.17 a | 1.11 a | |
| Organic N | 11.4 b | 0.74 a | 1.20 a | 1.32 a | |
| Phenylalanine | Control | 3.36 c | 0.54 a | 1.77 b | 3.13 c |
| Inorganic N | 3.36 c | 0.74 a | 1.84 b | 3.44 c | |
| Organic N | 3.36 c | 0.25 a | 2.32 b | 4.08 d | |
| Isoleucine | Control | 2.77 d | 0.27 a | 0.89 b | 1.27 c |
| Inorganic N | 2.77 c | 0.78 a | 1.02 a | 1.33 a | |
| Organic N | 2.77 c | 0.18 a | 1.21 b | 1.67 b | |
| Leucine | Control | 2.88 bc | 0.56 a | 2.70 b | 3.48 c |
| Inorganic N | 2.88 bc | 0.78 a | 2.44 b | 3.80 c | |
| Organic N | 2.88 b | 0.52 a | 3.41 b | 4.87 d | |
| Lysine | Control | nd a | nd a | nd a | 6.36 b |
| Inorganic N | nd a | nd a | nd a | 8.74 b | |
| Organic N | nd a | nd a | nd a | 7.77 b | |
| Sum of amino acids-proline | Control | 175.21 b | 34.28 a | 35.85 a | 58.04 a |
| Inorganic N | 175.21 c | 34.29 a | 36.59 a | 62.29 b | |
| Organic N | 175.21 d | 31.24 a | 44.71 b | 75.26 c | |
| Proline | Control | 338 a | 564 b | 613 b | 624 b |
| Inorganic N | 338 | 456 | 482 | 490 | |
| Organic N | 338 a | 564 b | 605 b | 639 b | |
| Biogenic Amines(mg/L) | After MLF | Five Months after Final MLF | ||||
|---|---|---|---|---|---|---|
| Control | Inorganic N | Organic N | Control | Inorganic N | Organic N | |
| Histamine | nd * | nd | nd | nd | nd | nd |
| Ethylamine | 1.05 | 1.16 | 0.99 | 1.02 | 1.10 | 1.08 |
| Tyramine | nd | nd | nd | nd | nd | nd |
| Phenylethylamine | nd | nd | nd | nd | nd | nd |
| Putrescine | 2.05 | 1.88 | 2.36 | 2.12 | 1.96 | 2.16 |
| Isoamylamine | nd | nd | nd | nd | nd | nd |
| Cadaverine | 0.33 | 0.33 | 0.33 | 0.34 | 0.35 | 0.34 |
| Sum of biogenic amines | 3.43 | 3.37 | 3.68 | 3.48 | 3.41 | 3.58 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santamaría, P.; González-Arenzana, L.; Garijo, P.; Gutiérrez, A.R.; López, R. Nitrogen Sources Added to Must: Effect on the Fermentations and on the Tempranillo Red Wine Quality. Fermentation 2020, 6, 79. https://doi.org/10.3390/fermentation6030079
Santamaría P, González-Arenzana L, Garijo P, Gutiérrez AR, López R. Nitrogen Sources Added to Must: Effect on the Fermentations and on the Tempranillo Red Wine Quality. Fermentation. 2020; 6(3):79. https://doi.org/10.3390/fermentation6030079
Chicago/Turabian StyleSantamaría, Pilar, Lucía González-Arenzana, Patrocinio Garijo, Ana Rosa Gutiérrez, and Rosa López. 2020. "Nitrogen Sources Added to Must: Effect on the Fermentations and on the Tempranillo Red Wine Quality" Fermentation 6, no. 3: 79. https://doi.org/10.3390/fermentation6030079
APA StyleSantamaría, P., González-Arenzana, L., Garijo, P., Gutiérrez, A. R., & López, R. (2020). Nitrogen Sources Added to Must: Effect on the Fermentations and on the Tempranillo Red Wine Quality. Fermentation, 6(3), 79. https://doi.org/10.3390/fermentation6030079