
A Statistical Workflow to Evaluate the Modulation of Wine Metabolome and Its Contribution to the Sensory Attributes


 , , and
, , and 
Abstract
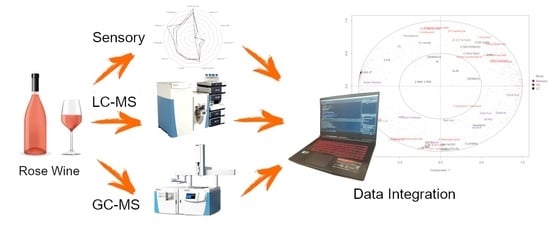
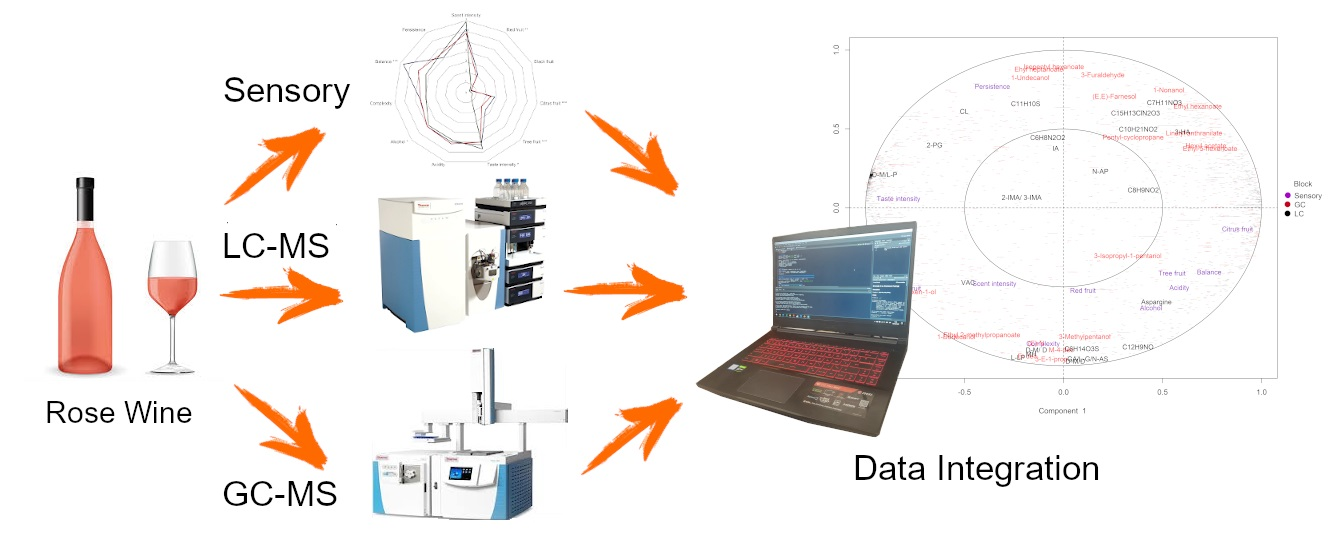
:
1. Introduction
2. Materials and Methods
2.1. Wine Samples
2.2. Analysis, Data Acquisition and Processing of Headspace Solid-Phase Microextraction Coupled with Gas Chromatography-Mass Spectrometry (HS-SPME-GC-MS) Data
2.3. Analysis, Data Acquisition and Processing of Using Ultra-High-Performance Liquid Chromatography High-Resolution Mass Spectrometry (UHPLC-HRMS) Data
2.4. Sensory Analysis
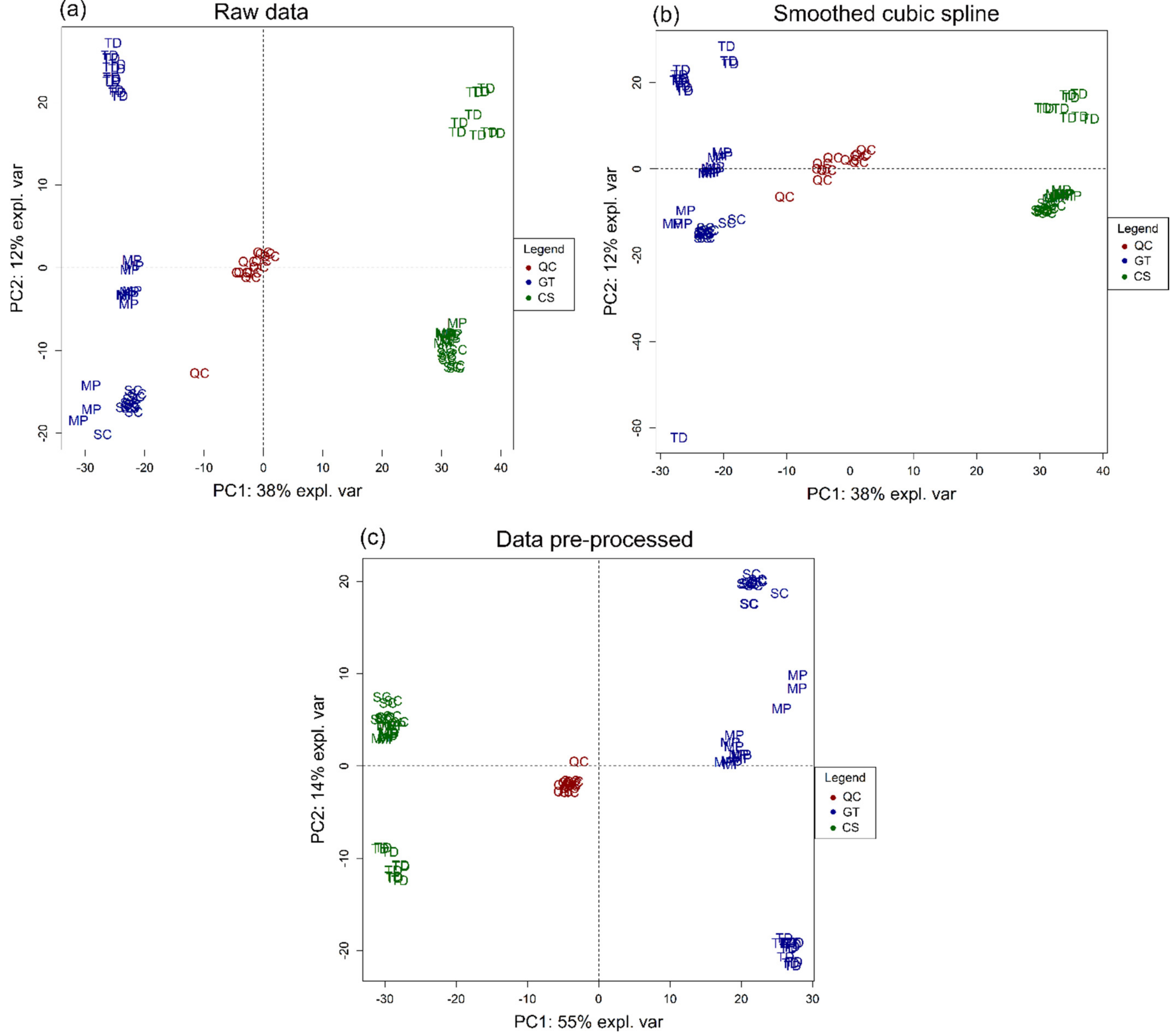
2.5. Statistical Analysis
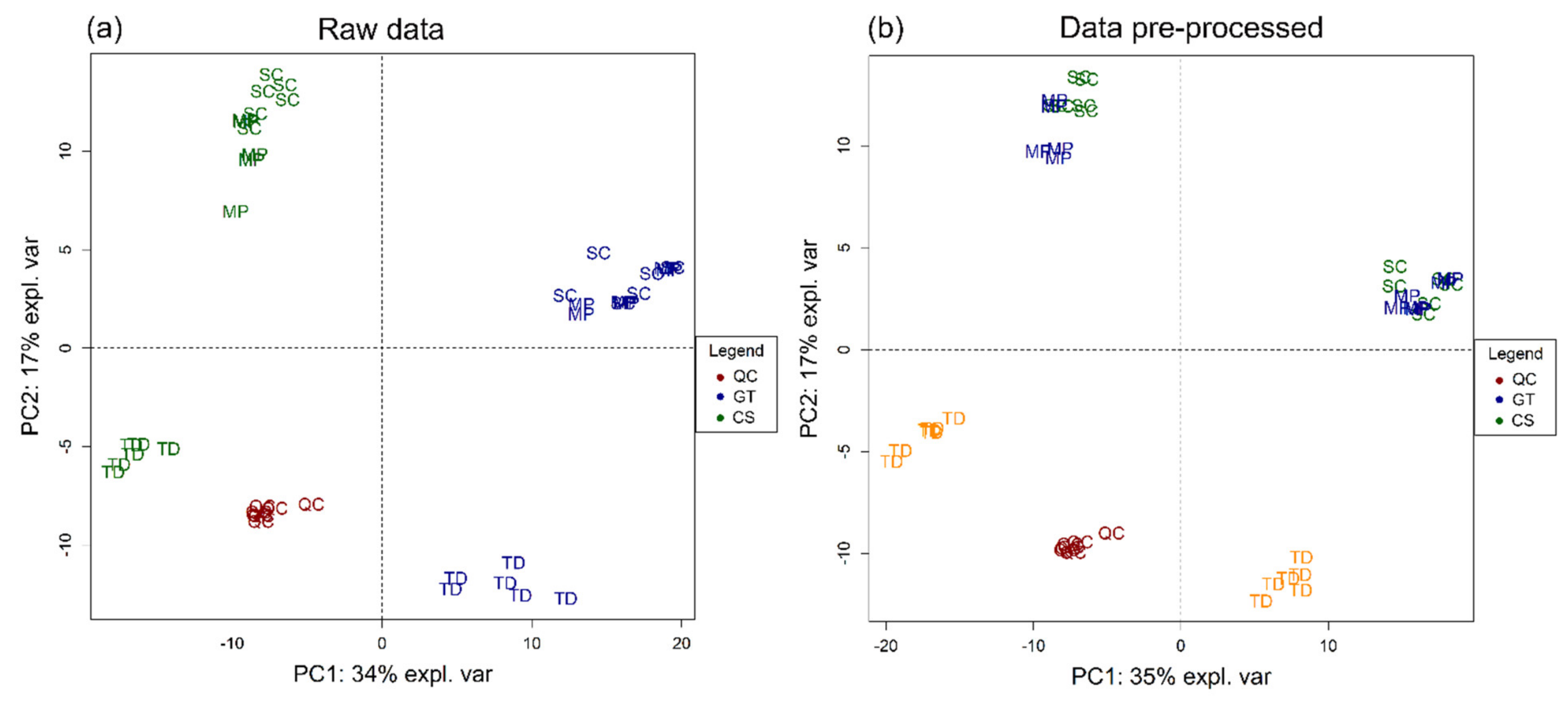
3. Results
3.1. Volatile Profile
3.2. Non-Volatile Profile
3.3. Sensory Attributes
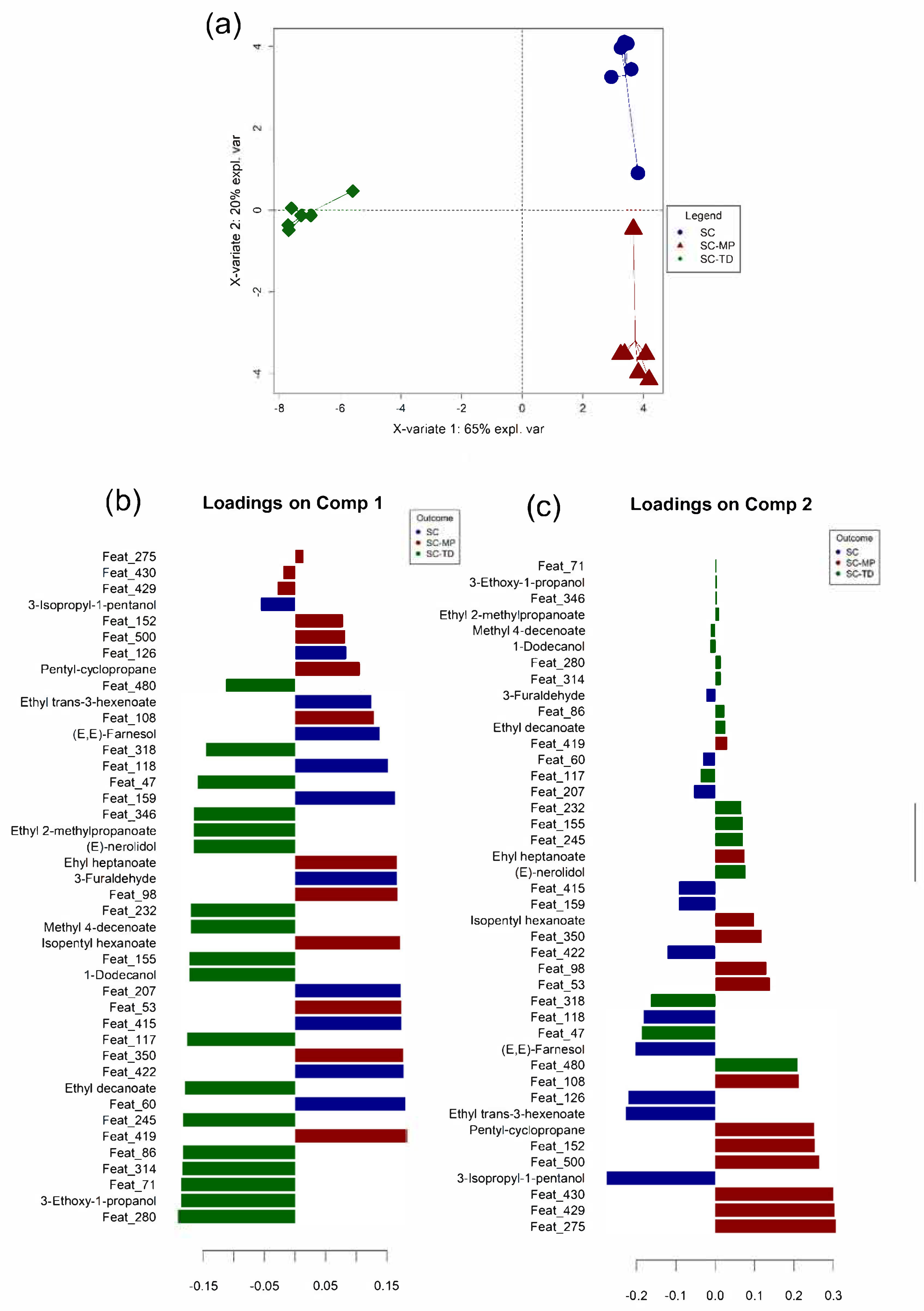
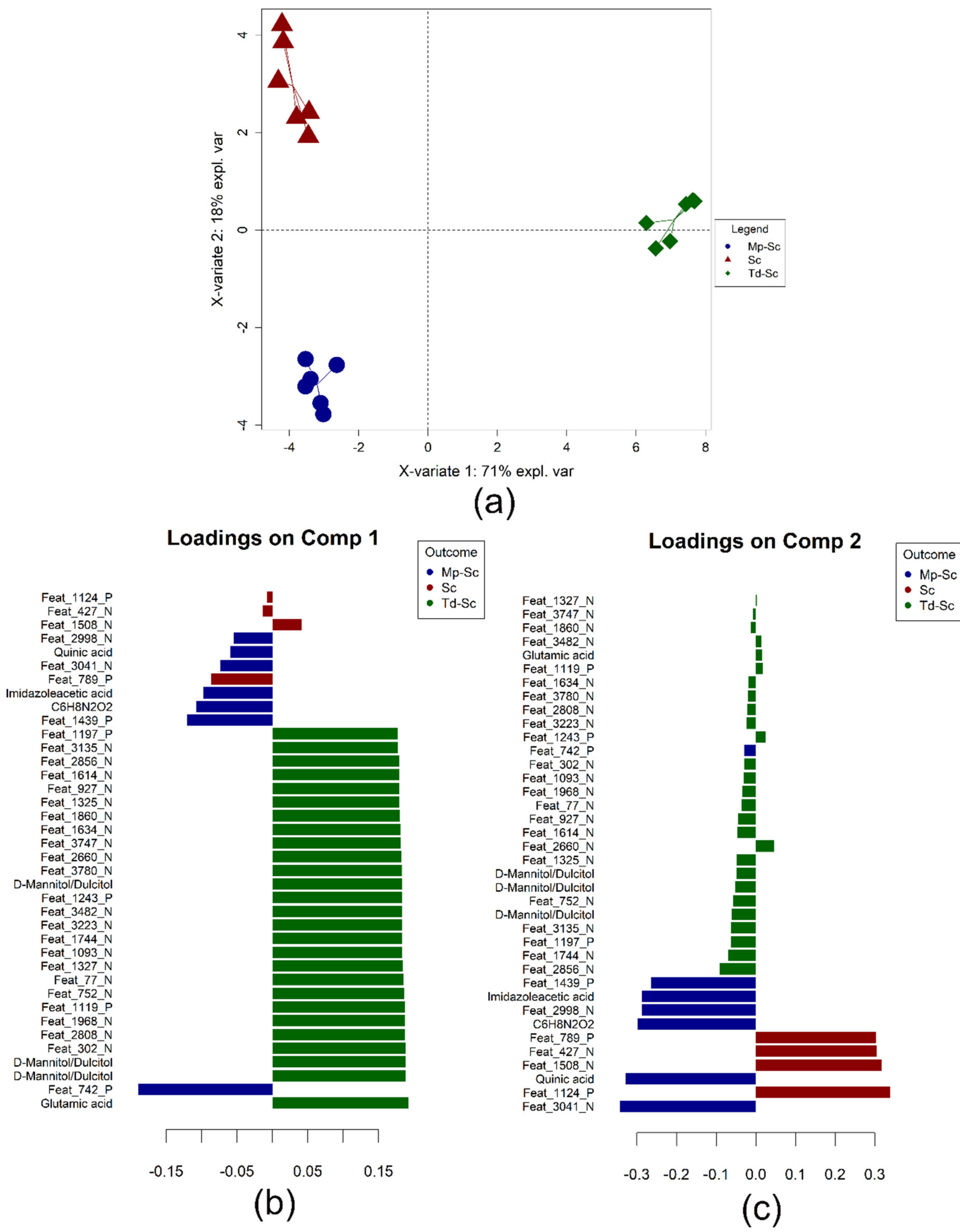
3.4. Data Integration: Sparse Generalised Canonical Correlation Analysis Discriminant Analysis (sGCC-DA) Approach
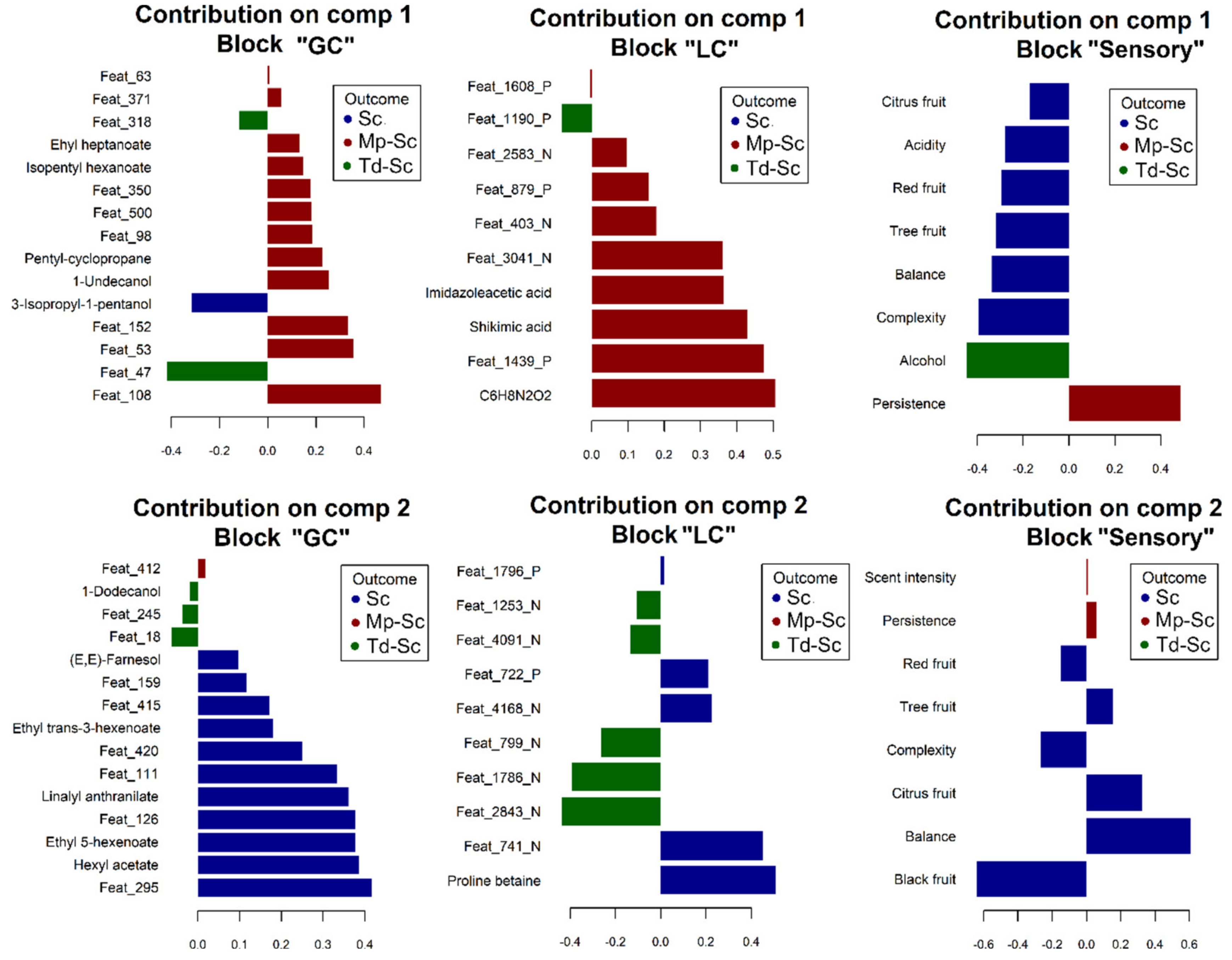
3.5. Partial Least Squares (PLS) Regression
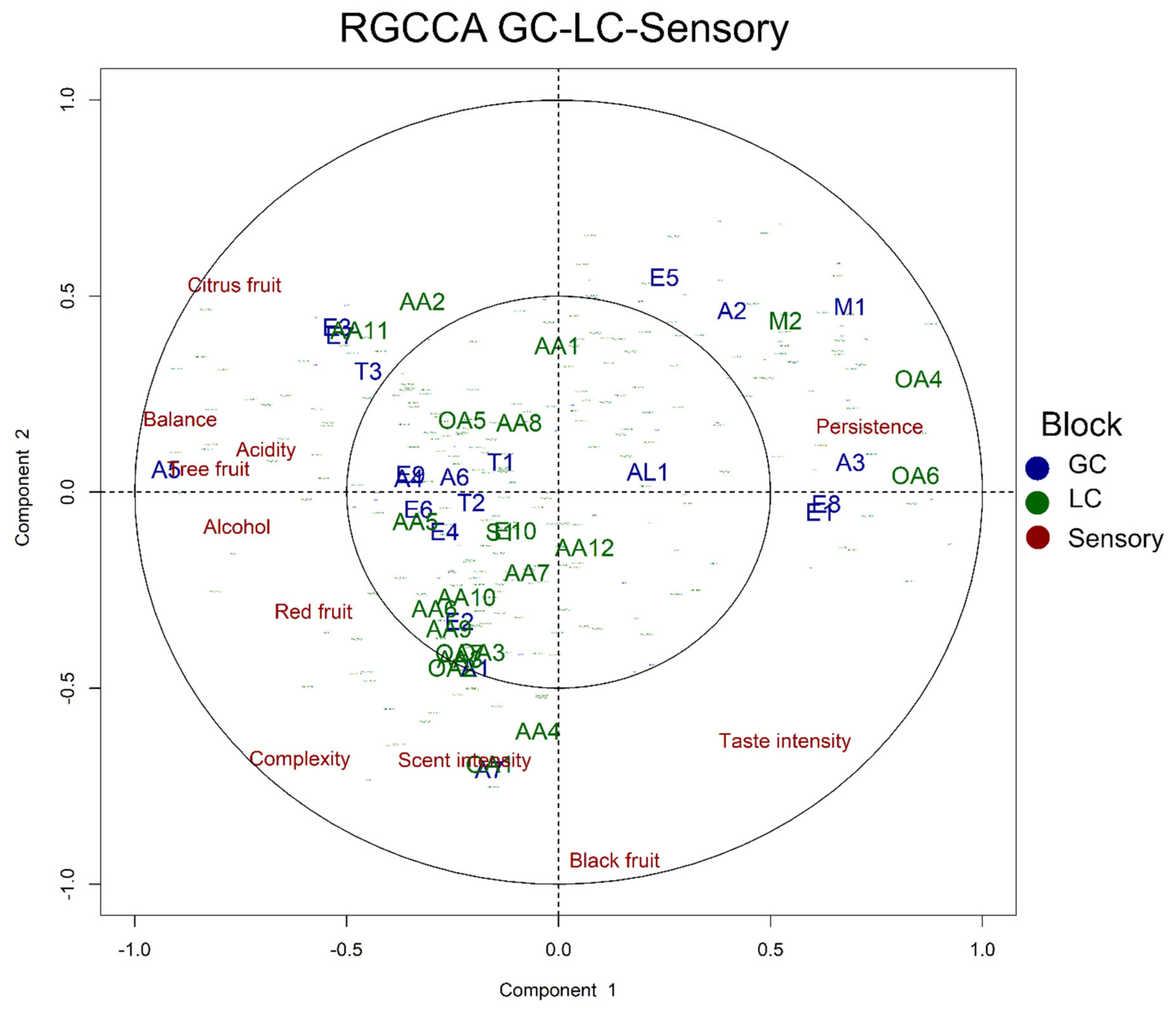
3.6. Data Integration: Regularised Generalised Canonical Correlation Analysis (RGCCA) Approach
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegrist, M.; Cousin, M.-E. Expectations Influence Sensory Experience in a Wine Tasting. Appetite 2009, 52, 762–765. [Google Scholar] [CrossRef] [PubMed]
- Sherman, E.; Coe, M.; Grose, C.; Martin, D.; Greenwood, D.R. Metabolomics Approach to Assess the Relative Contributions of the Volatile and Non-Volatile Composition to Expert Quality Ratings of Pinot Noir Wine Quality. J. Agric. Food Chem. 2020, 68, 13380–13396. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Lu, Y.; Guo, Y.; Cao, H.; Wang, Q.; Shui, W. Comprehensive Evaluation of Untargeted Metabolomics Data Processing Software in Feature Detection, Quantification and Discriminating Marker Selection. Anal. Chim. Acta 2018, 1029, 50–57. [Google Scholar] [CrossRef]
- Arapitsas, P.; Ugliano, M.; Marangon, M.; Piombino, P.; Rolle, L.; Gerbi, V.; Versari, A.; Mattivi, F. Use of Untargeted Liquid Chromatography–Mass Spectrometry Metabolome to Discriminate Italian Monovarietal Red Wines, Produced in Their Different Terroirs. J. Agric. Food Chem. 2020, 68, 13353–13366. [Google Scholar] [CrossRef]
- Šuklje, K.; Carlin, S.; Stanstrup, J.; Antalick, G.; Blackman, J.W.; Meeks, C.; Deloire, A.; Schmidtke, L.M.; Vrhovsek, U. Unravelling Wine Volatile Evolution during Shiraz Grape Ripening by Untargeted HS-SPME-GC × GC-TOFMS. Food Chem. 2019, 277, 753–765. [Google Scholar] [CrossRef] [PubMed]
- Pinu, F.R. Grape and Wine Metabolomics to Develop New Insights Using Untargeted and Targeted Approaches. Fermentation 2018, 4, 92. [Google Scholar] [CrossRef] [Green Version]
- Rocchetti, G.; Gatti, M.; Bavaresco, L.; Lucini, L. Untargeted Metabolomics to Investigate the Phenolic Composition of Chardonnay Wines from Different Origins. J. Food Compos. Anal. 2018, 71, 87–93. [Google Scholar] [CrossRef]
- Whitener, M.E.B.; Stanstrup, J.; Panzeri, V.; Carlin, S.; Divol, B.; Du Toit, M.; Vrhovsek, U. Untangling the Wine Metabolome by Combining Untargeted SPME–GCxGC-TOF-MS and Sensory Analysis to Profile Sauvignon Blanc Co-Fermented with Seven Different Yeasts. Metabolomics 2016, 12, 53. [Google Scholar] [CrossRef]
- Arbulu, M.; Sampedro, M.C.; Gómez-Caballero, A.; Goicolea, M.A.; Barrio, R.J. Untargeted Metabolomic Analysis Using Liquid Chromatography Quadrupole Time-of-Flight Mass Spectrometry for Non-Volatile Profiling of Wines. Anal. Chim. Acta 2015, 858, 32–41. [Google Scholar] [CrossRef]
- Castro, C.C.; Martins, R.C.; Teixeira, J.A.; Ferreira, A.C.S. Application of a High-Throughput Process Analytical Technology Metabolomics Pipeline to Port Wine Forced Ageing Process. Food Chem. 2014, 143, 384–391. [Google Scholar] [CrossRef] [Green Version]
- Goodacre, R.; Broadhurst, D.; Smilde, A.K.; Kristal, B.S.; Baker, J.D.; Beger, R.; Bessant, C.; Connor, S.; Capuani, G.; Craig, A. Proposed Minimum Reporting Standards for Data Analysis in Metabolomics. Metabolomics 2007, 3, 231–241. [Google Scholar] [CrossRef]
- Johnsen, L.G.; Skou, P.B.; Khakimov, B.; Bro, R. Gas Chromatography–Mass Spectrometry Data Processing Made Easy. J. Chromatogr. A 2017, 1503, 57–64. [Google Scholar] [CrossRef]
- Amigo, J.M.; Skov, T.; Bro, R.; Coello, J.; Maspoch, S. Solving GC-MS Problems with Parafac2. TrAC Trends Anal. Chem. 2008, 27, 714–725. [Google Scholar] [CrossRef]
- Mahieu, N.G.; Genenbacher, J.L.; Patti, G.J. A Roadmap for the XCMS Family of Software Solutions in Metabolomics. Curr. Opin. Chem. Biol. 2016, 30, 87–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Considine, E.C.; Salek, R.M. A Tool to Encourage Minimum Reporting Guideline Uptake for Data Analysis in Metabolomics. Metabolites 2019, 9, 43. [Google Scholar] [CrossRef] [Green Version]
- Ren, S.; Hinzman, A.A.; Kang, E.L.; Szczesniak, R.D.; Lu, L.J. Computational and Statistical Analysis of Metabolomics Data. Metabolomics 2015, 11, 1492–1513. [Google Scholar] [CrossRef]
- Worley, B.; Powers, R. Multivariate Analysis in Metabolomics. Curr. Metab. 2013, 1, 92–107. [Google Scholar]
- Rohart, F.; Gautier, B.; Singh, A.; Le Cao, K.-A. MixOmics: An R Package for ‘omics Feature Selection and Multiple Data Integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanichthanarak, K.; Fahrmann, J.F.; Grapov, D. Genomic, Proteomic, and Metabolomic Data Integration Strategies. Biomark. Insights 2015, 10, BMI–S29511. [Google Scholar] [CrossRef] [Green Version]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.-M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed Minimum Reporting Standards for Chemical Analysis. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, C.A.; Want, E.J.; O’Maille, G.; Abagyan, R.; Siuzdak, G. XCMS: Processing Mass Spectrometry Data for Metabolite Profiling Using Nonlinear Peak Alignment, Matching, and Identification. Anal. Chem. 2006, 78, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Libiseller, G.; Dvorzak, M.; Kleb, U.; Gander, E.; Eisenberg, T.; Madeo, F.; Neumann, S.; Trausinger, G.; Sinner, F.; Pieber, T.; et al. IPO: A Tool for Automated Optimization of XCMS Parameters. BMC Bioinform. 2015, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhl, C.; Tautenhahn, R.; Böttcher, C.; Larson, T.R.; Neumann, S. CAMERA: An Integrated Strategy for Compound Spectra Extraction and Annotation of LC/MS Data Sets. Anal. Chem. 2012, 84, 283–289. [Google Scholar] [CrossRef] [Green Version]
- Arapitsas, P.; Della Corte, A.; Gika, H.; Narduzzi, L.; Mattivi, F.; Theodoridis, G. Studying the Effect of Storage Conditions on the Metabolite Content of Red Wine Using HILIC LC–MS Based Metabolomics. Food Chem. 2016, 197, 1331–1340. [Google Scholar] [CrossRef] [PubMed]
- Puertas, B.; Jimenez-Hierro, M.J.; Cantos-Villar, E.; Marrufo-Curtido, A.; Carbú, M.; Cuevas, F.J.; Moreno-Rojas, J.M.; González-Rodríguez, V.E.; Cantoral, J.M.; Ruiz-Moreno, M.J. The Influence of Yeast on Chemical Composition and Sensory Properties of Dry White Wines. Food Chem. 2018, 253, 227–235. [Google Scholar] [CrossRef]
- Kokla, M.; Klåvus, A.; Noerman, S.; Koistinen, V.M.; Tuomainen, M.; Zarei, I.; Meuronen, T.; Häkkinen, M.R.; Rummukainen, S.; Babu, A.F. “NoTaMe”: Workflow for Non-Targeted LC-MS Metabolic Profiling. Metabolites 2020, 10, 135. [Google Scholar]
- Kokla, M.; Virtanen, J.; Kolehmainen, M.; Paananen, J.; Hanhineva, K. Random Forest-Based Imputation Outperforms Other Methods for Imputing LC-MS Metabolomics Data: A Comparative Study. BMC Bioinform. 2019, 20, 492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noonan, M.J.; Tinnesand, H.V.; Buesching, C.D. Normalizing Gas-Chromatography–Mass Spectrometry Data: Method Choice Can Alter Biological Inference. BioEssays 2018, 40, 1700210. [Google Scholar] [CrossRef]
- Szymańska, E.; Saccenti, E.; Smilde, A.K.; Westerhuis, J.A. Double-Check: Validation of Diagnostic Statistics for PLS-DA Models in Metabolomics Studies. Metabolomics 2012, 8, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Redondo, J.M.; Ruiz-Moreno, M.J.; Puertas, B.; Cantos-Villar, E.; Moreno-Rojas, J.M. Multivariate Optimization of Headspace Solid-Phase Microextraction Coupled to Gas Chromatography-Mass Spectrometry for the Analysis of Terpenoids in Sparkling Wines. Talanta 2020, 208, 120483. [Google Scholar] [CrossRef]
- Shi, L.; Westerhuis, J.A.; Rosén, J.; Landberg, R.; Brunius, C. Variable Selection and Validation in Multivariate Modelling. Bioinformatics 2019, 35, 972–980. [Google Scholar] [CrossRef] [PubMed]
- Sadoudi, M.; Tourdot-Maréchal, R.; Rousseaux, S.; Steyer, D.; Gallardo-Chacón, J.-J.; Ballester, J.; Vichi, S.; Guérin-Schneider, R.; Caixach, J.; Alexandre, H. Yeast–Yeast Interactions Revealed by Aromatic Profile Analysis of Sauvignon Blanc Wine Fermented by Single or Co-Culture of Non-Saccharomyces and Saccharomyces Yeasts. Food Microbiol. 2012, 32, 243–253. [Google Scholar] [CrossRef]
- Renault, P.; Miot-Sertier, C.; Marullo, P.; Hernández-Orte, P.; Lagarrigue, L.; Lonvaud-Funel, A.; Bely, M. Genetic Characterization and Phenotypic Variability in Torulaspora Delbrueckii Species: Potential Applications in the Wine Industry. Int. J. Food Microbiol. 2009, 134, 201–210. [Google Scholar] [CrossRef]
- Benito, S. The Impact of Torulaspora Delbrueckii Yeast in Winemaking. Appl. Microbiol. Biotechnol. 2018, 102, 3081–3094. [Google Scholar] [CrossRef] [PubMed]
- Velázquez, R.; Zamora, E.; Álvarez, M.L.; Ramírez, M. Using Torulaspora Delbrueckii Killer Yeasts in the Elaboration of Base Wine and Traditional Sparkling Wine. Int. J. Food Microbiol. 2019, 289, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Sumby, K.M.; Grbin, P.R.; Jiranek, V. Microbial Modulation of Aromatic Esters in Wine: Current Knowledge and Future Prospects. Food Chem. 2010, 121, 1–16. [Google Scholar] [CrossRef]
- Oliveira, I.; Ferreira, V. Modulating Fermentative, Varietal and Aging Aromas of Wine Using Non-Saccharomyces Yeasts in a Sequential Inoculation Approach. Microorganisms 2019, 7, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renault, P.; Coulon, J.; de Revel, G.; Barbe, J.-C.; Bely, M. Increase of Fruity Aroma during Mixed T. Delbrueckii/S. Cerevisiae Wine Fermentation Is Linked to Specific Esters Enhancement. Int. J. Food Microbiol. 2015, 207, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Gobert, A.; Tourdot-Maréchal, R.; Morge, C.; Sparrow, C.; Liu, Y.; Quintanilla-Casas, B.; Vichi, S.; Alexandre, H. Non-Saccharomyces Yeasts Nitrogen Source Preferences: Impact on Sequential Fermentation and Wine Volatile Compounds Profile. Front. Microbiol. 2017, 8, 2175. [Google Scholar] [CrossRef] [Green Version]
- King, A.; Richard Dickinson, J. Biotransformation of Monoterpene Alcohols by Saccharomyces Cerevisiae, Torulaspora Delbrueckii and Kluyveromyces Lactis. Yeast 2000, 16, 499–506. [Google Scholar] [CrossRef]
- Jiang, B.; Zhang, Z. Volatile Compounds of Young Wines from Cabernet Sauvignon, Cabernet Gernischet and Chardonnay Varieties Grown in the Loess Plateau Region of China. Molecules 2010, 15, 9184–9196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coelho, E.; Coimbra, M.A.; Nogueira, J.M.F.; Rocha, S.M. Quantification Approach for Assessment of Sparkling Wine Volatiles from Different Soils, Ripening Stages, and Varieties by Stir Bar Sorptive Extraction with Liquid Desorption. Anal. Chim. Acta 2009, 635, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Soetaert, W.; Vanhooren, P.T.; Vandamme, E.J. The production of mannitol by fermentation. In Carbohydrate Biotechnology Protocols; Springer: Berlin/Heidelberg, Germany, 1999; pp. 261–275. [Google Scholar]
- Azzolini, M.; Tosi, E.; Lorenzini, M.; Finato, F.; Zapparoli, G. Contribution to the Aroma of White Wines by Controlled Torulaspora Delbrueckii Cultures in Association with Saccharomyces Cerevisiae. World J. Microbiol. Biotechnol. 2015, 31, 277–293. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analytical Platform | Model | Mean Overall BER (1) | Ncomp | Class | Mean Class Error (2) | p-Value (3) |
|---|---|---|---|---|---|---|
| GC-MS | Allvariables | 0.11 ± 0.08 | 3 | Sc-Td | 0.01 ± 0.03 | BER: <0.001 |
| Sc-Mp | 0.15 ± 0.16 | NMC: 0.024 | ||||
| Sc | 0.18 ± 0.17 | AUROC: 0.020 | ||||
| GC-MS | Variable reduction | 0.08 ± 0.06 | 2 | Sc-Td | 0.00 ± 0.00 | BER: <0.001 |
| Sc-Mp | 0.12 ± 0.11 | NMC: 0.007 | ||||
| Sc | 0.14 ± 0.11 | AUROC: 0.003 | ||||
| LC-MS | Allvariables | 0.17 ± 0.10 | 4 | Sc-Td | 0.14 ± 0.09 | BER: <0.001 |
| Sc-Mp | 0.07 ± 0.13 | NMC: 0.120 | ||||
| Sc | 0.29 ± 0.21 | AUROC: 0.035 | ||||
| LC-MS | Variable reduction | 0.02 ± 0.04 | 2 | Sc-Td | 0.00 ± 0.00 | BER: <0.001 |
| Sc-Mp | 0.05 ± 0.11 | NMC: 0.027 | ||||
| Sc | 0.00 ± 0.02 | AUROC: 0.002 |
| Sensory Descriptor | Sc-Td1 | Sc-Mp 2 | Sc 3 | p-Value 4 | CS 5 | GT 6 | p-Value | Interactions (p-Value) |
|---|---|---|---|---|---|---|---|---|
| Scent intensity | 6.0b | 6.2ab | 6.6a | * | 6.4 | 6.1 | ns | *** |
| Red fruit | 1.35b | 1.97a | 2.01a | *** | 1.90 | 1.66 | ns | ns |
| Black fruit | 1.47b | 1.99a | 2.03a | * | 2.95a | 0.71b | *** | *** |
| Citrus fruit | 0.86b | 1.65a | 1.84a | *** | 1.00b | 1.90a | *** | *** |
| Tree fruit | 1.10b | 1.78a | 2.12a | *** | 1.58 | 1.75 | ns | ** |
| Taste intensity | 5.8 | 5.5 | 5.6 | ns | 5.9a | 5.4b | ** | ns |
| Acidity | 4.5b | 4.9a | 5.0a | ** | 4.6b | 5.0a | * | ns |
| Alcohol | 4.7b | 5.0a | 4.9a | ** | 4.8 | 4.9 | ns | ns |
| Complexity | 4.1b | 4.6a | 4.7a | *** | 4.6a | 4.3b | * | ns |
| Balance | 4.1c | 5.0b | 5.7a | *** | 4.5b | 5.3a | *** | ** |
| Persistence | 5.1a | 4.2b | 4.3b | *** | 4.7 | 4.4 | ns | * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muñoz-Redondo, J.M.; Puertas, B.; Pereira-Caro, G.; Ordóñez-Díaz, J.L.; Ruiz-Moreno, M.J.; Cantos-Villar, E.; Moreno-Rojas, J.M. A Statistical Workflow to Evaluate the Modulation of Wine Metabolome and Its Contribution to the Sensory Attributes. Fermentation 2021, 7, 72. https://doi.org/10.3390/fermentation7020072
Muñoz-Redondo JM, Puertas B, Pereira-Caro G, Ordóñez-Díaz JL, Ruiz-Moreno MJ, Cantos-Villar E, Moreno-Rojas JM. A Statistical Workflow to Evaluate the Modulation of Wine Metabolome and Its Contribution to the Sensory Attributes. Fermentation. 2021; 7(2):72. https://doi.org/10.3390/fermentation7020072
Chicago/Turabian StyleMuñoz-Redondo, José Manuel, Belén Puertas, Gema Pereira-Caro, José Luis Ordóñez-Díaz, María José Ruiz-Moreno, Emma Cantos-Villar, and José Manuel Moreno-Rojas. 2021. "A Statistical Workflow to Evaluate the Modulation of Wine Metabolome and Its Contribution to the Sensory Attributes" Fermentation 7, no. 2: 72. https://doi.org/10.3390/fermentation7020072
APA StyleMuñoz-Redondo, J. M., Puertas, B., Pereira-Caro, G., Ordóñez-Díaz, J. L., Ruiz-Moreno, M. J., Cantos-Villar, E., & Moreno-Rojas, J. M. (2021). A Statistical Workflow to Evaluate the Modulation of Wine Metabolome and Its Contribution to the Sensory Attributes. Fermentation, 7(2), 72. https://doi.org/10.3390/fermentation7020072