
Unravelling the Impact of Grape Washing, SO2, and Multi-Starter Inoculation in Lab-Scale Vinification Trials of Withered Black Grapes

 ,
, 
Abstract
:1. Introduction
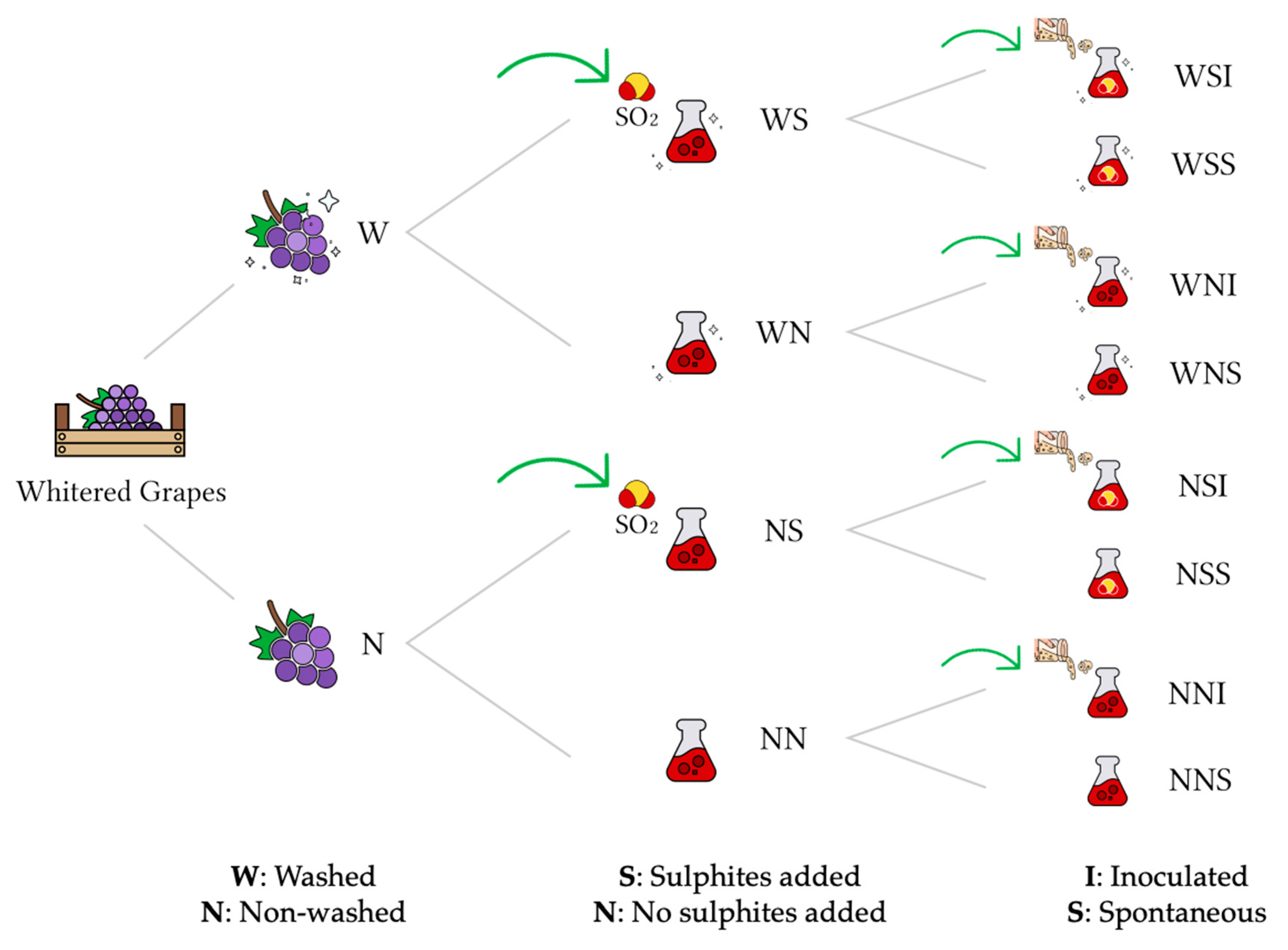
2. Materials and Methods
2.1. Withered Grapes
2.2. Grape Washing
2.3. Microvinifications
2.4. Fermentation Kinetics and Microbiological Analysis
2.5. Chemical Analysis
2.6. DNA Isolation
2.7. Molecular Analysis
2.8. Statistical Analysis
3. Results and Discussion
3.1. Impact of Washing Treatment on Grape Microbiota and Metals Content
3.2. Fermentation Performance
3.3. Population Dynamics
3.4. Chemical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guerrini, L.; Masella, P.; Angeloni, G.; Calamai, L.; Spinelli, S.; Di Biasi, S.; Parenti, A. Harvest of Sangiovese grapes: The influence of material other than grape and unripe berries on wine quality. Eur. Food Res. Technol. 2018, 244, 1487–1496. [Google Scholar] [CrossRef]
- Salvetti, E.; Campanaro, S.; Campedelli, I.; Fracchetti, F.; Gobbi, A.; Tornielli, G.B.; Torriani, S.; Felis, G. Whole-metagenome-sequencing-based community profiles of Vitis vinifera L. cv. Corvina berries withered in two post-harvest conditions. Front. Microbiol. 2016, 7, 937. [Google Scholar] [CrossRef] [Green Version]
- Stefanini, I.; Carlin, S.; Tocci, N.; Albanese, D.; Donati, C.; Franceschi, P.; Paris, M.; Zenato, A.; Tempesta, S.; Bronzato, A.; et al. Core microbiota and metabolome of Vitis vinifera L. cv. Corvina grapes and musts. Front. Microbiol. 2017, 8, 457. [Google Scholar] [CrossRef] [PubMed]
- Lorenzini, M.; Zapparoli, G. Yeast-like fungi and yeasts in withered grape carposphere: Characterization of Aureobasidium pullulans population and species diversity. Int. J. Food Microbiol. 2019, 289, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Slaghenaufi, D.; Boscaini, A.; Prandi, A.; Dal Cin, A.; Zandonà, V.; Luzzini, G.; Ugliano, M. Influence of different modalities of grape withering on volatile compounds of young and aged Corvina wines. Molecules 2020, 25, 2141. [Google Scholar] [CrossRef]
- Cisterna, B.; Boschi, F.; Croce, A.C.; Podda, R.; Zanzoni, S.; Degl’Innocenti, D.; Bernardi, P.; Costanzo, M.; Marzola, P.; Covi, V.; et al. Ozone treatment of grapes during withering for Amarone wine: A multimodal imaging and spectroscopic analysis. Microsc. Microanal. 2018, 24, 564–573. [Google Scholar] [CrossRef] [PubMed]
- Cavazza, A.; Franciosi, E.; Pojer, M.; Mattivi, F. Washing the grapes before crushing: Effects on contaminants and fermentation. Bull. L’OIV 2007, 80, 483–496. [Google Scholar]
- Bertoldi, D.; Nicolini, G.; Larcher, R.; Bertamini, M.; Concheri, G. Washing methods for removal of residues of 44 mineral elements from grapes. In Proceedings of the 5th International Specialized Conference on Sustainable Viticulture: Winery Waste and Ecologic Impacts Management, Trento-Verona, Italy, 30 March–3 April 2009; pp. 425–428. [Google Scholar]
- Vaquero-Fernández, L.; Sanz-Asensio, J.; Fernández-Zurbano, P.; López-Alonso, M.; Martínez-Soria, M.T. Determination of fungicide pyrimethanil in grapes, must, fermenting must and wine. J. Sci. Food Agric. 2013, 93, 1960–1966. [Google Scholar] [CrossRef]
- Agarbati, A.; Canonico, L.; Ciani, M.; Comitini, F. The impact of fungicide treatments on yeast biota of Verdicchio and Montepulciano grape varieties. PLoS ONE 2019, 14, e0217385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Wang, R.; Zhan, J.; Huang, W. High levels of copper retard the growth of Saccharomyces cerevisiae by altering cellular morphology and reducing its potential for ethanolic fermentation. Int. J. Food Sci. Technol. 2020. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Thorngate, J.H.; Richardson, P.M.; Mills, D.A. Microbial biogeography of wine grapes is conditioned by cultivar, vintage, and climate. Proc. Natl. Acad. Sci. USA 2014, 111, E139–E148. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Zhang, P.; Chen, D.; Howell, K.S. From the vineyard to the winery: How microbial ecology drives regional distinctiveness of wine. Front. Microbiol. 2019, 10, 2679. [Google Scholar] [CrossRef] [PubMed]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past and future of non-Saccharomyces yeasts: From spoilage microorganisms to biotechnological tools for improving wine aroma complexity. Front. Microbiol. 2016, 7, 411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benito, Á.; Calder, F.; Benito, S. The influence of non-Saccharomyces species on wine fermentation quality parameters. Fermentation 2019, 5, 54. [Google Scholar] [CrossRef] [Green Version]
- Capozzi, V.; Garofalo, C.; Chiriatti, M.A.; Grieco, F.; Spano, G. Microbial terroir and food innovation: The case of yeast biodiversity in wine. Microbiol. Res. 2015, 181, 75–83. [Google Scholar] [CrossRef]
- Romano, P.; Capece, A. La fermentazione spontanea. In Microbiologia Enologica, 2nd ed.; Suzzi, G., Tofalo, R., Eds.; Edagricole—Edizioni Agricole di New Business Media srl: Milano, Italy, 2018; pp. 1–13. [Google Scholar]
- Pretorius, I.S. Tasting the terroir of wine yeast innovation. FEMS Yeast Res. 2020, 20, foz084. [Google Scholar] [CrossRef] [Green Version]
- Comitini, F.; Capece, A.; Ciani, M.; Romano, P. New insights on the use of wine yeasts. Curr. Opin. Food Sci. 2017, 13, 44–49. [Google Scholar] [CrossRef]
- Nardi, T. Microbial resources as a tool for enhancing sustainability in winemaking. Microorganisms 2020, 8, 507. [Google Scholar] [CrossRef] [Green Version]
- Lisanti, M.T.; Blaiotta, G.; Nioi, C.; Moio, L. Alternative methods to SO2 for microbiological stabilization of wine. Compr. Rev. Food Sci. Food Saf. 2019, 18, 455–479. [Google Scholar] [CrossRef] [Green Version]
- Capitello, R.; Sirieix, L. Consumers’ perceptions of sustainable wine: An exploratory study in France and Italy. Economies 2019, 7, 33. [Google Scholar] [CrossRef] [Green Version]
- Binati, R.L.; Innocente, G.; Gatto, V.; Celebrin, A.; Polo, M.; Felis, G.E.; Torriani, S. Exploring the diversity of a collection of native non-Saccharomyces yeasts to develop co-starter cultures for winemaking. Food Res. Int. 2019, 12, 432–442. [Google Scholar] [CrossRef]
- Binati, R.L.; Lemos Junior, W.J.F.; Luzzini, G.; Slaghenaufi, D.; Ugliano, M.; Torriani, S. Contribution of non-Saccharomyces yeasts to wine volatile and sensory diversity: A study on Lachancea thermotolerans, Metschnikowia spp. and Starmerella bacillaris strains isolated in Italy. Int. J. Food Microbiol. 2020, 318, 108470. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Ohta, M.; Richardson, P.M.; Mills, D.A. Monitoring seasonal changes in winery resident microbiota. PLoS ONE 2013, 8, e66437. [Google Scholar] [CrossRef] [PubMed]
- Cocolin, L.; Bisson, L.F.; Mills, D.A. Direct profiling of the dynamics in wine fermentations. FEMS Microbiol. Lett. 2000, 189, 81–87. [Google Scholar] [CrossRef]
- Pfliegler, W.P.; Horváth, E.; Kállai, Z.; Sipiczki, M. Diversity of Candida zemplinina isolates inferred from RAPD, micro/minisatellite and physiological analysis. Microbiol. Res. 2014, 169, 402–410. [Google Scholar] [CrossRef] [PubMed]
- Muyzer, G.; de Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes encoding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenzini, M.; Azzolini, M.; Tosi, E.; Zapparoli, G. Postharvest grape infection of Botrytis cinerea and its interactions with other molds under withering conditions to produce noble-rotten grapes. J. Appl. Microbiol. 2013, 114, 762–770. [Google Scholar] [CrossRef]
- Prakitchaiwattana, C.J.; Fleet, G.H.; Heard, G.M. Application and evaluation of denaturing gradient gel electrophoresis to analyze the yeast ecology of wine grapes. FEMS Yeast Res. 2004, 4, 865–877. [Google Scholar] [CrossRef] [Green Version]
- Norocel, L.; Gutt, G. Study of the evolution of micro- and macroelements during the winemaking stages: The importance of copper and iron quantification. Food Environ. Saf. 2017, 16, 5–12. [Google Scholar]
- Claus, H. How to deal with uninvited guests in wine: Copper and copper-containing oxidases. Fermentation 2020, 6, 38. [Google Scholar] [CrossRef] [Green Version]
- Durguti, V.; Aliu, S.; Laha, F.; Feka, F. Determination of iron, copper and zinc in the wine by FAAS. Emerg. Sci. J. 2020, 4, 411–417. [Google Scholar] [CrossRef]
- Brandolini, V.; Tedeschi, P.; Capece, A.; Maietti, A.; Mazzotta, D.; Salzano, G.; Paparella, A.; Romano, P. Saccharomyces cerevisiae wine strains differing in copper resistance exhibit different capability to reduce copper content in wine. World J. Microbiol. Biotechnol. 2002, 18, 499–503. [Google Scholar] [CrossRef]
- Ferreira, J.; Du Toit, M.; Du Toit, W.J. The effects of copper and high sugar concentrations on growth, fermentation efficiency and volatile acidity production of different commercial wine yeast strains. Aust. J. Grape Wine Res. 2006, 12, 50–56. [Google Scholar] [CrossRef]
- Cavazza, A.; Guzzon, R.; Malacarne, M.; Larcher, R. The influence of the copper content in grape must on alcoholic fermentation kinetics and wine quality: A survey on the performance of 50 commercial active dry yeasts. Vitis 2013, 52, 149–155. [Google Scholar]
- Sun, X.; Liu, L.; Ma, T.; Yu, J.; Huang, W.; Fang, Y.; Zhan, J. Effect of high Cu2+ stress on fermentation performance and copper biosorption of Saccharomyces cerevisiae during wine fermentation. Food Sci. Technol. 2019, 39, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.Y.; Liu, L.; Zhao, Y.; Ma, T.; Zhao, F.; Huang, W.; Zhan, J. Effect of copper stress on growth characteristics and fermentation properties of Saccharomyces cerevisiae and the pathway of copper adsorption during wine fermentation. Food Chem. 2016, 192, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Regueiro, J.; López-Fernández, O.; Rial-Otero, R.; Cancho-Grande, B.; Simal-Gándara, J. A review on the fermentation of foods and the residues of pesticides-biotransformation of pesticides and effects on fermentation and food quality. Crit. Rev. Food Sci. Nutr. 2015, 55, 839–863. [Google Scholar] [CrossRef] [PubMed]
- Morata, A.; Loira, I.; Escott, C.; Del Fresno, J.M.; Bañuelos, M.A.; Suàrez-Lepe, J.A. Applications of Metschnikowia pulcherrima in wine biotechnology. Fermentation 2019, 5, 63. [Google Scholar] [CrossRef] [Green Version]
- Tesnière, C.; Brice, C.; Blondin, B. Responses of Saccharomyces cerevisiae to nitrogen starvation in wine alcoholic fermentation. Appl. Microbiol. Biotechnol. 2015, 99, 7025–7034. [Google Scholar] [CrossRef]
- Kessi-Pérez, E.I.; Molinet, J.; Martínez, C. Disentangling the genetic bases of Saccharomyces cerevisiae nitrogen consumption and adaptation to low nitrogen environments in wine fermentation. Biol. Res. 2020, 53, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bisson, L.F. Stuck and sluggish fermentations. Am. J. Enol. Vitic. 1999, 50, 107–119. [Google Scholar]
- Medina, K.; Boido, E.; Dellacassa, E.; Carrau, F. Growth of non-Saccharomyces yeasts affects nutrient availability for Saccharomyces cerevisiae during wine fermentation. Int. J. Food Microbiol. 2012, 157, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Taillandier, P.; Lai, Q.P.; Julien-Ortiz, A.; Brandam, C. Interactions between Torulaspora delbrueckii and Saccharomyces cerevisiae in wine fermentation: Influence of inoculation and nitrogen content. World J. Microbiol. Biotechnol. 2014, 30, 1959–1967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemsawasd, V.; Viana, T.; Ardö, Y.; Arneborg, N. Influence of nitrogen sources on growth and fermentation performance of different wine yeast species during alcoholic fermentation. Appl. Microbiol. Biotechnol. 2015, 99, 10191–10207. [Google Scholar] [CrossRef]
- Andorrà, I.; Berradre, M.; Rozès, N.; Mas, A.; Guillamón, J.M.; Esteve-Zarzoso, B. Effect of pure and mixed cultures of the main wine yeast species on grape must fermentations. Eur. Food Res. Technol. 2010, 231, 215–224. [Google Scholar] [CrossRef] [Green Version]
- Medina, K.; Boido, E.; Fariña, L.; Gioia, O.; Gomez, M.E.; Barquet, M.; Gaggero, C.; Dellacassa, E.; Carrau, F. Increased flavour diversity of Chardonnay wines by spontaneous fermentation and co-fermentation with Hanseniaspora vinae. Food Chem. 2013, 141, 2513–2521. [Google Scholar] [CrossRef]
- <named-content content-type="background:white">Liu, S.-Q.; Pilone, G.J. An overwiew of formation and roles of acetaldehyde in winemaking with emphasis on microbiological implications. Int. J. Food Sci. Technol. 2000, 35, 49–61. [Google Scholar]
- Li, E.; Mira de Orduña, R. Acetaldehyde kinetics of enological yeast during alcoholic fermentation in grape must. J. Ind. Microbiol. Biotechnol. 2017, 44, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Mira de Orduña, R.; Lamon, J. Acetaldehyde Management during Winemaking. Available online: https://www.winemak-in.com/en/publications/acetaldehyde-management-during-winemaking (accessed on 22 November 2020).
- Danilewicz, J.C. Review of reaction mechanisms of oxygen and proposed intermediate reaction products in wine: Central role of iron and copper. Am. J. Enol. Vitic. 2003, 54, 73–85. [Google Scholar]
- Jackowetz, J.N.; Dierschke, S.E.; Mira de Orduña, R. Multifactorial analysis of acetaldehyde kinetics during alcoholic fermentation by Saccharomyces cerevisiae. Food Res. Int. 2011, 44, 310–316. [Google Scholar] [CrossRef]
- Ochando, T.; Mouret, J.R.; Humbert-Goffard, A.; Aguera, E.; Sablayrolles, J.M.; Farines, V. Comprehensive study of the dynamic interaction between SO2 and acetaldehyde during alcoholic fermentation. Food Res. Int. 2020, 136, 109607. [Google Scholar] [CrossRef] [PubMed]
- Herraiz, T.; Martin-Alvarez, P.J.; Reglero, G.; Herraiz, M.; Cabezudo, M.D. Differences between wines fermented with and without sulphur dioxide using various selected yeasts. J. Sci. Food Agric. 1989, 49, 249–258. [Google Scholar] [CrossRef]
- Zea, L.; Serratosa, M.P.; Mérida, J.; Moyano, L. Acetaldehyde as key compound for the authenticity of sherry wines: A study covering 5 decades. Compr. Rev. Food Sci. Food Saf. 2015, 14, 681–693. [Google Scholar] [CrossRef]
- Sadoudi, M.; Rousseaux, S.; David, V.; Alexandre, H. Metschnikowia pulcherrima Influences the Expression of Genes Involved in PDH Bypass and Glyceropyruvic Fermentation in Saccharomyces cerevisiae. Front. Microbiol. 2017, 8, 1137. [Google Scholar] [CrossRef]
- Hranilovic, A.; Gambetta, J.M.; Jeffery, D.W.; Grbin, P.R.; Jiranek, V. Lower-alcohol wines produced by Metschnikowia pulcherrima and Saccharomyces cerevisiae co-fermentations: The effect of sequential inoculation timing. Int. J. Food Microbiol. 2020, 329, 108651. [Google Scholar] [CrossRef]
- Seguinot, P.; Ortiz-Julien, A.; Camarasa, C. Impact of nutrient availability on the fermentation and production of aroma compounds under sequential inoculation with M. pulcherrima and S. cerevisiae. Front. Microbiol. 2020, 11, 305. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Residual Sugar (g/L) | CO2 Middle (g/L) † | YAN (mg/L) ‡ | Free SO2 (mg/L) | Total SO2 (mg/L) | Acetaldehyde (mg/L) | Acetic Acid (g/L) |
|---|---|---|---|---|---|---|---|
| WSI | 44.36 ± 2.49 a | 94.74 ± 0.27 a | 83.20 ± 5.80 a | 8.85 ± 1.20 a | 66.00 ± 2.83 a | 25.65 ± 0.49 ab | 0.74 ± 0.13 b |
| WSS | 34.09 ± 4.38 a | 92.00 ± 0.96 a | 76.05 ± 10.68 ab | 7.65 ± 0.64 a | 49.00 ± 2.83 ab | 27.00 ± 1.41 a | 1.33 ± 0.04 a |
| WNI | 53.02 ± 8.83 a | 97.07 ± 4.44 a | 81.70 ± 1.27 a | 9.50 ± 2.69 a | 62.00 ± 8.49 ab | 25.16 ± 0.54 ab | 0.73 ± 0.03 b |
| WNS | 32.62 ± 6.57 a | 88.02 ± 1.05 a | 62.70 ± 3.82 b | 7.70 ± 1.27 a | 64.50 ± 2.12 ab | 28.51 ± 2.21 a | 1.09 ± 0.12 a |
| NSI | 38.02 ± 8.46 a | 87.11 ± 1.93 a | 75.35 ± 1.06 ab | 11.30 ± 0.00 a | 50.00 ± 8.49 ab | 26.50 ± 0.71 a | 0.78 ± 0.04 b |
| NSS | 34.00 ± 20.40 a | 72.20 ± 1.33 b | 71.00 ± 0.99 ab | 6.95 ± 0.49 a | 46.50 ± 3.54 b | 24.50 ± 0.71 ab | 1.16 ± 0.01 a |
| NNI | 42.91 ± 6.69 a | 94.78 ± 2.27 a | 87.85 ± 4.03 a | 11.00 ± 0.14 a | 50.50 ± 2.12 ab | 21.50 ± 0.71 b | 0.82 ± 0.01 b |
| NNS | 49.36 ± 2.72 a | 68.69 ± 7.19 b | 71.90 ± 1.41 ab | 6.90 ± 1.27 a | 46.50 ± 2.12 b | 24.00 ± 1.41 ab | 1.21 ± 0.02 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Checchia, I.; Binati, R.L.; Troiano, E.; Ugliano, M.; Felis, G.E.; Torriani, S. Unravelling the Impact of Grape Washing, SO2, and Multi-Starter Inoculation in Lab-Scale Vinification Trials of Withered Black Grapes. Fermentation 2021, 7, 43. https://doi.org/10.3390/fermentation7010043
Checchia I, Binati RL, Troiano E, Ugliano M, Felis GE, Torriani S. Unravelling the Impact of Grape Washing, SO2, and Multi-Starter Inoculation in Lab-Scale Vinification Trials of Withered Black Grapes. Fermentation. 2021; 7(1):43. https://doi.org/10.3390/fermentation7010043
Chicago/Turabian StyleChecchia, Ilaria, Renato L. Binati, Eleonora Troiano, Maurizio Ugliano, Giovanna E. Felis, and Sandra Torriani. 2021. "Unravelling the Impact of Grape Washing, SO2, and Multi-Starter Inoculation in Lab-Scale Vinification Trials of Withered Black Grapes" Fermentation 7, no. 1: 43. https://doi.org/10.3390/fermentation7010043
APA StyleChecchia, I., Binati, R. L., Troiano, E., Ugliano, M., Felis, G. E., & Torriani, S. (2021). Unravelling the Impact of Grape Washing, SO2, and Multi-Starter Inoculation in Lab-Scale Vinification Trials of Withered Black Grapes. Fermentation, 7(1), 43. https://doi.org/10.3390/fermentation7010043