
Bacteriocinogenic Bacillus spp. Isolated from Korean Fermented Cabbage (Kimchi)—Beneficial or Hazardous?

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Antagonistic Strains and Evaluation of Antimicrobial Activity
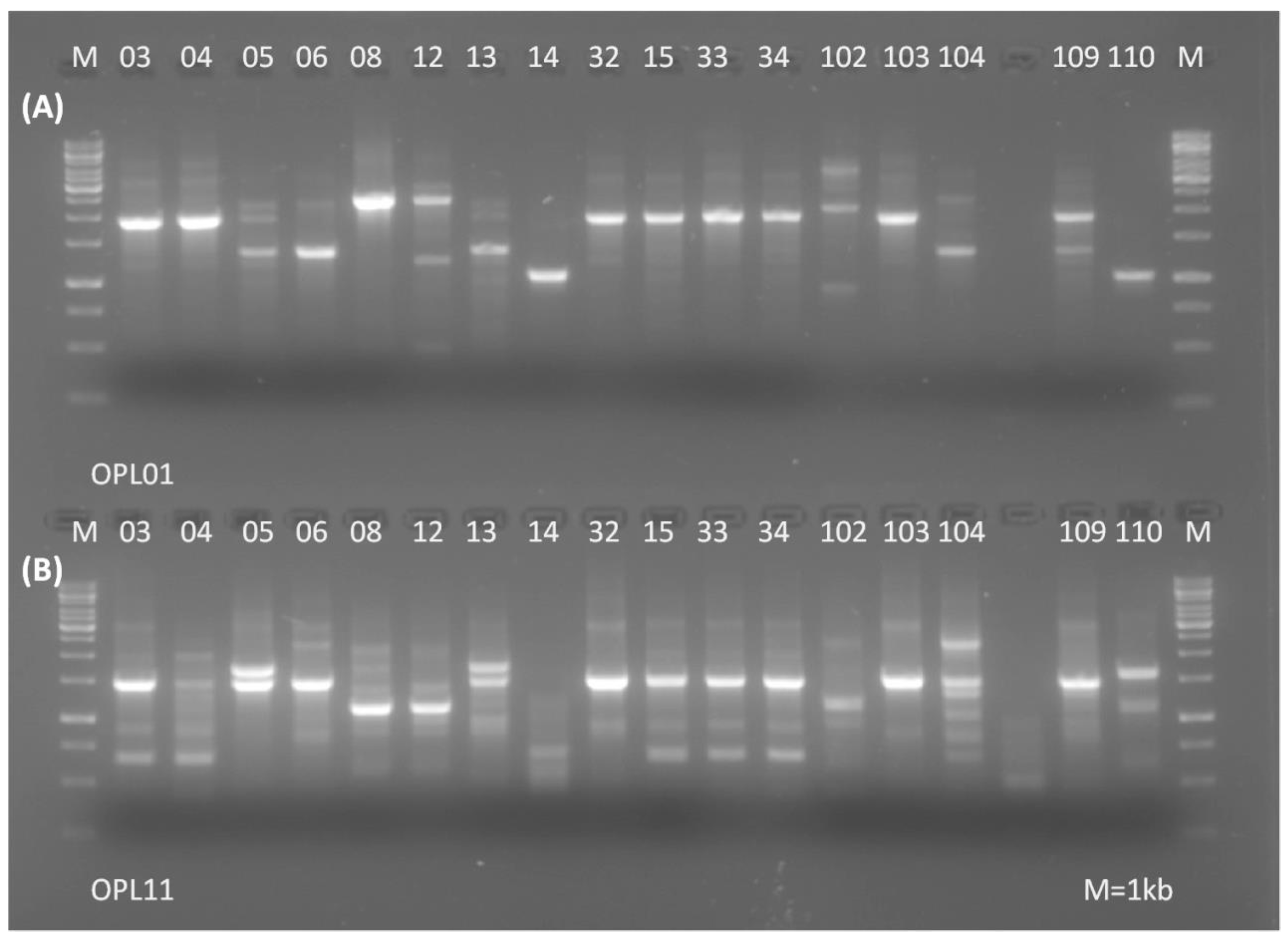
2.2. Differentiation and Identification of Selected Isolates
2.3. Sugar Fermentation Profile (API50CHB) and Production of Enzymes (APIZym)
2.4. Detection of Bacteriocin Genes and Other Antimicrobial Genes
2.5. Determination of Antimicrobial Activity and Nature of Antimicrobial Agent(s)
2.6. Stability of the Antimicrobial Substance(s)—pH, Temperature, and Chemicals
2.7. Screening of Virulence Genes
2.8. Virulence Activity—Physiological Approach
2.9. Antimicrobial Susceptibility Profiling
2.10. Detection of Vancomycin Resistant Genes
3. Results
3.1. Differentiation and Identification
3.2. Sugar Fermentation Profile (API50CHB) and Enzymes Production (APIZym)
3.3. Detection of Bacteriocin and Other Antimicrobial Genes
3.4. Stability of the Antimicrobial Substances—Effect of pH, Temperature, and Chemicals
3.5. In Vitro Hemolytic Activity and Presence of Genes Related to Hemolysin and Non-Hemolytic Enterotoxin Virulence Factors; Gelatinase and Biogenic Amine Production
3.6. Antibiotic Resistance
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ouoba, L.I.; Diawara, B.; Amoa-Awua, W.K.; Traoré, A.S.; Møller, P.L. Genotyping of starter cultures of Bacillus subtilis and Bacillus pumilus for fermentation of African locust bean (Parkia biglobosa) to produce Soumbala. Int. J. Food Microbiol. 2004, 90, 197–205. [Google Scholar] [CrossRef]
- Aliakbarpour, H.R.; Chamani, M.; Rahimi, G.; Sadeghi, A.A.; Qujeq, D. The Bacillus subtilis and lactic acid bacteria probiotics influences intestinal mucin gene expression, histomorphology and growth performance in broilers. Asian Australas. J. Anim. Sci. 2012, 25, 1285–1293. [Google Scholar] [CrossRef] [Green Version]
- Tamang, J.P.; Watanabe, K.; Holzapfel, W.H. Diversity of microorganisms in global fermented foods and beverages. A review. Front. Microbiol. 2016, 7, 377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Encyclopedia Britannica. 2008. Available online: https://www.britannica.com/ (accessed on 25 March 2020).
- Jung, J.Y.; Lee, S.H.; Kim, J.M.; Park, M.S.; Bae, J.-W.; Hahn, Y.; Madsen, E.L.; Jeon, C.O. Metagenomic analysis of kimchi, a traditional Korean fermented food. Appl. Environ. Microbiol. 2011, 77, 2264–2274. [Google Scholar] [CrossRef] [Green Version]
- Chikindas, M.L.; Weeks, R.; Drider, D.; Chistyakov, V.A.; Dicks, L.M. Functions and emerging applications of bacteriocins. Curr. Opin. Biotechnol. 2018, 49, 23–28. [Google Scholar] [CrossRef]
- Todorov, S.D.; Franco, B.D.G.M.; Tagg, J.R. Bacteriocins of Gram positive bacteria having activity spectra extending beyond closely-related species. Benef. Microbes 2019, 10, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Syamaladevi, R.M.; Tang, J.; Villa-Rojas, R.; Sablani, S.; Carter, B.; Campbell, G. Influence of water activity on thermal resistance of microorganisms in low-moistre foods: A review. Compr. Rev. Food Sci. Food Saf. 2016, 15, 353–370. [Google Scholar] [CrossRef] [Green Version]
- Bover-Cid, S.; Holzapfel, W.H. Improved screening procedure for biogenic amine production by lactic acid bacteria. Int. J. Food Microbiol. 1999, 53, 33–41. [Google Scholar] [CrossRef]
- Dos Anjos, T.R.; Cavicchioli, V.Q.; Lima, J.A.S.; Vasconcellos, A.N.; Vaz, A.C.N.; Rossi, G.A.M.; Campos-Galvão, M.E.M.; Todorov, S.D.; Mathias, L.A.; Schocken-Iturrino, R.P.; et al. Unsatisfactory microbiological aspects of UHT goat milk, soy milk and dairy beverage of goat milk and soy protein: A public health issue. Food Sci. Technol. 2019, 40, 349–354. [Google Scholar] [CrossRef] [Green Version]
- Colombo, M.; Nero, L.A.; Todorov, S.D. Safety profile of beneficial lactic acid bacteria isolated from dairy systems. Braz. J. Microbiol. 2020, 51, 787–795. [Google Scholar] [CrossRef]
- Perin, L.M.; Miranda, R.O.; Todorov, S.D.; Franco, B.D.G.M.; Nero, L.A. Virulence, antibiotic resistance and biogenic amines of bacteriocinogenic lactococci and enterococci isolated from goat milk. Int. J. Food Microbiol. 2014, 185, 121–126. [Google Scholar] [CrossRef]
- Priest, F.G. Extracellular enzyme synthesis in the genus Bacillus. Bacteriol. Rev. 1977, 41, 711–753. [Google Scholar] [CrossRef] [PubMed]
- Barg, H.; Malten, M.; Jahn, M.; Jahn, D. Protein and vitamin production in Bacillus megaterium. In Microbial Processes and Products. Methods in Biotechnology; Barredo, J.L., Ed.; Humana Press: Totowa, NJ, USA, 2005; Volume 18. [Google Scholar] [CrossRef]
- Stein, T. Bacillus subtilis antibiotics: Structures, syntheses and specific functions. Mol. Microbiol. 2005, 56, 845–857. [Google Scholar] [CrossRef] [PubMed]
- Latorre, J.D.; Hernandez-Velasco, X.; Wolfenden, R.E.; Vicente, J.L.; Wolfender, A.D.; Menconi, A.; Bielke, L.R.; Hargis, B.M.; Tellez, G. Evaluation and selection of Bacillus species based on enzyme production, antimicrobial activity, and biofilm synthesis as direct-fed microbial candidates for poultry. Front. Vet. Sci. 2016, 3, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daliri, E.B.; Oh, D.H.; Lee, B.H. Bioactive Peptides. Foods 2017, 6, 32. [Google Scholar] [CrossRef] [PubMed]
- Field, D.; Cotter, P.D.; Hill, C.; Ross, R.P. Bioengineering Lantibiotics for therapeutic success. Front. Microbiol. 2015, 6, 1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khatri, I.; Sharma, G.; Subramanian, S. Composite genome sequence of Bacillus clausii, a probiotic commercially available as Enterogermina®, and insights into its probiotic properties. BMC Microbiol. 2019, 19, 307. [Google Scholar] [CrossRef] [Green Version]
- Hongu, N.; Kim, A.S.; Suzuki, A.; Wilson, H.; Tsui, K.C.; Park, S. Korean kimchi: Promoting healthy meals through cultural tradition. J. Ethn. Foods 2016, 4, 172–180. [Google Scholar] [CrossRef]
- Spencer, R.C. Bacillus anthracis. J. Clin. Pathol. 2003, 56, 182–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messelhäußer, U.; Ehling-Schulz, M. Bacillus cereus—A multifaceted opportunistic pathogen. Curr. Clin. Microbiol. Rep. 2018, 5, 120–125. [Google Scholar] [CrossRef] [Green Version]
- Baccigalupi, L.; Ricca, E.; Ghelardi, E. Non-LAB probiotics: Spore formers. In Probiotics and Prebiotics: Current Research and Future Trends; Venema, K., do Carmo, A.P., Eds.; Caister Academic Press: Poole, UK, 2015; pp. 93–104. [Google Scholar]
- Elshaghabee, F.M.F.; Rokana, N.; Gulhane, R.D.; Sharma, C.; Panwar, H. Bacillus as potential probiotics: Status, concerns, and future perspectives. Front. Microbiol. 2017, 8, 1490. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.-W.; Yoon, S.-R.; Kim, S.-J.; Lee, H.M.; Lee, J.Y.; Lee, J.-H.; Kim, S.H.; Ha, J.-H. Identification of microbial communities, with a focus on foodborne pathogens, during kimchi manufacturing process using culture-independent and -dependent analyses. LWT Food Sci. Technol. 2016, 81, 153–159. [Google Scholar] [CrossRef]
- Senesi, S.; Celandroni, F.; Tavanti, A.; Ghelardi, E. Molecular characterization and identification of Bacillus clausii strains marketed for use in oral bacteriotherapy. Appl. Environ. Microbiol. 2001, 67, 834–839. [Google Scholar] [CrossRef] [Green Version]
- De Vos, P.; Garrity, G.M.; Jones, D.; Kreig, N.R.; Ludwig, W.; Rainey, F.A.; Schleifel, K.-H.; Whitman, W.B. Bergey’s Manual of Systematic Bacteriology; The Firmicutes; Wiley Publishing Group: Hoboken, NJ, USA, 2009; Volume 3. [Google Scholar] [CrossRef]
- De Moraes, G.M.D.; de Abreu, L.R.; do Egito, A.S.; Salles, H.O.; da Silva, L.M.F.; Nero, L.A.; Todorov, S.D.; dos Santos, K.M.O. Functional properties of Lactobacillus mucosae strains isolated from Brazilian goat milk. Probiotics Antimicrob. Proteins 2016, 9, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Chopra, L.; Singh, G.; Choudhary, V.; Sahoo, D.K. Sonorensin: An antimicrobial peptide, belonginig to the heterocycloantharicin subfamily of bacteriocins, from a new marine isolate, Bacillus sonorensis MT93. Appl. Environ. Microbiol. 2014, 80, 2981–2990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todorov, S.D.; Stojanovski, S.; Iliev, I.; Moncheva, P.; Nero, L.A.; Ivanova, I.V. Technology and safety assessment for lactic acid bacteria isolated from traditional Bulgarian fermented meat product “Lukanka”. Braz. J. Microbiol. 2017, 48, 576–586. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, K.M.O.; de Matos, C.R.; Salles, H.O.; Franco, B.D.G.M.; Arellano, K.; Holzapfel, W.H.; Todorov, S.D. Exploring beneficial/virulence properties of two dairy related strains of Streptococcus infantarius subsp. infantarius. Probiotics Antimicrob. Proteins 2020, 12, 1524–1541. [Google Scholar] [CrossRef]
- Todorov, S.D.; Cavicchioli, V.Q.; Ananieva, M.; Bivolarski, V.P.; Vasileva, T.A.; Hinkov, A.V.; Todorov, D.G.; Shishkov, S.; Haertlé, T.; Iliev, I.N.; et al. Expression of coagulin A with low cytotoxic activity by Pediococcus pentosaceus ST65ACC isolated from raw milk cheese. J. Appl. Microbiol. 2019, 128, 458–472. [Google Scholar] [CrossRef]
- Guinebretiere, M.-H.; Broussolle, V.; Nguyen-The, C. Enterotoxigenic profile of food-poisoning and food-borne Bacillus cereus strains. J. Clin. Microbiol. 2002, 40, 3053–3056. [Google Scholar] [CrossRef] [Green Version]
- EUCAST. Available online: https://www.eucast.org/ast_of_bacteria/ (accessed on 25 March 2020).
- Valledor, S.J.D.; Bucheli, J.E.V.; Holzapfel, W.H.; Todorov, S.D. Exploring beneficial properties of the bacteriocinogenic Enterococcus faecium ST10Bz strain isolated from boza, a Bulgarian cereal-based beverage. Microorganisms 2020, 8, 1474. [Google Scholar] [CrossRef]
- Ciffo, F. Determination of the spectrum of antibiotic resistance of the Bacillus subtilis strains of Enterogermina. Chemioterapia 1984, 3, 45–52. [Google Scholar]
- McAuliffe, O.; Ryan, M.P.; Ross, R.P.; Hill, C.; Breeuwer, P.; Abee, T. Lacticin 3147, a broad-spectrum bacteriocin which selectively dissipates the membrane potential. Appl. Environ. Microbiol. 1998, 64, 439–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shelburne, C.E.; An, F.Y.; Dholpe, V.; Ramamoorthy, A.; Lopatin, D.E.; Lantz, M.S. The spectrum of antimicrobial activity of the bacteriocin subtilosin A. J. Antimicrob. Chemother. 2006, 59, 297–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuda, Y.; Ono, H.; Kitagawa, H.; Ito, H.; Mu, F.; Sawa, N.; Zendo, T.; Sonomoto, K. Identification and characterization of leucocyclicin Q, a novel cyclic bacteriocin produced by Leuconostoc mesenteroides TK41401. Appl. Environ. Microbiol. 2011, 77, 8164–8170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Compaore, C.S.; Nielsen, D.S.; Sawadogo-Lingani, H.; Berner, T.S.; Nielsen, K.F.; Adimpong, D.B.; Diawara, B.; Ouedraogo, G.A.; Jakobsen, M.; Thorsen, L. Bacillus amyloliquefaciens subsp. plantarum strains as potential protective starter cultures for the production of Bikalga, an alkaline fermented food. J. Appl. Microbiol. 2013, 115, 133–146. [Google Scholar] [CrossRef]
- Anihouvi, V.B.; Kpoclou, E.Y.; Hounhouigan, J.D. Use of starter cultures of Bacillus and staphylococcus in the controlled fermentation of Lanhouin, a traditional fish-based condiment from West Africa. Afr. J. Microbiol. Res. 2012, 6, 4767–4774. [Google Scholar] [CrossRef]
- Chang, H.C. Healthy and safe Korean traditional fermented foods: Kimchi and chongkukjang. J. Ethn. Foods. 2018, 5, 161–166. [Google Scholar] [CrossRef]
- Katz, E.; Demain, A.L. The peptide antibiotics of Bacillus: Chemistry, biogenesis, and possible functions. Biotechnol. Rev. 1977, 41, 449–474. [Google Scholar] [CrossRef] [Green Version]
- Hosoi, T.; Kiuchi, K. Natto—A food made by fermenting cooked soybeanswith Bacillus subtilis (natto). In Fermented Functional Foods; Farnworth, E.R., Ed.; CRC Press: Boca Raton, FL, USA, 2003; pp. 227–251. [Google Scholar]
- Cho, M.S.; Jin, Y.J.; Kang, B.K.; Park, Y.K.; Kim, C.K.; Park, D.S. Understanding the ontogeny and succession of Bacillus velezensis and B. subtilis subsp. subtilis by focusing on kimchi fermentation. Sci. Rep. 2018, 8, 7045. [Google Scholar] [CrossRef] [Green Version]
- Jeon, H.H.; Jung, J.Y.; Chun, B.H.; Kim, M.D.; Baek, S.Y.; Moon, J.Y.; Yeo, S.H.; Jeon, C.O. Screening and characterization of potential Bacillus starter cultures for fermenting low-salt soybean paste (Doenjang). J. Microbiol. Biotechnol. 2016, 26, 666–674. [Google Scholar] [CrossRef] [Green Version]
- Montel, M.C.; Masson, F.; Talon, R. Bacterial role in flavor development. Meat Sci. 1998, 49, S111–S123. [Google Scholar] [CrossRef]
- El Mecherfi, K.-E.; Todorov, S.D.; de Albuquerque, M.C.; Denery-Papini, S.; Lupi, R.; Haertlé, T.; Franco, B.D.G.M.; Larré, C. Allergenicity of fermented foods: Emphasis on seeds protein based product. Foods 2020, 9, 792. [Google Scholar] [CrossRef]
- Park, H.; Lee, M.; Ji, Y.; Todorov, S.D.; Holzapfel, W.H. Safety evaluation and in vivostrain-specific functionality of Bacillus strains isolated from Korean traditional fermented foods. Probiotics Antimicrob. Proteins 2020, 13, 60–71. [Google Scholar] [CrossRef]
- George, J. Elevated serum β-glucuronidase reflects hepatic lysosomal fragility following toxic liver injury in rats. Biochem. Cell Biol. 2008, 86, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.J.; Chang, H.C. Characterization of bacteriocin-like substances produced by Bacillus subtilis MJP1. Korean J. Microbiol. Biotechnol. 2007, 35, 339–346. [Google Scholar]
- Wu, S.; Jia, S.; Sun, D.; Chen, M.; Chen, X.; Zhong, J.; Huan, L. Purification and characterization of two novel antimicrobial peptides subpeptinJM4-A and subpeptinJM4-B Produced by Bacillus subtilis JM4. Curr. Microbiol. 2005, 51, 292–296. [Google Scholar] [CrossRef]
- Salazar, F.; Ortiz, A.; Sansinenea, E. Characterisation of two novel bacteriocin-like substances produced by Bacillus amyloliquefaciens ELI149 with broad-spectrum antimicrobial activity. J. Glob. Antimicrob. Resist. 2017, 11, 177–182. [Google Scholar] [CrossRef]
- Entian, K.-D.; de Vos, W.M. Genetics of subtilin and nisin biosyntheses. Antonie Van Leeuwenhoek 1996, 69, 109–117. [Google Scholar] [CrossRef]
- Parisot, J.; Carey, S.; Breukink, E.; Chan, W.C.; Narbad, A.; Bonev, B. Molecular mechanism of target recognition by subtilin, a class I lanthionine antibiotic. Antimicrob. Agents Chemother. 2008, 52, 612–618. [Google Scholar] [CrossRef] [Green Version]
- Zheng, G.; Yan, L.Z.; Vederas, J.C.; Zuber, P. Genes of the sbo-alb locus of Bacillus subtilis are required for production of the antilisterial bacteriocin subtilosin. J. Bacteriol. 1999, 181, 7346–7355. [Google Scholar] [CrossRef] [Green Version]
- Nakano, M.M.; Zheng, G.; Zuber, P. Dual control of sbo-alb operon expression by the Spo0 and ResDE systems of signal transduction under anaerobic conditions in Bacillus subtilis. J. Bacteriol. 2000, 182, 3274–3277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, N.-K.; Kim, W.-S.; Paik, H.-D. Bacillus strains as human probiotic: Characterization, safety, microbiome, and probiotic carrier. Food Sci. Biotechnol. 2019, 28, 1297–1305. [Google Scholar] [CrossRef]
- Lee, H.; Kim, H.-Y. Lantibiotics, Class I Bacteriocins from the Genus Bacillus. J. Microbiol. Biotechnol. 2011, 21, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Le Marrec, C.; Hyronimus, B.; Bressollier, P.; Verneuil, B.; Urdaci, M.C. Biochemical and genetic characterization of coagulin, a new antilisterial bacteriocin in the pediocin family of bacteriocins, produced by Bacillus coagulans I(4). Appl. Environ. Microbiol. 2000, 66, 5213–5220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocins—A viable alternative to antibiotics? Nat. Rev. Microbiol. 2013, 11, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Joerger, R.D. Alternatives to antibiotics: Bacteriocins, antimicrobial peptides and bacteriophages. Poult. Sci. 2003, 82, 640–647. [Google Scholar] [CrossRef] [PubMed]
- Vesterlund, S.; Vankerckhoven, V.; Saxelin, M.; Goossens, H.; Salminen, S.; Ouwehand, A.C. Safety assessment of Lactobacillus strains: Presence of putative risk factors in faecal, blood and probiotic isolates. Int. J. Food Microbiol. 2007, 116, 325–331. [Google Scholar] [CrossRef]
- Barbosa, J.; Gibbs, P.A.; Teixeira, P. Virulence factors among enterococci isolated from traditional fermented meat products produced in the north of Portugal. Food Control 2010, 21, 651–656. [Google Scholar] [CrossRef]
- Franz, C.M.A.P.; Muscholl-Silberhorn, A.B.; Yousif, N.M.K.; Vancanneyt, M.; Swings, J.; Holzapfel, W.H. Incidence of virulence factors and antibiotic resistance among enterococci isolated from food. Appl. Environ. Microbiol. 2001, 67, 4385–4389. [Google Scholar] [CrossRef] [Green Version]
- Simon-Sarkadi, L.; Holzapfel, W.H.; Halász, A. Biogenic amine content and microbial contamination of leafy vegetables during storage at 5 °C. J. Food Biochem. 1994, 17, 407–418. [Google Scholar] [CrossRef]
- Özogul, Y.; Özogul, F. Chapter 1: Biogenic amines formation, toxicity, regulations in food. In Biogenic Amines in Food: Analysis, Occurrence and Toxicity; Royal Society of Chemistry: London, UK, 2019; pp. 1–17. [Google Scholar] [CrossRef]
- Halász, A.; Barath, A.; Simon-Sarkadi, L.; Holzapfel, W.H. Biogenic amines and their production by microorganisms in food. Trends Food Sci. Technol. 1994, 5, 42–49. [Google Scholar] [CrossRef]
- Halász, A.; Baráth, Ă.; Holzapfel, W.H. The influence of starter culture selection on sauerkraut fermentation. Z. Lebensm. Forsch. A 1999, 208, 434–438. [Google Scholar] [CrossRef]
- Barbieri, F.; Montanari, C.; Gardini, F.; Tabanelli, G. Biogenic amine production by lactic acid bacteria: A Review. Foods 2019, 8, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, M.H.S. Biogenic amines: Their importance in foods. Int. J. Food Microbiol. 1996, 29, 213–231. [Google Scholar] [CrossRef]
- Brink, B.T.; Damink, C.; Joosten, H.M.L.J.; Veld, J.H.I. Occurrence and formation of biologically active amines in foods. Int. J. Food Microbiol. 1990, 11, 73–84. [Google Scholar] [CrossRef]
- Atanasova, J.; Dalgalarrondo, M.; Iliev, I.; Moncheva, P.; Todorov, S.D.; Ivanova, I.V. Formation of free amino acids and bioactive peptides during the ripening of Bulgarian white brined cheeses. Probiotics Antimicrob. Proteins 2020, 13, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Stribny, J.; Gamero, A.; Pérez-Torrado, R.; Querol, A. Saccharomyces kudriavzevii and Saccharomyces uvarum differ from Saccharomyces cerevisiae during the production of aroma-active higher alcohols and acetate esters using their amino acidic precursors. Int. J. Food Microbiol. 2015, 205, 41–46. [Google Scholar] [CrossRef] [Green Version]
- Perea-Sanz, L.; Peris, D.; Belloch, C.; Flores, M. Debaryomyces hansenii metabolism of sulfur amino acids as precursors of volatile sulfur compounds of interest in meat products. J. Agric. Food Chem. 2019, 67, 9335–9343. [Google Scholar] [CrossRef]
- Mah, J.-H.; Kim, Y.J.; No, H.-K.; Hwang, H.-J. Determination of biogenic amines in kimchi, Korean traditional fermented vegetable products. Food Sci. Biotechnol. 2004, 13, 826–829. [Google Scholar]
- Tsai, Y.-H.; Kung, H.-F.; Lin, Q.-L.; Hwang, J.-H.; Cheng, S.-H.; Wei, C.-I.; Hwang, D.-F. Occurrence of histamine and histamine-forming bacteria in kimchi products in Taiwan. Food Chem. 2005, 90, 635–641. [Google Scholar] [CrossRef]
- Jeong, D.-W.; Lee, J.-H. Antibiotic resistance, hemolysis and biogenic amine production assessments of Leuconostoc and Weissella isolates for kimchi starter development. LWT Food Sci. Technol. 2015, 64, 1078–1084. [Google Scholar] [CrossRef]
- Jin, Y.H.; Lee, J.H.; Park, Y.K.; Lee, J.-H.; Mah, J.-H. The occurrence of biogenic amines and determination of biogenic amine-producing lactic acid bacteria in kkakdugi and chonggak kimchi. Foods 2019, 8, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.-J.; Kim, K.-S. Tyramine production among lactic acid bacteria and other species isolated from kimchi. LWT Food Sci. Technol. 2014, 56, 406–413. [Google Scholar] [CrossRef]
- Kang, H.-J.; Kim, B.-C.; Park, W. Isolation of tetracycline-resistnt lactic acid bacteria from kimchi. Korean J. Microbiol. 2004, 40, 1–6. (In Korean) [Google Scholar]
- Suvorov, A. What is wrong with enterococcal probiotics? Probiotics Antimicrob. Proteins 2020, 12, 1–4. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Activity of Bacteriocins Produced by | |||||
|---|---|---|---|---|---|
| ST03 | ST06 | ST08 | ST32 | ST109 | |
| Bacillus spp. | 0/12 * | 2/12 | 1/12 | 1/12 | 2/12 |
| Bacillus amyloliquefaciens | 0/2 | 0/2 | 0/2 | 0/2 | 0/2 |
| Bacillus velezensis | 0/1 | 0/1 | 0/1 | 0/1 | 0/1 |
| Bacillus subtilis | 0/4 | 2/4 | 1/4 | 0/4 | 2/4 |
| Bacillus cereus | 0/1 | 0/1 | 0/1 | 0/1 | 0/1 |
| Bacillus pumilus | 0/1 | 0/1 | 0/1 | 0/1 | 0/1 |
| Bacillus sonorensis | 0/1 | 0/1 | 0/1 | 0/1 | 0/1 |
| Bacillus megaterium | 0/1 | 0/1 | 0/1 | 0/1 | 0/1 |
| Bacillus firmus | 0/1 | 0/1 | 0/1 | 1/1 | 0/1 |
| Enterococcus spp. | 0/51 | 1/51 | 8/51 | 3/51 | 5/51 |
| Enterococcus faecium | 0/12 | 1/12 | 2/12 | 1/12 | 1/12 |
| Enterococcus avium | 0/3 | 0/3 | 0/3 | 0/3 | 1/3 |
| Enterococcus faecalis | 0/2 | 0/2 | 0/2 | 0/2 | 0/2 |
| Enterococcus thailandicus | 0/1 | 0/1 | 1/1 | 0/1 | 0/1 |
| Enterococcus durans | 0/1 | 0/1 | 0/1 | 0/1 | 0/1 |
| Enterococcus gallinarum | 0/1 | 0/1 | 1/1 | 1/1 | 0/1 |
| VRE (vancomycin resistant enterococci) | 0/31 | 0/31 | 4/31 | 1/31 | 3/31 |
| Lactobacillus spp. | 0/50 | 0/50 | 0/50 | 6/50 | 0/50 |
| Lactobacillus coryniformis | 0/1 | 0/1 | 0/1 | 0/1 | 0/1 |
| Lactobacillus plantarum | 0/9 | 0/9 | 0/9 | 0/9 | 0/9 |
| Lactobacillus brevis | 0/3 | 0/3 | 0/3 | 2/3 | 0/3 |
| Lactobacillus curvatus | 0/2 | 0/2 | 0/2 | 2/2 | 0/2 |
| Lactobacillus fermentum | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 |
| Lactobacillus paracasei | 0/5 | 0/5 | 0/5 | 1/5 | 0/5 |
| Lactobacillus plantarum | 0/9 | 0/9 | 0/9 | 1/9 | 0/9 |
| Lactobacillus reuteri | 0/2 | 0/2 | 0/2 | 0/2 | 0/2 |
| Lactobacillus rhamnosus | 0/5 | 0/5 | 0/5 | 0/5 | 0/5 |
| Lactobacillus sakei | 0/4 | 0/4 | 0/4 | 0/4 | 0/4 |
| Lactobacillus salivarius | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 |
| Lactobacillus sanikiri | 0/4 | 0/4 | 0/4 | 0/4 | 0/4 |
| Lactococcus spp. | 0/2 | 0/2 | 0/2 | 0/2 | 0/2 |
| Lactococcus lactis | 0/2 | 0/2 | 0/2 | 0/2 | 0/2 |
| Leuconostoc spp. | 0/8 | 0/8 | 0/8 | 2/8 | 0/8 |
| Leuconostoc citreum | 0/1 | 0/1 | 0/1 | 0/1 | 0/1 |
| Leuconostoc mesenteroides | 0/7 | 0/7 | 0/7 | 2/7 | 0/7 |
| Listeria spp. | 5/5 | 5/5 | 5/5 | 5/5 | 5/5 |
| Listeria innocua | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 |
| Listeria monocytogenes | 3/3 | 3/3 | 3/3 | 3/3 | 3/3 |
| Pediococcus spp. | 0/9 | 0/9 | 2/9 | 0/9 | 0/9 |
| Pediococcus acidilactici | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 |
| Pediococcus lolii | 0/1 | 0/1 | 0/1 | 0/1 | 0/1 |
| Pediococcus pentosaceus | 0/5 | 0/5 | 2/5 | 0/5 | 0/5 |
| Weissella spp. | 0/1 | 0/1 | 0/1 | 0/1 | 0/1 |
| Weissella cibaria | 0/1 | 0/1 | 0/1 | 0/1 | 0/1 |
| Total tested strains | 5/138 | 11/138 | 16/138 | 16/138 | 12/138 |
| Carbohydrates | ST03 | ST06 | ST08 | ST32 | ST109 |
|---|---|---|---|---|---|
| control (no carbohydrate) | - | - | - | - | - |
| glycerol, L-arabinose, ribose, D-xylose, D-glucose, D-fructose, D-mannose, inositol, mannitol, sorbitol, α-methyl-D-glucosamine, amygdalin, arbutin, esculin, salicin, cellobiose, maltose, saccharose, trehalose, D-raffinose, amidon, glycogen | + | + | + | + | + |
| L-xylose, α-methyl-D-mannoside | - | - | - | - | + |
| galactose, β-gentiobiose | - | + | - | - | + |
| lactose | + | + | - | + | + |
| melibiose, D-turanose | - | + | + | - | + |
| inulin | - | - | + | - | - |
| erythritol, D-arabinose, adonitol, β-methyl-xyloside, L-sorbose, rhamnose, dulcitol, N-acetylglucosamine, melezitose, xylitol, D-lyxose, D-tagatose, D-fucose, L-fucose, D-arabitol, L-arabitol, gluconate, 2-keto-gluconate, 5-keto-gluconate | - | - | - | - | - |
| Target Enzymes | ST03 | ST06 | ST08 | ST32 | ST109 |
|---|---|---|---|---|---|
| Control | 0 * | 0 | 0 | 0 | 0 |
| alkaline phosphatase | 5 | 3 | 3 | 3 | 3 |
| esterase (C4) | 4 | 4 | 4 | 5 | 3 |
| lipase esterase (C8) | 5 | 4 | 4 | 5 | 3 |
| acid phosphatase | 0 | 0 | 1 | 0 | 3 |
| naphthol phosphohydrolase | 3 | 3 | 3 | 3 | 3 |
| α-galactosidase | 0 | 0 | 1 | 0 | 0 |
| α-glucosidase | 3 | 3 | 5 | 3 | 3 |
| β-glucosidase | 1 | 3 | 4 | 4 | 4 |
| lipase (C14), leucinearylamidase, valinearylamidase, cystinarylamidase, trypsin, α-chymotrypsin, β-galactosidase, β-glucuronidase, N-acetyl-β-glucosaminidase, α-mannosidase, α-fucosidase | 0 | 0 | 0 | 0 | 0 |
| Bacillus velezensis ST03 | Bacillusamyloliquefaciens ST06 | Bacillus subtilis ST08 | Bacillus velezensis ST32 | Bacillusamyloliquefaciens ST109 | |
|---|---|---|---|---|---|
| Antimicrobials related genes | |||||
| Class IIa bacteriocins | - | - | - | - | - |
| Lichenicidin | - | - | + | - | - |
| Thuricin | - | - | - | - | - |
| Surfactin | + | + | + | + | + |
| Iturin | + | + | - | - | + |
| Coagulin | - | - | - | - | - |
| Subtilosin | + | - | + | - | + |
| Pediocin PA-1 | - | - | - | - | - |
| Virulence genes | |||||
| hblA | - | - | - | - | - |
| hblB | - | - | - | - | - |
| hblC | - | - | + | - | - |
| nheA | - | - | - | - | - |
| nheB | - | - | - | - | + |
| nheC | - | - | + | - | - |
| Vancomycin resistance genes | |||||
| vanA | - | - | - | - | - |
| vanB | - | - | - | - | - |
| vanC | - | - | - | - | - |
| vanD | - | - | - | - | - |
| vanE | - | - | - | - | - |
| vanG | - | - | - | - | - |
| Antibiotics | Experimental MIC (μg/mL) Values for Strains | CLSI Cut-Offs (S, I, R) for Bacillus spp. (μg/mL) | EFSA Cut-Offs for Bacillus spp. (μg/mL) | ||||
|---|---|---|---|---|---|---|---|
| ST03 | ST06 | ST08 | ST32 | ST109 | |||
| ampicillin | 128 | 16 | ≤0.25 | 128 | 64 | ≤0.25, -, ≥0.5 | n.r. |
| vancomycin | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | ≤4, *, * | 4 |
| gentamycin | 0.5 | 0.5 | 2 | 0.5 | 0.5 | ≤4, 8, ≥16 | 4 |
| kanamycin | 2 | 2 | 8 | 4 | 16 | ≤1, 2, ≥4 | 8 |
| streptomycin | 8 | 8 | 64 | 8 | 2 | n.r. | 8 |
| erythromycin | 2 | 2 | 2 | 2 | 2 | ≤0.5, 1-2, ≥4 | 4 |
| clindamycin | 0.5 | ≤0.25 | 1 | 0.5 | 0.5 | ≤0.5, 1-2, ≥4 | 4 |
| tetracycline | 4 | 4 | 4 | 8 | ≤0.25 | <4, 8, >16 | 8 |
| chloramphenicol | 4 | 2 | 4 | 1 | 2 | ≤8, 16, ≥32 | 8 |
| ciprofloxacin | ≤0.25 | ≤0.25 | ≤0.25 | ≤0.25 | ≤0.25 | ≤1, 2, ≥4 | n.r. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Irorita Fugaban, J.I.; Vazquez Bucheli, J.E.; Holzapfel, W.H.; Todorov, S.D. Bacteriocinogenic Bacillus spp. Isolated from Korean Fermented Cabbage (Kimchi)—Beneficial or Hazardous? Fermentation 2021, 7, 56. https://doi.org/10.3390/fermentation7020056
Irorita Fugaban JI, Vazquez Bucheli JE, Holzapfel WH, Todorov SD. Bacteriocinogenic Bacillus spp. Isolated from Korean Fermented Cabbage (Kimchi)—Beneficial or Hazardous? Fermentation. 2021; 7(2):56. https://doi.org/10.3390/fermentation7020056
Chicago/Turabian StyleIrorita Fugaban, Joanna Ivy, Jorge Enrique Vazquez Bucheli, Wilhelm Heinrich Holzapfel, and Svetoslav Dimitrov Todorov. 2021. "Bacteriocinogenic Bacillus spp. Isolated from Korean Fermented Cabbage (Kimchi)—Beneficial or Hazardous?" Fermentation 7, no. 2: 56. https://doi.org/10.3390/fermentation7020056
APA StyleIrorita Fugaban, J. I., Vazquez Bucheli, J. E., Holzapfel, W. H., & Todorov, S. D. (2021). Bacteriocinogenic Bacillus spp. Isolated from Korean Fermented Cabbage (Kimchi)—Beneficial or Hazardous? Fermentation, 7(2), 56. https://doi.org/10.3390/fermentation7020056