
Lactate Metabolism and Microbiome Composition Are Affected by Nitrogen Gas Supply in Continuous Lactate-Based Chain Elongation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Substrate, Mineral Medium, and Inoculum
2.2. Reactor Setup and Operation
2.3. Calculations
2.4. DNA Extraction and Sequencing Analysis
2.5. Analytical Methods
3. Results
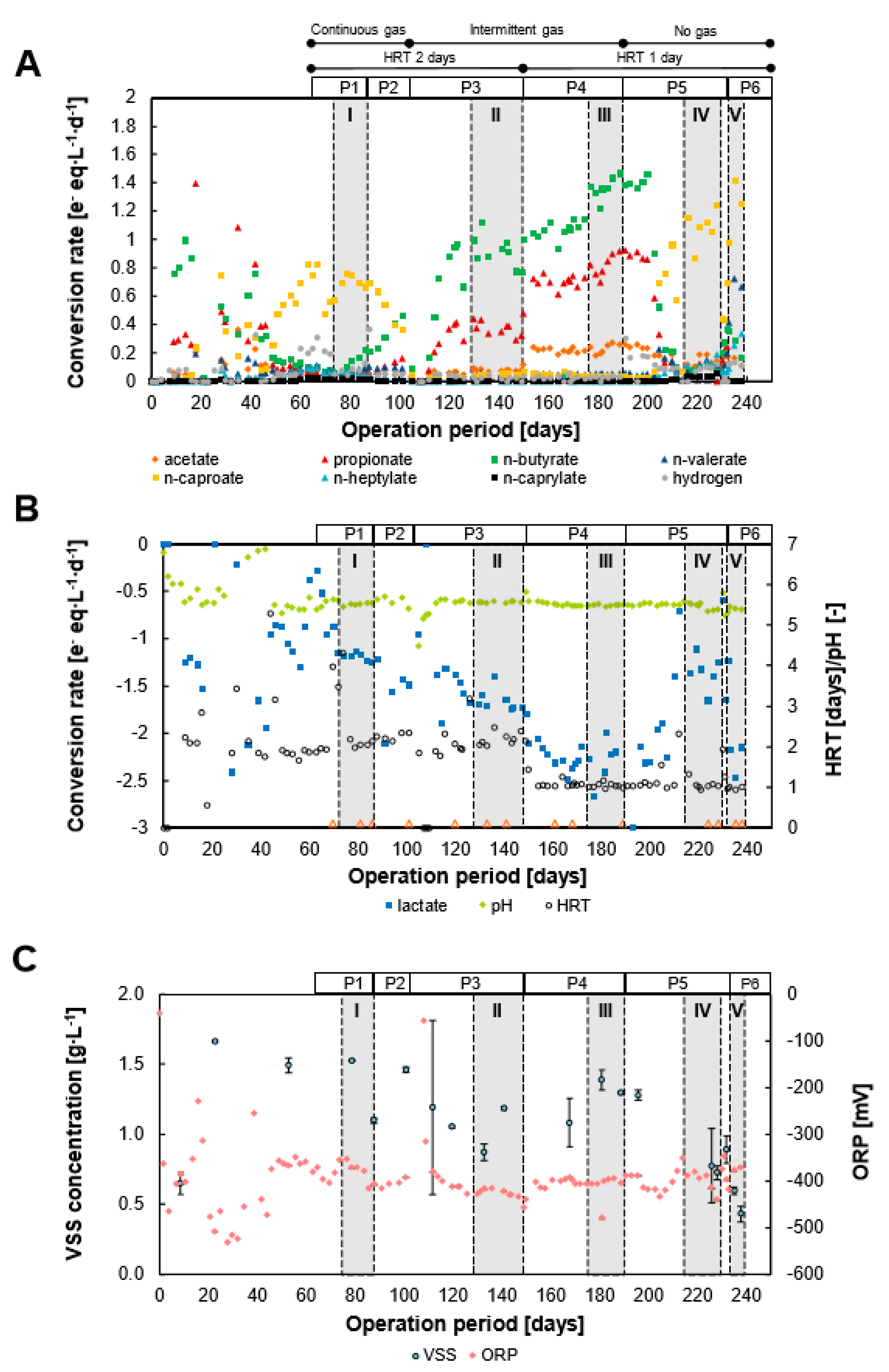
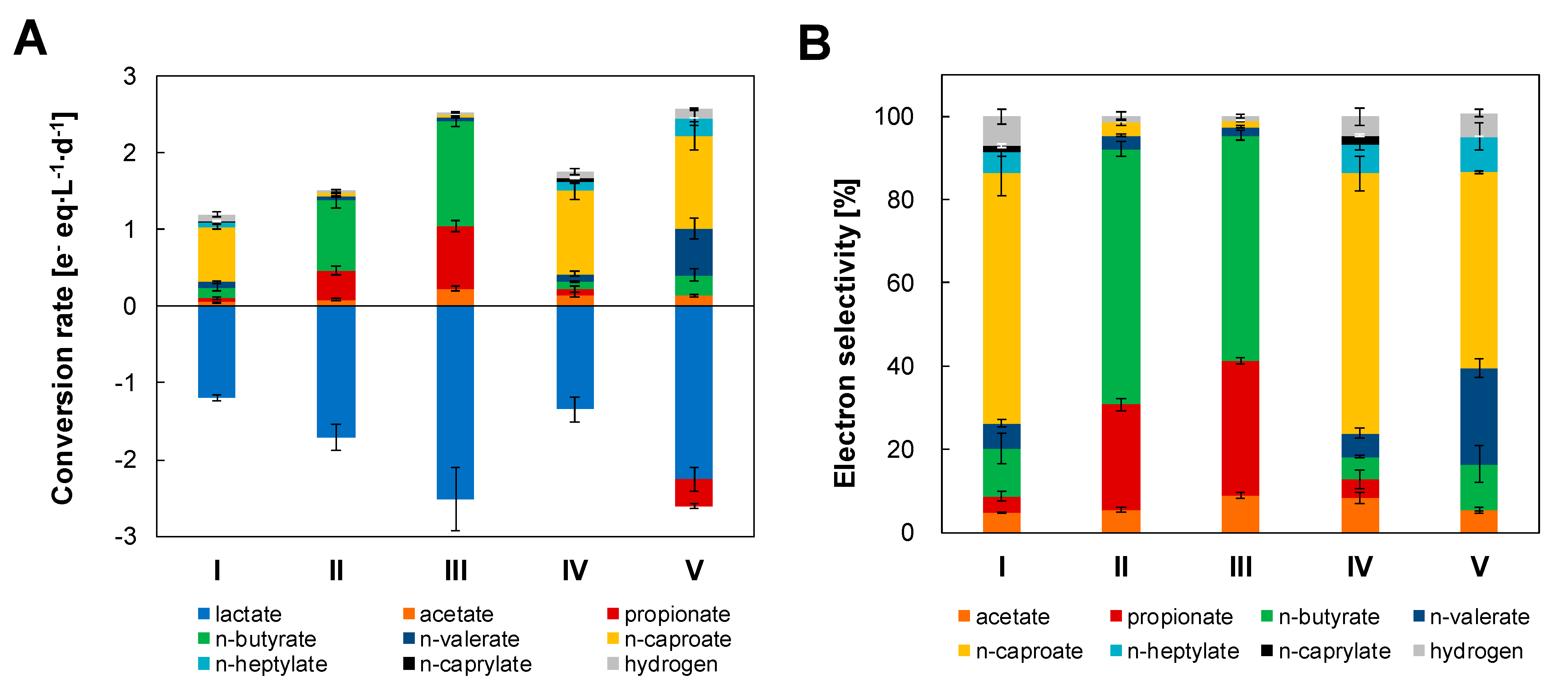
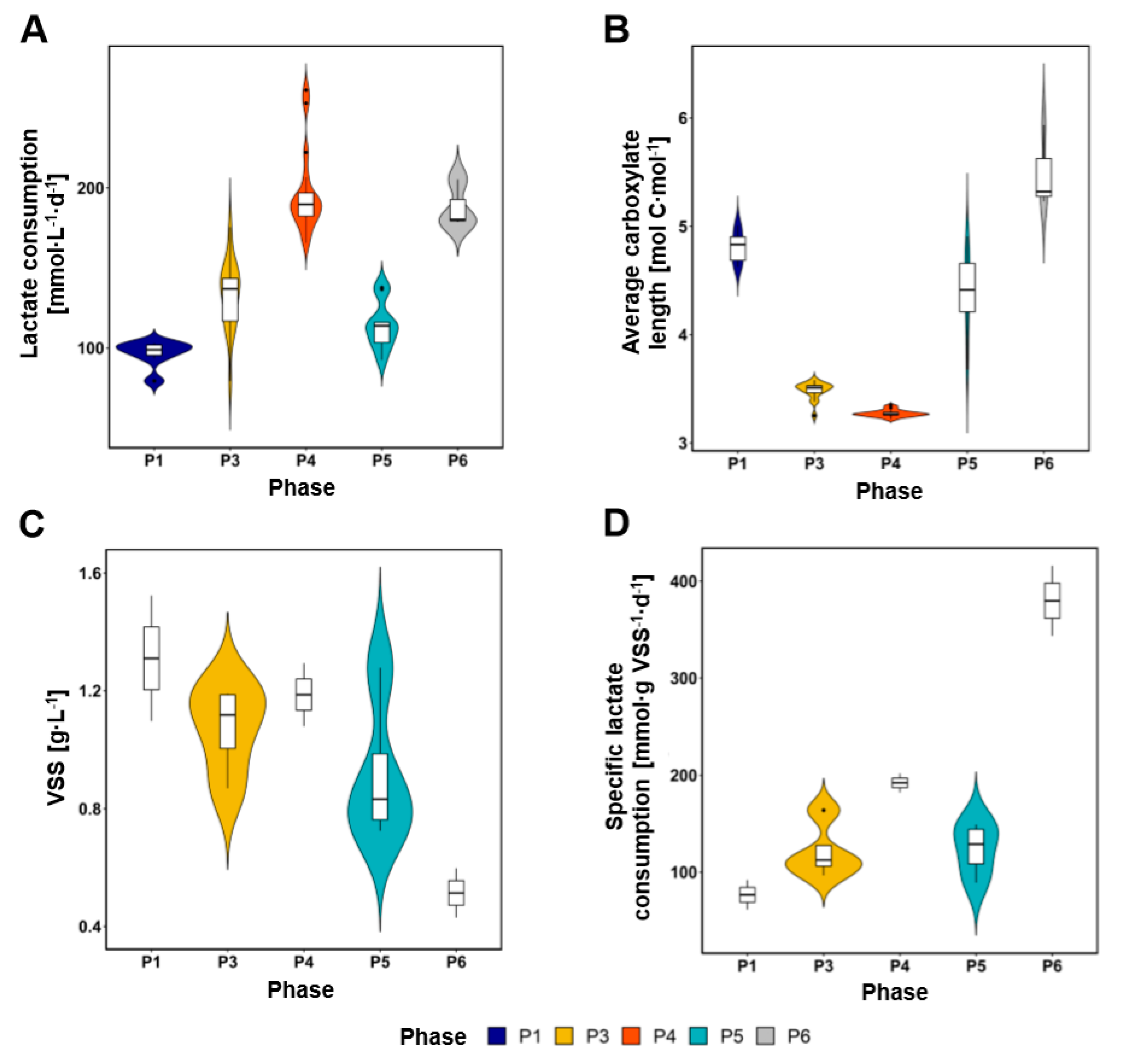
3.1. Lactate Conversion Performance and Biomass Growth
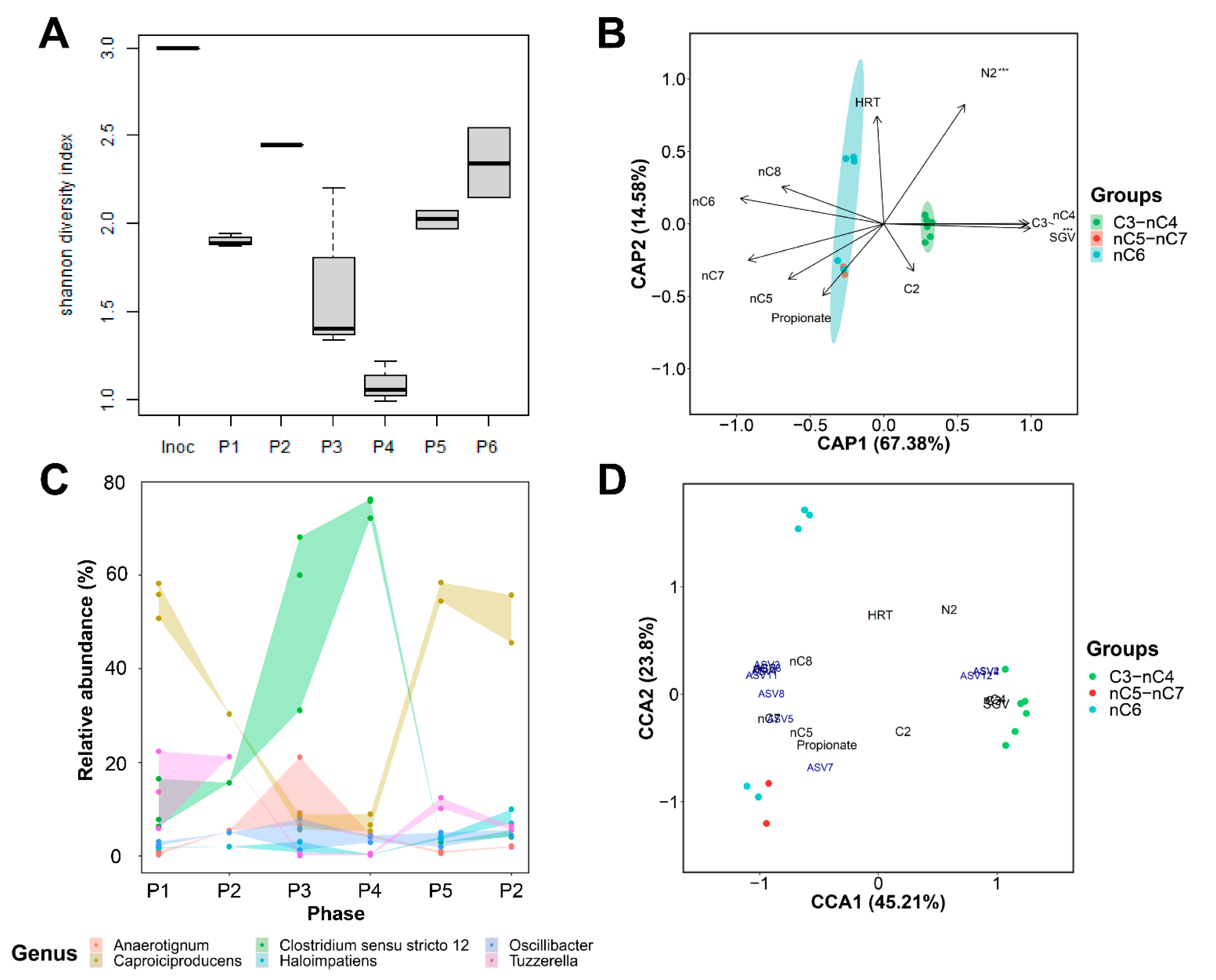
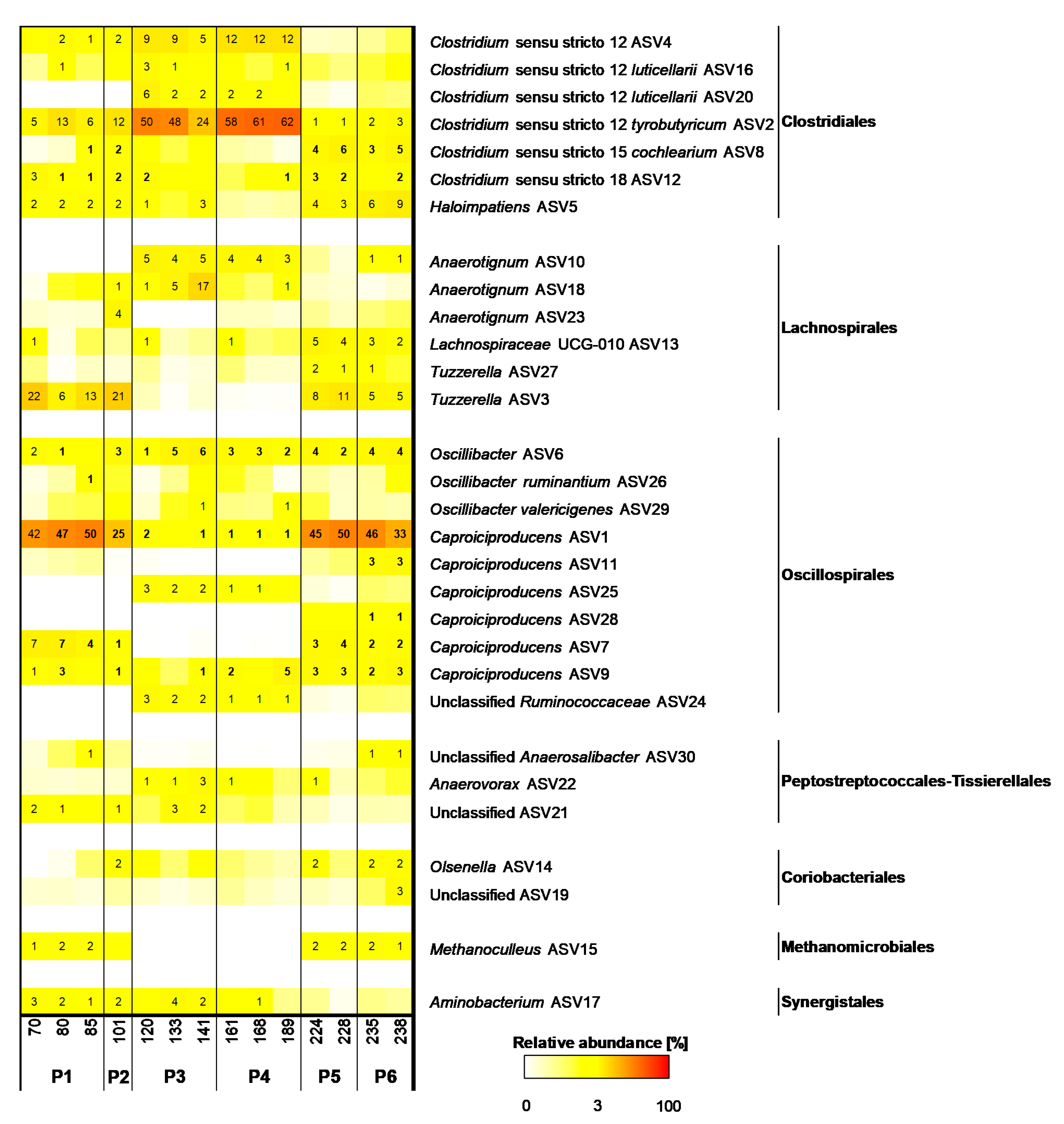
3.2. Caproiciproducens Outcompeted by Clostridium at High Superficial Gas Velocities
4. Discussion
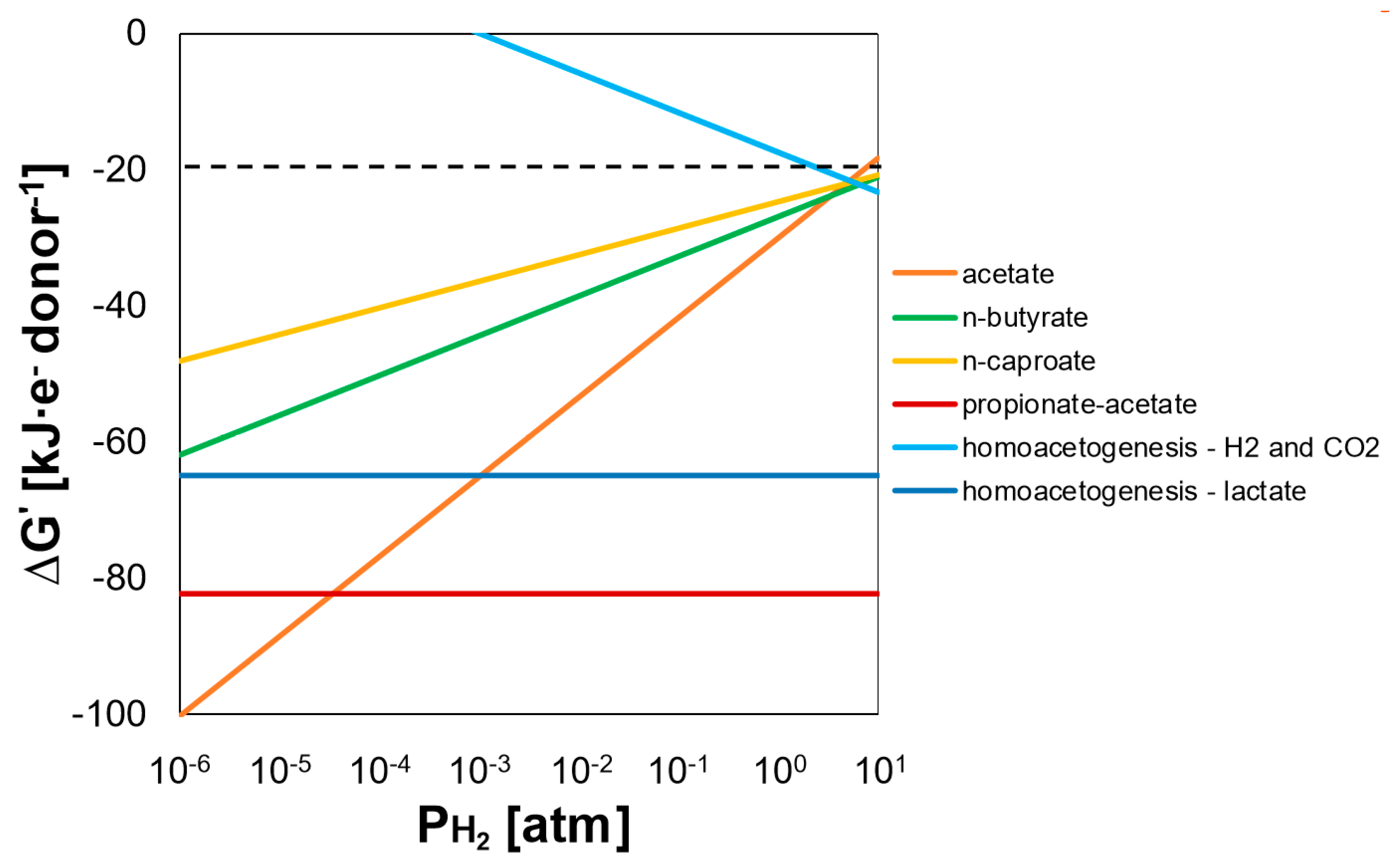
4.1. Lactate Was Efficiently Converted into MCC at Low Gas Velocities with Conversion Rates Improved after Electron Acceptor Supplementation
4.2. High Gas Velocities and Dilution Rates May Favor Fast Lactate Metabolism
4.3. Shift from MCC to SCC Production Was Unrelated to Hydrogen Partial Pressure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Angenent, L.T.; Richter, H.; Buckel, W.; Spirito, C.M.; Steinbusch, K.J.J.; Plugge, C.M.; Strik, D.P.B.T.B.; Grootscholten, T.I.M.; Buisman, C.J.N.; Hamelers, H.V.M. Chain Elongation with Reactor Microbiomes: Open-Culture Biotechnology to Produce Biochemicals. Environ. Sci. Technol. 2016, 50, 2796–2810. [Google Scholar] [CrossRef]
- Sun, Z.; Ramsay, J.A.; Guay, M.; Ramsay, B.A. Carbon-limited fed-batch production of medium-chain-length polyhydroxyalkanoates from nonanoic acid by Pseudomonas putida KT2440. Appl. Microbiol. Biotechnol. 2007, 74, 69–77. [Google Scholar] [CrossRef]
- Costa, S.; Summa, D.; Semeraro, B.; Zappaterra, F.; Rugiero, I.; Tamburini, E. Fermentation as a Strategy for Bio-Transforming Waste into Resources: Lactic Acid Production from Agri-Food Residues. Fermentation 2021, 7, 3. [Google Scholar] [CrossRef]
- Contreras-Dávila, C.A.; Carrión, V.J.; Vonk, V.R.; Buisman, C.N.J.; Strik, D.P.B.T.B. Consecutive lactate formation and chain elongation to reduce exogenous chemicals input in repeated-batch food waste fermentation. Water Res. 2020, 169, 1–10. [Google Scholar] [CrossRef]
- Kim, M.-S.; Na, J.-G.; Lee, M.-K.; Ryu, H.; Chang, Y.-K.; Triolo, J.M.; Yun, Y.-M.; Kim, D.-H. More value from food waste: Lactic acid and biogas recovery. Water Res. 2016, 96, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Candry, P.; Radić, L.; Favere, J.; Carvajal-Arroyo, J.M.; Rabaey, K.; Ganigué, R. Mildly acidic pH selects for chain elongation to caproic acid over alternative pathways during lactic acid fermentation. Water Res. 2020, 186. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Tao, Y.; Liang, C.; Li, X.; Wei, N.; Zhang, W.; Zhou, Y.; Yang, Y.; Bo, T. The synthesis of n-caproate from lactate: A new efficient process for medium-chain carboxylates production. Sci. Rep. 2015, 5, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Kucek, L.A.; Nguyen, M.; Angenent, L.T. Conversion of L-lactate into n-caproate by a continuously fed reactor microbiome. Water Res. 2016, 93, 163–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seeliger, S.; Janssen, P.H.; Schink, B. Energetics and kinetics of lactate fermentation to acetate and propionate via methylmalonyl-CoA or acrylyl-CoA. FEMS Microbiol. Lett. 2002, 211, 65–70. [Google Scholar] [CrossRef]
- Grootscholten, T.I.M.; Steinbusch, K.J.J.; Hamelers, H.V.M.; Buisman, C.J.N. High rate heptanoate production from propionate and ethanol using chain elongation. Bioresour. Technol. 2013, 136, 715–718. [Google Scholar] [CrossRef]
- Carvajal-Arroyo, J.M.; Candry, P.; Andersen, S.J.; Props, R.; Seviour, T.; Ganigué, R.; Rabaey, K. Granular fermentation enables high rate caproic acid production from solid-free thin stillage. Green Chem. 2019, 21, 1330–1339. [Google Scholar] [CrossRef]
- Roghair, M.; Hoogstad, T.; Strik, D.P.B.T.B.; Plugge, C.M.; Timmers, P.H.A.; Weusthuis, R.A.; Bruins, M.E.; Buisman, C.J.N. Controlling Ethanol Use in Chain Elongation by CO2 Loading Rate. Environ. Sci. Technol. 2018, 52, 1496–1505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinbusch, K.J.J.; Hamelers, H.V.M.; Plugge, M.; Buisman, C.J.N. Biological formation of caproate and caprylate from acetate: Fuel and chemical production from low grade biomass. Energy Environ. Sci. 2011, 4, 216–224. [Google Scholar] [CrossRef]
- Wu, Q.; Guo, W.; You, S.; Bao, X.; Luo, H.; Wang, H.; Ren, N. Concentrating lactate-carbon flow on medium chain carboxylic acids production by hydrogen supply. Bioresour. Technol. 2019, 291. [Google Scholar] [CrossRef]
- Noorman, H.J.; Van Winden, W.; Heijnen, J.J.; Van Der Lans, R.G.J.M. Intensified Fermentation Processes and Equipment. In Intensification of Biobased Processes; Górak, A., Stankiewicz, A., Eds.; The Royal Society of Chemistry: Cambridge, UK, 2018; pp. 1–41. ISBN 9781782624127. [Google Scholar]
- EFSA (European Food Safety Authority). Evaluation of the application for a new alternative processing method for animal by-products of Category 3 material (ChainCraft B.V.). EFSA J. 2018, 16. [CrossRef]
- Wu, J.; Zhou, H.; Li, H.; Zhang, P.; Jiang, J. Impacts of hydrodynamic shear force on nucleation of flocculent sludge in anaerobic reactor. Water Res. 2009, 43, 3029–3036. [Google Scholar] [CrossRef]
- Tay, H.; Liu, S. The effects of shear force on the formation, structure and metabolism of aerobic granules. Appl. Microbiol. Biotechnol. 2001, 57, 227–233. [Google Scholar] [CrossRef]
- Roghair, M.; Strik, D.P.B.T.B.; Steinbusch, K.J.J.; Weusthuis, R.A.; Bruins, M.E.; Buisman, C.J.N. Granular sludge formation and characterization in a chain elongation process. Process Biochem. 2016, 51, 1594–1598. [Google Scholar] [CrossRef]
- Wu, Q.; Feng, X.; Guo, W.; Bao, X.; Ren, N. Long-term medium chain carboxylic acids production from liquor-making wastewater: Parameters optimization and toxicity mitigation. Chem. Eng. J. 2020, 388, 124218. [Google Scholar] [CrossRef]
- Sudmalis, D.; Gagliano, M.C.; Pei, R.; Grolle, K.; Plugge, C.M.; Rijnaarts, H.H.M.; Zeeman, G.; Temmink, H. Fast anaerobic sludge granulation at elevated salinity. Water Res. 2018, 128, 293–303. [Google Scholar] [CrossRef]
- Gagliano, M.C.; Sudmalis, D.; Pei, R.; Temmink, H.; Plugge, C.M. Microbial Community Drivers in Anaerobic Granulation at High Salinity. Front. Microbiol. 2020, 11, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Li, X.; Wang, Y.; Tao, Y.; Lu, S.; Zhu, X.; Li, D. Improvement of n-caproic acid production with Ruminococcaceae bacterium CPB6: Selection of electron acceptors and carbon sources and optimization of the culture medium. Microb. Cell Fact. 2018, 17, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Spirito, C.M.; Marzilli, A.M.; Angenent, L.T. Higher Substrate Ratios of Ethanol to Acetate Steered Chain Elongation toward n-Caprylate in a Bioreactor with Product Extraction. Environ. Sci. Technol. 2018, 52, 13438–13447. [Google Scholar] [CrossRef]
- Roghair, M.; Liu, Y.; Strik, D.P.B.T.B.; Weusthuis, R.A.; Bruins, M.E.; Buisman, C.J.N. Development of an Effective Chain Elongation Process from Acidified Food Waste and Ethanol Into n-Caproate. Front. Bioeng. Biotechnol. 2018, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kleerebezem, R.; Van Loosdrecht, M.C.M. A generalized method for thermodynamic state analysis of environmental systems. Crit. Rev. Environ. Sci. Technol. 2010, 40, 1–54. [Google Scholar] [CrossRef]
- Takahashi, S.; Tomita, J.; Nishioka, K.; Hisada, T.; Nishijima, M. Development of a Prokaryotic Universal Primer for Simultaneous Analysis of Bacteria and Archea Using Next-Generation Sequencing. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [Green Version]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Glo, F.O.; Yarza, P. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef]
- Balcome, S.; Carlson, M. InteractiveDisplay: Package for Enabling Powerful Shiny Web Displays of Bioconductor Objects. 2020. Available online: http://bioconductor.org/packages/release/bioc/html/interactiveDisplay.html (accessed on 19 January 2021).
- Paulson, J.N.; Stine, O.C.; Bravo, H.C.; Pop, M. Differential abundance analysis for microbial marker-gene surveys. Nat. Methods 2013, 10, 1200–1202. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. 2019. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 19 January 2021).
- Beck, M.W.; Mikryukov, V. ggord: Ordination Plots with ggplot2. 2020. Available online: https://fawda123.github.io/ggord/ (accessed on 19 January 2021).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2008; ISBN 978-0-387-78170-9. [Google Scholar]
- Zhu, X.; Zhou, Y.; Wang, Y.; Wu, T.; Li, X.; Li, D.; Tao, Y. Production of high-concentration n-caproic acid from lactate through fermentation using a newly isolated Ruminococcaceae bacterium CPB6. Biotechnol. Biofuels 2017, 10, 1–12. [Google Scholar] [CrossRef]
- Scarborough, M.J.; Lawson, C.E.; Hamilton, J.J.; Donohue, T.J.; Noguera, D.R. Metatranscriptomic and Thermodynamic Insights into Medium-Chain Fatty Acid Production Using an Anaerobic Microbiome. mSystems 2018, 3, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Zagrodnik, R.; Duber, A.; Łȩżyk, M.; Oleskowicz-Popiel, P. Enrichment Versus Bioaugmentation—Microbiological Production of Caproate from Mixed Carbon Sources by Mixed Bacterial Culture and Clostridium kluyveri. Environ. Sci. Technol. 2020, 54, 5864–5873. [Google Scholar] [CrossRef]
- Lee, J.; Jang, Y.; Han, M.; Kim, Y.; Lee, Y. Deciphering Clostridium tyrobutyricum Metabolism Based on the Whole-Genome Sequence and Proteome Analyses. MBio 2016, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Ingham, S.C.; Hassler, J.R.; Tsai, Y.; Ingham, B.H. Differentiation of lactate-fermenting, gas-producing Clostridium spp. isolated from milk. Int. J. Food Microbiol. 1998, 43, 173–183. [Google Scholar] [CrossRef]
- Weimer, P.J.; Moen, G.N. Quantitative analysis of growth and volatile fatty acid production by the anaerobic ruminal bacterium Megasphaera elsdenii T81. Appl. Microbiol. Biotechnol. 2013, 97, 4075–4081. [Google Scholar] [CrossRef]
- Diez-Gonzalez, F.; Russell, J.B.; Hunter, J.B. The role of an NAD-independent lactate dehydrogenase and acetate in the utilization of lactate by Clostridium acetobutylicum strain P262. Arch. Microbiol. 1995, 164, 36–42. [Google Scholar] [CrossRef]
- Tomlinson, N.; Barker, H.A. Carbon dioxide and acetate utilization by clostridium kluyveri. I. Influence of nutritional conditions on utilization patterns. J. Biol. Chem. 1954, 209, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Schink, B. Energetics of syntrophic cooperation in methanogenic degradation. Microbiol. Mol. Biol. Rev. 1997, 61, 262–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nzeteu, C.O.; Trego, A.C.; Abram, F.; Flaherty, V.O. Reproducible, high—Yielding, biological caproate production from food waste using a single-phase anaerobic reactor system. Biotechnol. Biofuels 2018, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Godwin, S.; Kang, A.; Gulino, L.-M.; Manefield, M.; Gutierrez-Zamora, M.-L.; Kienzle, M.; Ouwerkerk, D.; Dawson, K.; Klieve, A.V. Investigation of the microbial metabolism of carbon dioxide and hydrogen in the kangaroo foregut by stable isotope probing. ISME J. 2014, 8, 1855–1865. [Google Scholar] [CrossRef] [PubMed]
- Poehlein, A.; Bremekamp, R.; Lutz, V.T.; Schulz, L.M.; Daniel, R. Draft Genome Sequence of the Butanoic Acid-Producing Bacterium Clostridium luticellarii DSM 29923, Used for Strong Aromatic Chinese Liquor Production. Genome Announc. 2018, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weghoff, M.C.; Bertsch, J.; Müller, V. A novel mode of lactate metabolism in strictly anaerobic bacteria. Environ. Microbiol. 2015, 17, 670–677. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phase | Days | Supply | Flow Rate | SGV | HRT | ULV |
|---|---|---|---|---|---|---|
| [mL∙min−1] | [m∙h−1] | [d] | [m∙h−1] | |||
| 1 | 62.7–85.7 | Continuous gas | 12 | 0.22 | 2 | 3 |
| 2 | 87.7–100.8 | Continuous gas | 120 | 2.2 | 2 | 3 |
| 3 | 104.8–149.9 | Intermittent gas | 11.3 | 3.6 | 2 | 3 |
| 4 | 154–188.8 | Intermittent gas | 11.3 | 3.6 | 1 | 3 |
| 5 | 190.7–231.9 | No gas | 0 | 0 | 1 | 9 |
| 6 | 232.7–237.7 | Propionate, no gas | 0 | 0 | 1 | 9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Contreras-Dávila, C.A.; Ali, A.; Buisman, C.J.N.; Strik, D.P.B.T.B. Lactate Metabolism and Microbiome Composition Are Affected by Nitrogen Gas Supply in Continuous Lactate-Based Chain Elongation. Fermentation 2021, 7, 41. https://doi.org/10.3390/fermentation7010041
Contreras-Dávila CA, Ali A, Buisman CJN, Strik DPBTB. Lactate Metabolism and Microbiome Composition Are Affected by Nitrogen Gas Supply in Continuous Lactate-Based Chain Elongation. Fermentation. 2021; 7(1):41. https://doi.org/10.3390/fermentation7010041
Chicago/Turabian StyleContreras-Dávila, Carlos A., Arielle Ali, Cees J. N. Buisman, and David P. B. T. B. Strik. 2021. "Lactate Metabolism and Microbiome Composition Are Affected by Nitrogen Gas Supply in Continuous Lactate-Based Chain Elongation" Fermentation 7, no. 1: 41. https://doi.org/10.3390/fermentation7010041
APA StyleContreras-Dávila, C. A., Ali, A., Buisman, C. J. N., & Strik, D. P. B. T. B. (2021). Lactate Metabolism and Microbiome Composition Are Affected by Nitrogen Gas Supply in Continuous Lactate-Based Chain Elongation. Fermentation, 7(1), 41. https://doi.org/10.3390/fermentation7010041