
Aeration and Stirring in Yarrowia lipolytica Lipase Biosynthesis during Batch Cultures with Waste Fish Oil as a Carbon Source




Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Microorganism
2.3. Media and Culture Conditions
2.4. Determination of Biomass
2.5. Determination of Extracellular Lipase Activity
2.6. Determination of Selected Culture Parameters
2.7. Statistical Analysis
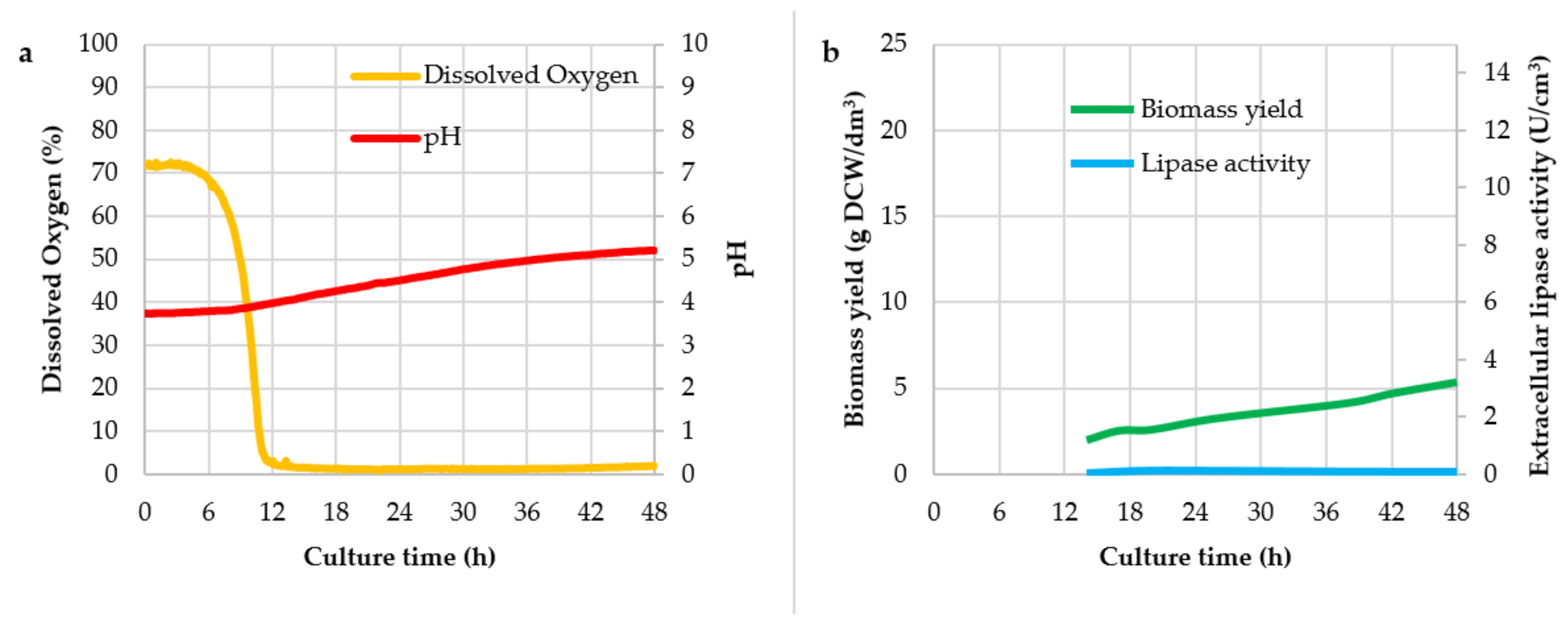
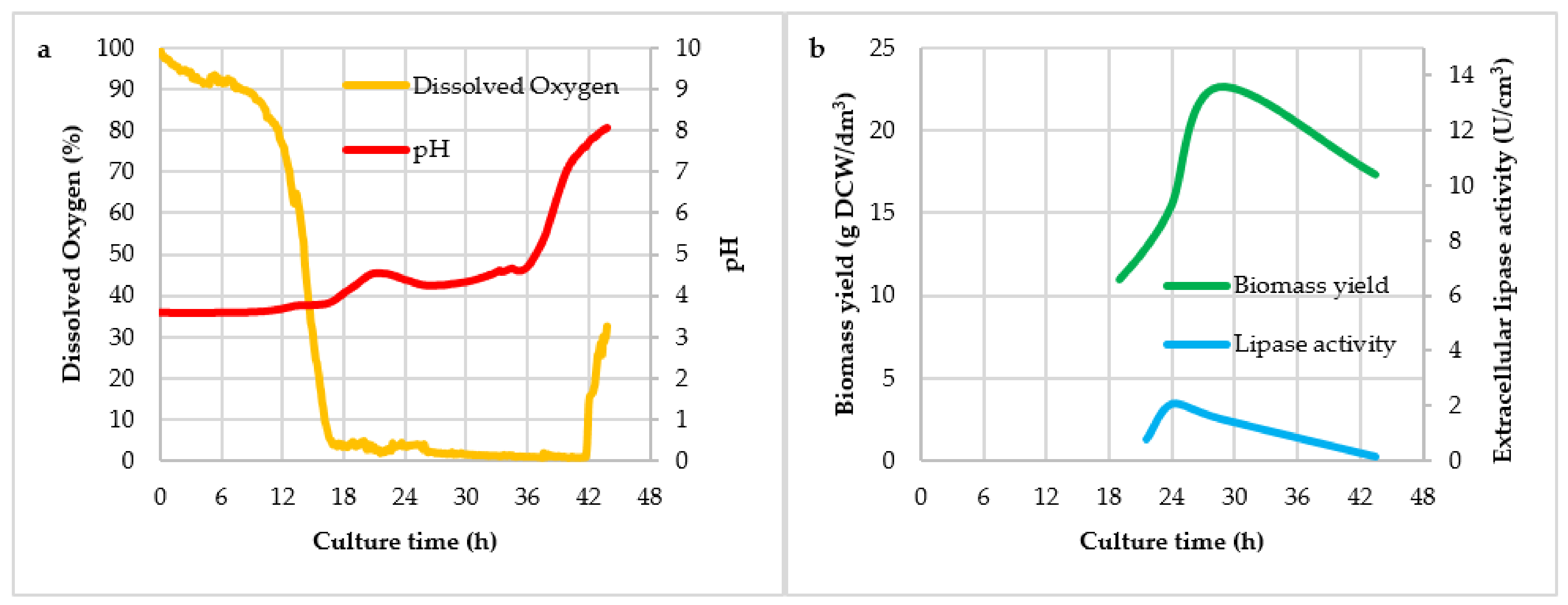
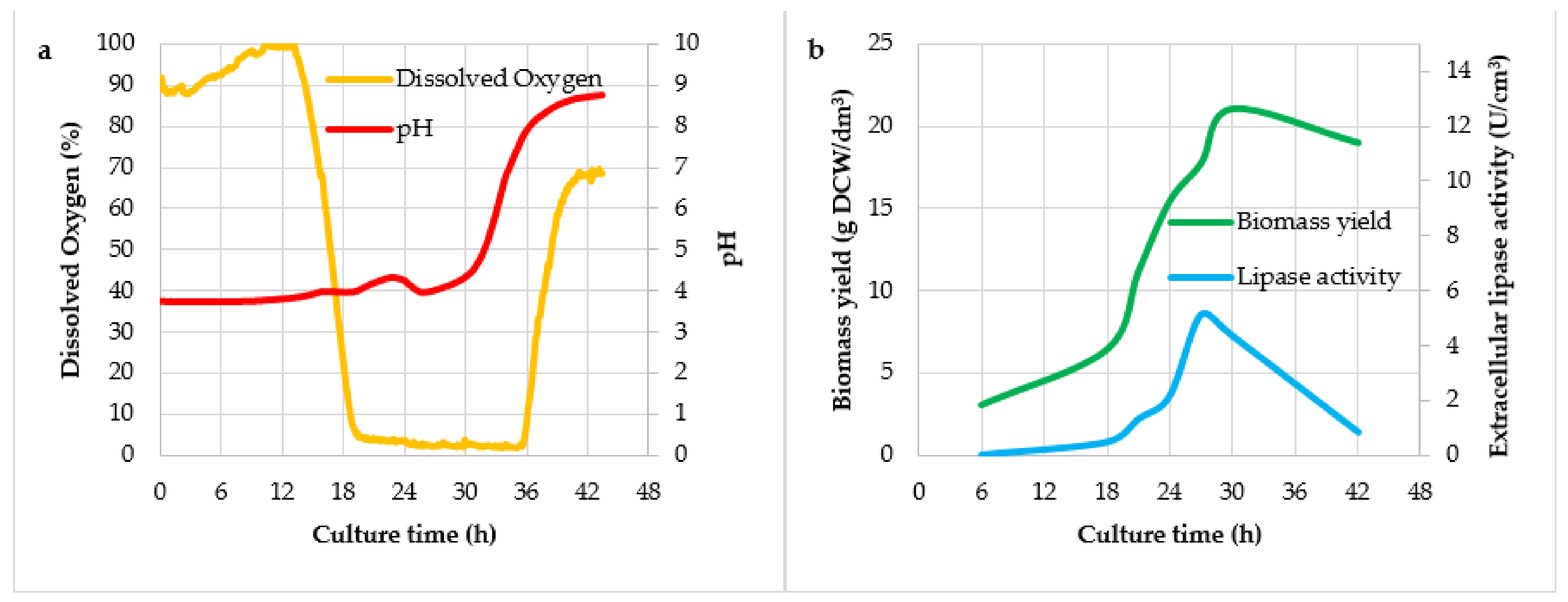
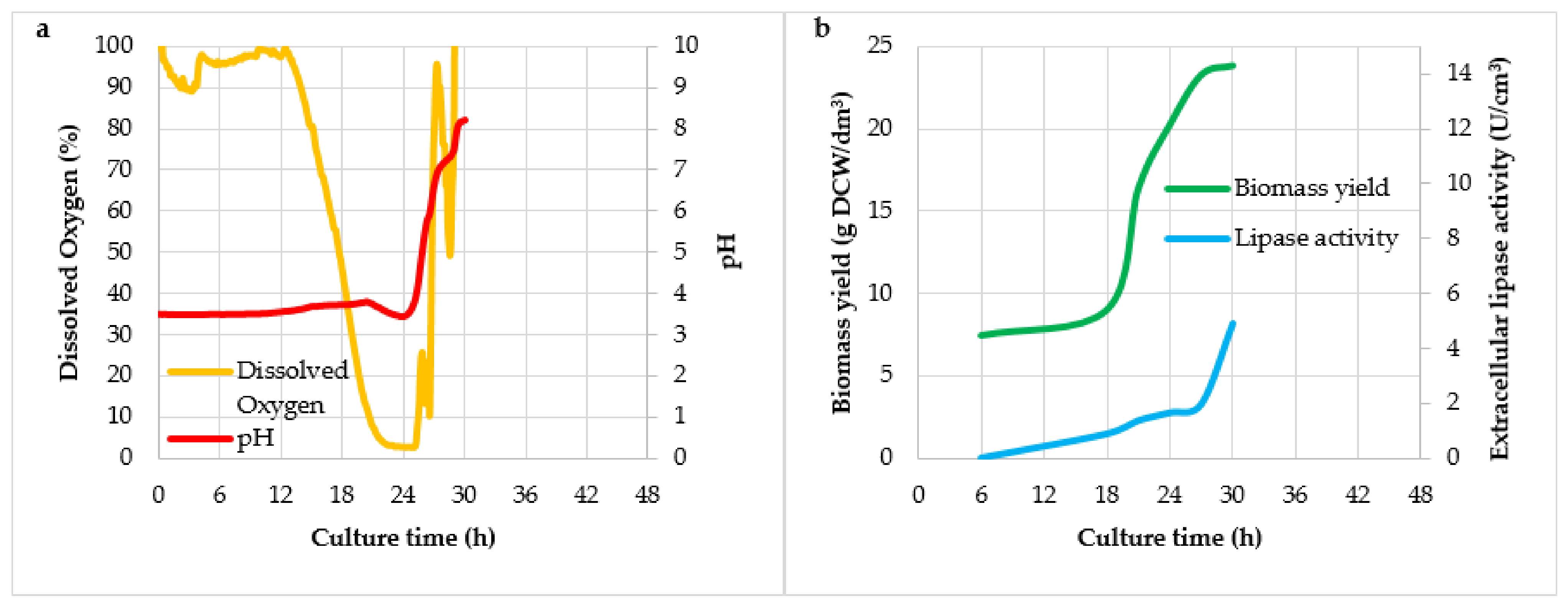
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gupta, P.; Chaubey, A.; Mahajan, N.; Anand, N. A review on Arthrobacter sp. lipase: A versatile biocatalyst for the kinetic resolution to access enantiomerically pure/enriched compounds. Chirality 2021, 33, 209–225. [Google Scholar] [CrossRef]
- Kamil, J.P.; Tischer, B.K.; Trapp, S.; Nair, V.K.; Osterrieder, N.; Kung, H.J. vLIP, a Viral Lipase Homologue, Is a Virulence Factor of Marek’s Disease Virus. J. Virol. 2005, 6, 6984–6996. [Google Scholar] [CrossRef] [Green Version]
- Zeng, S.; Liu, J.; Anankanbil, S.; Chen, M.; Guo, Z.; Adams, J.P.; Snajdrova, R.; Li, Z. Amide Synthesis via Aminolysis of Ester or Acid with an Intracellular Lipase. ACS Catal. 2018, 8, 8856–8865. [Google Scholar] [CrossRef]
- Zhou, N.; Shen, L.; Dong, Z.; Shen, J.; Du, L.; Luo, X. Enzymatic Synthesis of Thioesters from Thiols and Vinyl Esters in a Continuous-Flow Microreactor. Catalysts 2018, 8, 249. [Google Scholar] [CrossRef] [Green Version]
- Mhetras, N.; Mapare, V.; Gokhale, D. Cold Active Lipases: Biocatalytic Tools for Greener Technology. Appl. Biochem. Biotechnol. 2021. [Google Scholar] [CrossRef]
- Coelho, A.L.S.; Orlandelli, R.C. Immobilized microbial lipases in the food industry: A systematic literature review. Crit. Rev. Food Sci. Nutr. 2021, 61, 1689–1703. [Google Scholar] [CrossRef] [PubMed]
- Vishnoi, N.; Dixit, S.; Mishra, J. Microbial Lipases and Their Versatile Applications. In Microbial Enzymes: Roles and Applications in Industries. Microorganisms for Sustainability; Arora, N., Mishra, J., Mishra, V., Eds.; Springer: Singapore, 2020; Volume 11, pp. 207–230. [Google Scholar] [CrossRef]
- Mehta, A.; Guleria, S.; Sharma, R.; Gupta, R. 6-The lipases and their applications with emphasis on food industry. In Microbial Biotechnology in Food and Health; Ray, R.C., Ed.; Academic Press: Cambridge, MA, USA, 2021; pp. 143–164. [Google Scholar] [CrossRef]
- Dwivedee, B.; Soni, S.; Sharma, M.; Bhaumik, J.; Laha, J.K.; Banerjee, U.C. Promiscuity of Lipase-Catalyzed Reactions for Organic Synthesis: A Recent Update. ChemistrySelect 2018, 3, 2441–2446. [Google Scholar] [CrossRef]
- Chandra, P.; Enespa; Singh, R.; Arora, P.K. Microbial lipases and their industrial applications: A comprehensive review. Microb. Cell Factories 2020, 19, 169. [Google Scholar] [CrossRef]
- Gurung, N.; Ray, S.; Bose, S.; Rai, V. A Broader View: Microbial Enzymes and Their Relevance in Industries, Medicine, and Beyond. BioMed. Res. Int. 2013, 329121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kebabci, O.; Cihangir, N. Comparison of three Yarrowia lipolytica strains for lipase production: NBRC 1658, IFO 1195, and a local strain. Turk. J. Biol. 2012, 36, 15–24. [Google Scholar]
- Lopes, M.; Araujo, C.; Aguedo, M.; Gomes, N.; Goncalves, C.; Teixeira, J.A.; Belo, I. The use of olive mill wastewater by wild type Yarrowia lipolytica strains: Medium supplementation and surfactant presence effect. J. Chem. Technol. Biotechnol. 2009, 84, 533–537. [Google Scholar] [CrossRef] [Green Version]
- Fickers, P.; Nicaud, J.M.; Gaillardin, C.; Destain, J.; Thonart, P. Carbon and nitrogen sources modulate lipase production in the yeast Yarrowia lipolytica. J. Appl. Microbiol. 2004, 96, 742–749. [Google Scholar] [CrossRef] [PubMed]
- Fabiszewska, A.U.; Białecka-Florjańczyk, E. Factors influencing synthesis of extracellular lipases by Yarrowia lipolytica in medium containing vegetable oils. J. Microbiol. Biotechnol. Food Sci. 2014, 4, 231–237. [Google Scholar] [CrossRef] [Green Version]
- Bharathi, D.; Rajalakshmi, G. Microbial lipases: An overview of screening, production and purification. Biocatal. Agric. Biotechnol. 2019, 22, 101368. [Google Scholar] [CrossRef]
- de Souza, C.E.C.; Ribeiro, B.D.; Coelho, M.A.Z. Characterization and Application of Yarrowia lipolytica Lipase Obtained by Solid-State Fermentation in the Synthesis of Different Esters Used in the Food Industry. Appl. Biochem. Biotechnol. 2019, 189, 933–959. [Google Scholar] [CrossRef]
- Lopes, M.; Miranda, S.M.; Alves, J.M.; Pereira, A.S.; Belo, I. Waste Cooking Oils as Feedstock for Lipase and Lipid-Rich Biomass Production. Eur. J. Lipid Sci. Technol. 2019, 121, 1800188. [Google Scholar] [CrossRef] [Green Version]
- Amaral, P.F.F.; Freire, M.G.; Rocha-Leao, M.H.M.; Marrucho, I.M.; Coutinho, J.A.P.; Coelho, M.A.Z. Optimization of oxygen mass transfer in a multiphase bioreactor with perfluorodecalin as a second liquid phase. Biotechnol. Bioeng. 2008, 99, 588–598. [Google Scholar] [CrossRef] [PubMed]
- Zieniuk, B.; Mazurczak-Zieniuk, P.; Fabiszewska, A. Exploring the Impact of Lipid-Rich Food Industry Waste Carbon Sources on the Growth of Candida cylindracea DSM 2031. Fermentation 2020, 6, 122. [Google Scholar] [CrossRef]
- Yan, J.; Han, B.; Gui, X.; Wang, G.; Xu, L.; Yan, Y.; Madzak, C.; Pan, D.; Wang, Y.; Zha, G.; et al. Engineering Yarrowia lipolytica to Simultaneously Produce Lipase and Single Cell Protein from Agro-industrial Wastes for Feed. Sci. Rep. 2018, 8, 758. [Google Scholar] [CrossRef] [Green Version]
- Sarris, D.; Sampani, Z.; Rapti, A.; Papanikolaou, S. Valorization of Crude Glycerol, Residue Deriving from Biodiesel-Production Process, with the Use of Wild-type New Isolated Yarrowia lipolytica Strains: Production of Metabolites with Pharmaceutical and Biotechnological Interest. Curr. Pharm. Biotechnol. 2019, 20, 881–894. [Google Scholar] [CrossRef] [PubMed]
- Sarris, D.; Stoforos, N.G.; Mallouchos, A.; Kookos, I.K.; Koutinas, A.A.; Aggelis, G.; Papanikolaou, S. Production of added-value metabolites by Yarrowia lipolytica growing in olive mill wastewater-based media under aseptic and non-aseptic conditions. Eng. Life Sci. 2017, 17, 695–709. [Google Scholar] [CrossRef] [Green Version]
- Zieniuk, B.; Fabiszewska, A.; Białecka-Florjańczyk, E. Screening of solvents for favoring hydrolytic activity of Candida antarctica Lipase B. Bioproc. Biosyst. Eng. 2020, 43, 605–613. [Google Scholar] [CrossRef]
- Fabiszewska, A.U.; Zieniuk, B.; Kozłowska, M.; Mazurczak-Zieniuk, P.M.; Wołoszynowska, M.; Misiukiewicz-Stępień, P.; Nowak, D. Studies on Upgradation of Waste Fish Oil to Lipid-Rich Yeast Biomass in Yarrowia lipolytica Batch Cultures. Foods 2021, 10, 436. [Google Scholar] [CrossRef]
- Kapturowska, A.; Stolarzewicz, I.; Krzyczkowska, J.; Białecka-Florjańczyk, E. Studies on lipolytic activity of sonicated enzymes from Yarrowia lipolytica. Ultrason. Sonochem. 2012, 19, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Dervishi, F.; Destain, J.; Nahvi, I.; Thonart, P.; Zarkesh-Esfahani, H. High-level production of extracellular lipase by Yarrowia lipolytica mutants from methyl oleate. New Biotechnol. 2011, 28, 756–760. [Google Scholar] [CrossRef] [PubMed]
- López-Fernández, J.; Barrero, J.J.; Benaiges, M.D.; Valero, F. Truncated Prosequence of Rhizopus oryzae Lipase: Key Factor for Production Improvement and Biocatalyst Stability. Catalysts 2019, 9, 961. [Google Scholar] [CrossRef] [Green Version]
- Nooh, H.M.; Masomian, M.; Salleh, A.B.; Mohamad, R.; Ali, M.S.M.; Rahman, R.N.Z.R.A. Production of Thermostable T1 Lipase Using Agroindustrial Waste Medium Formulation. Catalysts 2018, 8, 485. [Google Scholar] [CrossRef] [Green Version]
- Corzo, G.; Revah, S. Production and characteristics of the lipase from Yarrowia lipolytica 681. Bioresour. Technol. 1999, 70, 173–180. [Google Scholar] [CrossRef]
- Vylkova, S.; Carmar, A.J.; Danhof, H.A.; Collete, J.R.; Zhou, H.; Lorenz, M.C. The fungal pathogen Candida albicans autoinduces hyphal morphogenesis by raising extracellular pH. mBio 2011, 2, e00055-11. [Google Scholar] [CrossRef] [Green Version]
- Vardar-Sukan, F. Foaming: Consequences, prevention and destruction. Biotechnol. Adv. 1998, 16, 913–948. [Google Scholar] [CrossRef]
- Bouchedja, D.N.; Danthine, S.; Kar, T.; Fickers, P.; Sassi, H.; Boudjellal, A.; Blecker, C.; Delvigne, F. pH level has a strong impact on population dynamics of the yeast Yarrowia lipolytica and oil micro-droplets in multiphasic bioreactor. FEMS Microbiol. Lett. 2018, 365, fny173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso, F.O.M.; Oliveira, E.B.L.; Dellamora-Ortiz, G.M.; Pereira-Meirelles, F.V. Improvement of lipase production at different stiring speeds and oxygen levels. Braz. J. Chem. Eng. 2005, 22, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Stolarzewicz, I.A.; Zaborniak, P.; Fabiszewska, A.U.; Białecka-Florjańczyk, E. Study on the properties of immobilized biocatalysts with lipase activity produced by Yarrowia lipolytica in batch culture. Chem. Biochem. Eng. Q. 2017, 31, 251–259. [Google Scholar] [CrossRef]
- Shu, Z.Y.; Jiang, H.; Lin, R.F.; Jiang, Y.M.; Lin, L.; Huang, J.Z. Technical methods to improve yield, activity and stability in the development of microbial lipases. J. Mol. Catal. B Enzym. 2010, 62, 1–8. [Google Scholar] [CrossRef]
- Najjar, A.; Robert, S.; Guerin, C.; Violet-Asther, M.; Carriere, F. Quantitative study of lipase secretion, extracellular lipolysis, and lipid storage in the yeast Yarrowia lipolytica grown in the presence of olive oil: Analogies with lipolysis in humans. Appl. Microbiol. Biotechnol. 2011, 89, 1947–1962. [Google Scholar] [CrossRef] [PubMed]
- Jahic, M.; Gustavsson, M.; Jansen, A.K.; Martinelle, M.; Enfors, S.O. Analysis and control of proteolysis of a fusion protein in Pichia pastoris fed-batch processes. J. Biotechnol. 2003, 102, 45–53. [Google Scholar] [CrossRef]
- Lopes, M.; Gomes, N.; Mota, M.; Belo, I. Yarrowia lipolytica Growth Under Increased Air Pressure: Influence on Enzyme Production. Appl. Biochem. Biotechnol. 2009, 159, 46–53. [Google Scholar] [CrossRef] [Green Version]
- Kaur, P.; Ghoshal, G.; Jain, A. Bio-utilization of fruits and vegetables waste to produce β-carotene in solid-state fermentation: Characterization and antioxidant activity. Process. Biochem. 2019, 76, 155–164. [Google Scholar] [CrossRef]
- Morgunov, I.G.; Kamzolova, S.V.; Lunina, J.N. Citric Acid Production by Yarrowia lipolytica Yeast on Different Renewable Raw Materials. Fermentation 2018, 4, 36. [Google Scholar] [CrossRef] [Green Version]
- Sarris, D.; Rapti, A.; Papafotis, N.; Koutinas, A.A.; Papanikolaou, S. Production of Added-Value Chemical Compounds through Bioconversions of Olive-Mill Wastewaters Blended with Crude Glycerol by a Yarrowia lipolytica Strain. Molecules 2019, 24, 222. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
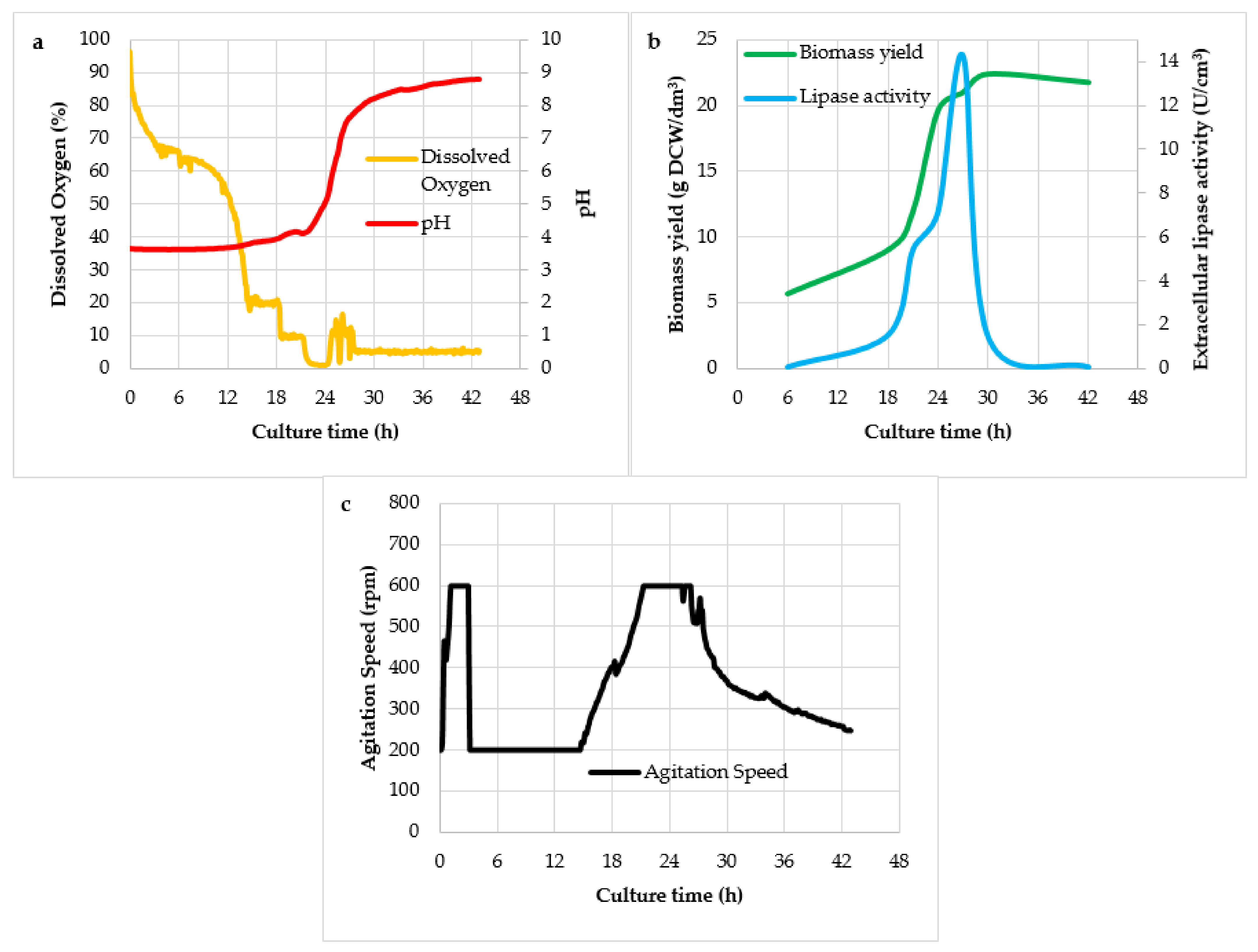{kind=link}
{kind=link}
| Culture No. | Mark of Culture | Agitator Speed (rpm) | Air Flow (dm3/(dm3 × min)) |
|---|---|---|---|
| 1 | 200 C 0.375 | 200 | 0.375 |
| 2 | 400 C 1.0 | 400 | 1.0 |
| 3 | 400 C 2.0 | 400 | 2.0 |
| 4 | 600 C 2.0 | 600 | 2.0 |
| 5 | (200–600) V 1.75 | Variable agitator speed dependent on the medium aeration (200–600) | 1.750 |
| Culture Time (h) | Set Aeration Level (%) |
|---|---|
| 0.0–3.0 | 80 |
| 3.0–6.5 | 60 |
| 6.5–12.0 | 40 |
| 12.0–18.5 | 20 |
| 18.5–27.5 | 10 |
| Above 27.5 | 5 |
| Culture Variant | Y(P/X) (U/g)1 | Y(X/S) (g/g) | qp (U/g × h) | vp (U/dm3 × h) |
|---|---|---|---|---|
| 200 C 0.375 | 37.50 | 0.16 | 1.44 | 4.73 |
| 400 C 1.0 | 134.58 | 0.78 | 5.61 | 86.92 |
| 400 C 2.0 | 290.55 | 0.89 | 10.76 | 191.22 |
| 600 C 2.0 | 205.85 | 1.19 | 6.86 | 163.10 |
| (200–600) V 1.75 | 677.41 | 1.05 | 25.09 | 526.37 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Snopek, P.; Nowak, D.; Zieniuk, B.; Fabiszewska, A. Aeration and Stirring in Yarrowia lipolytica Lipase Biosynthesis during Batch Cultures with Waste Fish Oil as a Carbon Source. Fermentation 2021, 7, 88. https://doi.org/10.3390/fermentation7020088
Snopek P, Nowak D, Zieniuk B, Fabiszewska A. Aeration and Stirring in Yarrowia lipolytica Lipase Biosynthesis during Batch Cultures with Waste Fish Oil as a Carbon Source. Fermentation. 2021; 7(2):88. https://doi.org/10.3390/fermentation7020088
Chicago/Turabian StyleSnopek, Paulina, Dorota Nowak, Bartłomiej Zieniuk, and Agata Fabiszewska. 2021. "Aeration and Stirring in Yarrowia lipolytica Lipase Biosynthesis during Batch Cultures with Waste Fish Oil as a Carbon Source" Fermentation 7, no. 2: 88. https://doi.org/10.3390/fermentation7020088
APA StyleSnopek, P., Nowak, D., Zieniuk, B., & Fabiszewska, A. (2021). Aeration and Stirring in Yarrowia lipolytica Lipase Biosynthesis during Batch Cultures with Waste Fish Oil as a Carbon Source. Fermentation, 7(2), 88. https://doi.org/10.3390/fermentation7020088