
Anti-Osteoporotic Activity of Pueraria lobata Fermented with Lactobacillus paracasei JS1 by Regulation of Osteoblast Differentiation and Protection against Bone Loss in Ovariectomized Mice


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Bacterial Culture and Fermentation
2.3. Metabolite Profiling of FPE Using Liquid Chromatography-Mass Spectrometry Analysis
2.4. Cell Culture
2.5. Osteoblast Proliferation Assay
2.6. Alkaline Phosphatase Assay
2.7. Real-Time PCR
2.8. Osteoclast Differentiation Assay
2.9. Animals
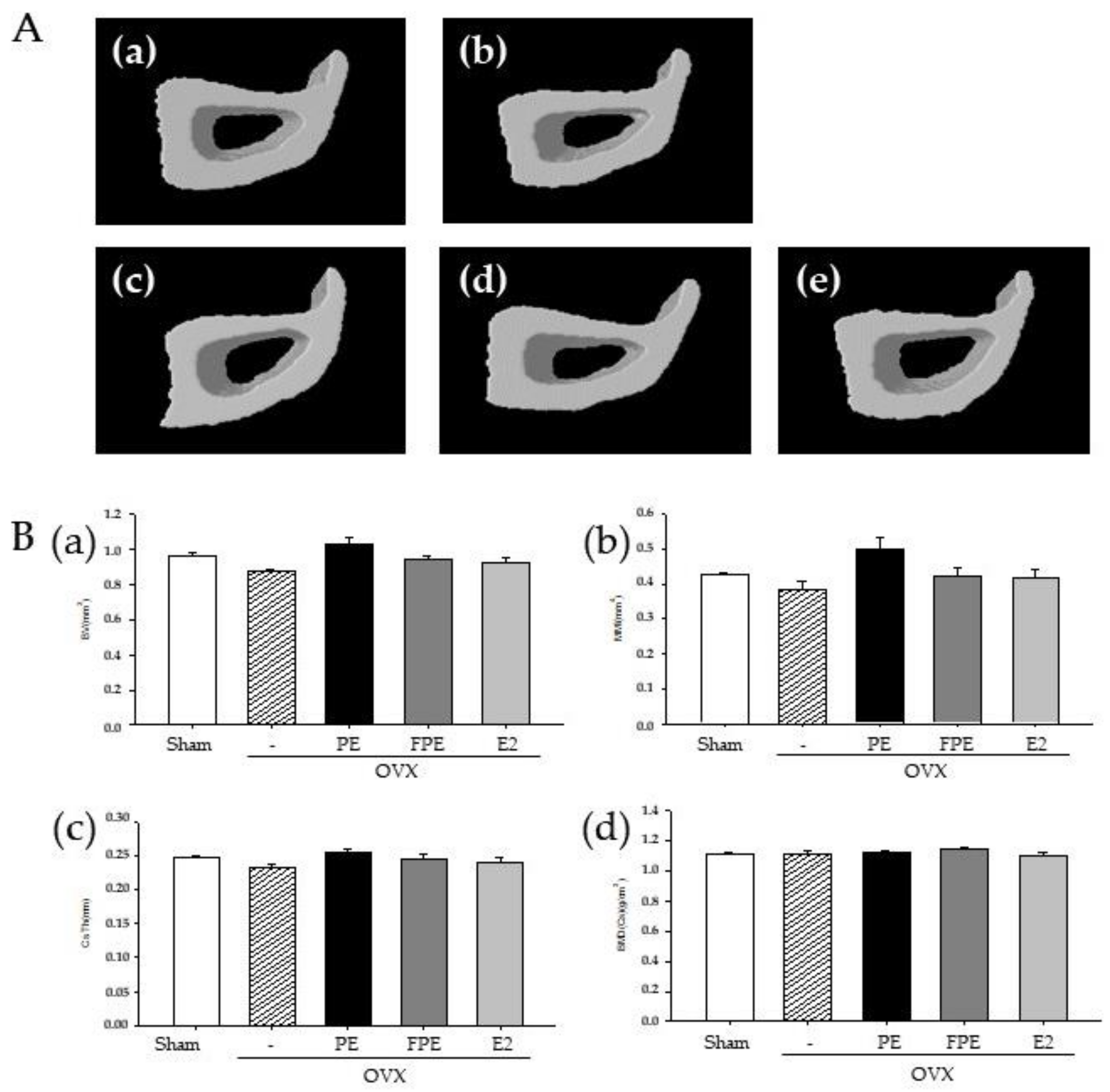
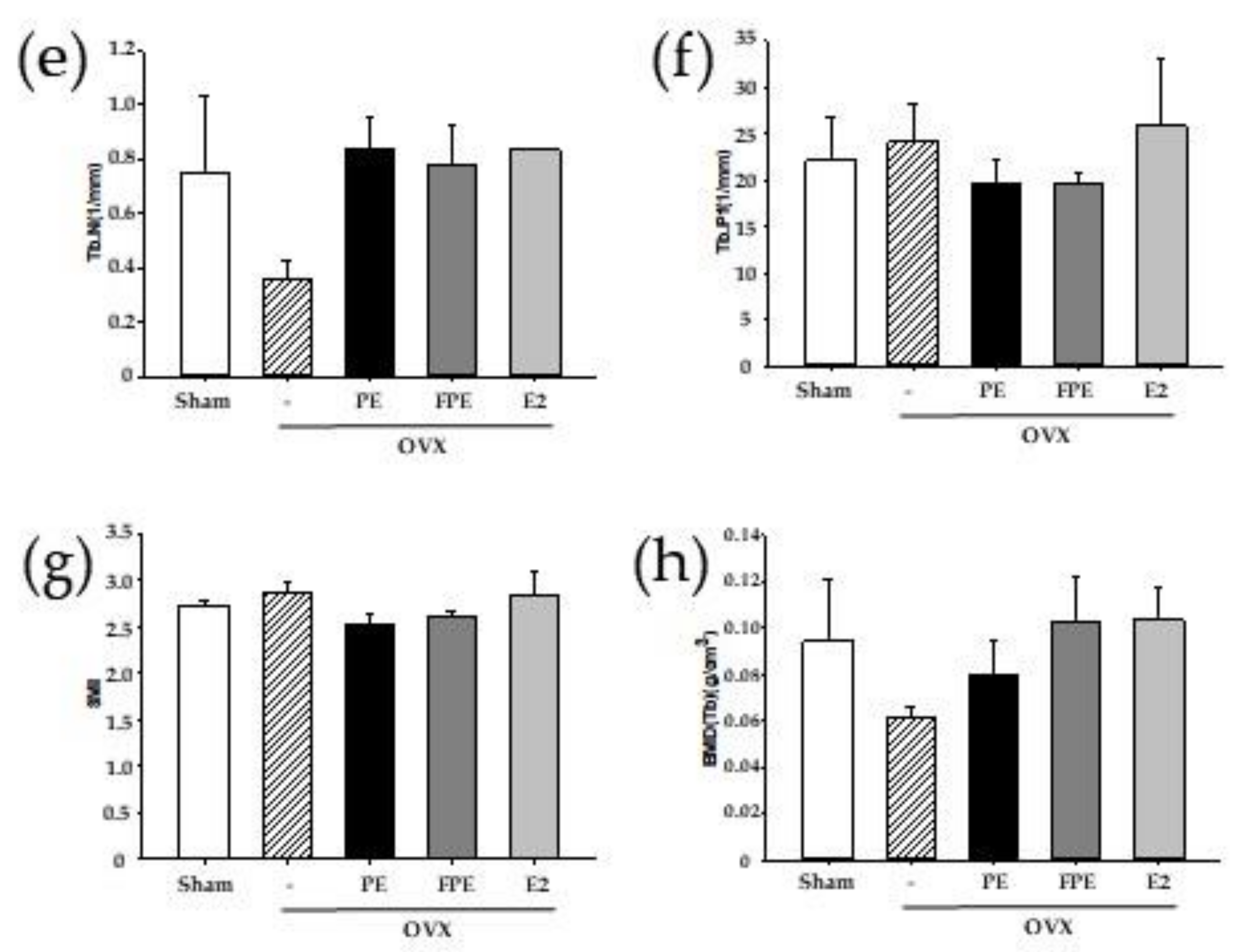
2.10. Micro-CT Bone Analysis
2.11. Serum Analysis
2.12. Statistical Analysis
3. Results and Discussion
3.1. Identification of Isoflavones in FPE
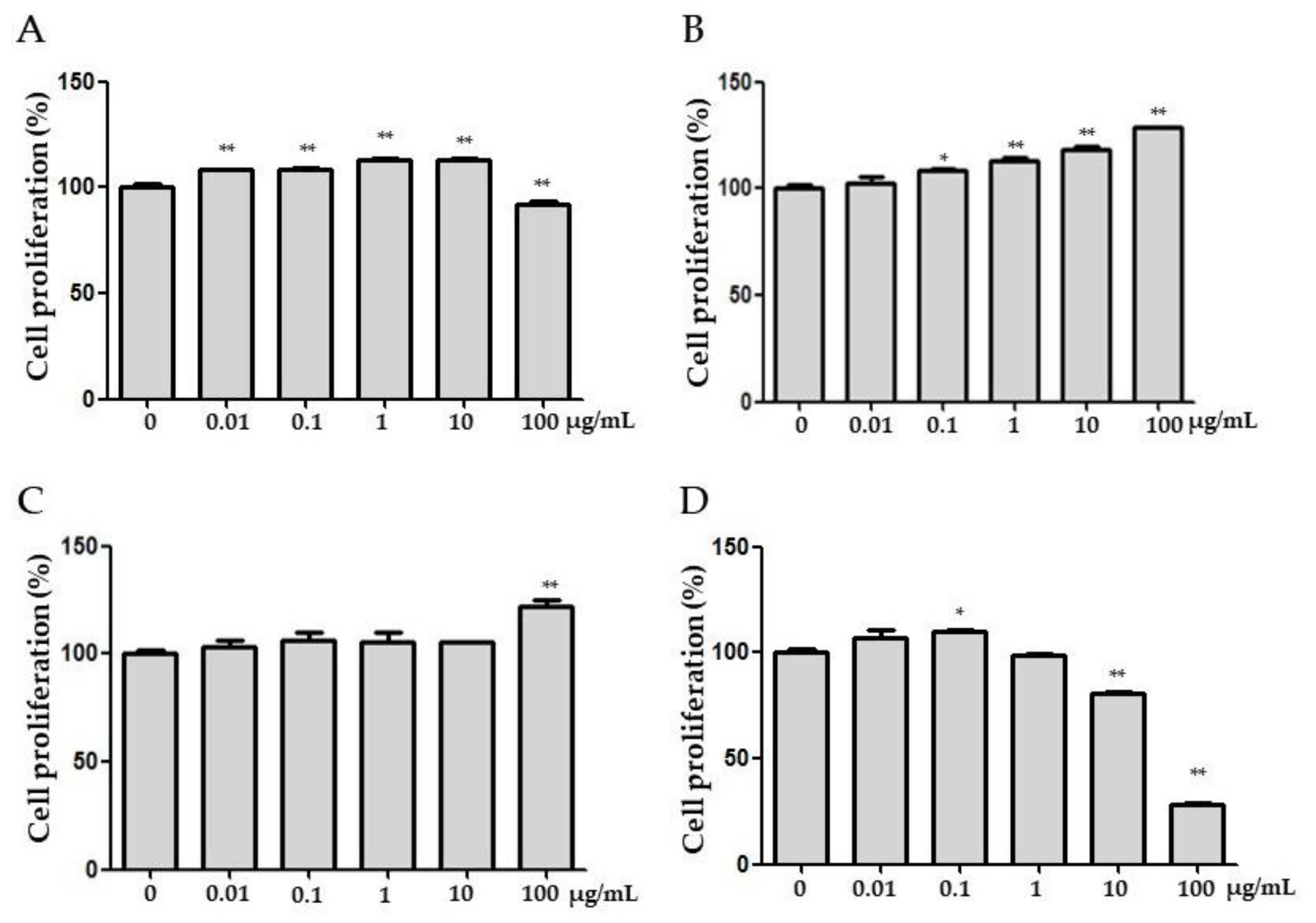
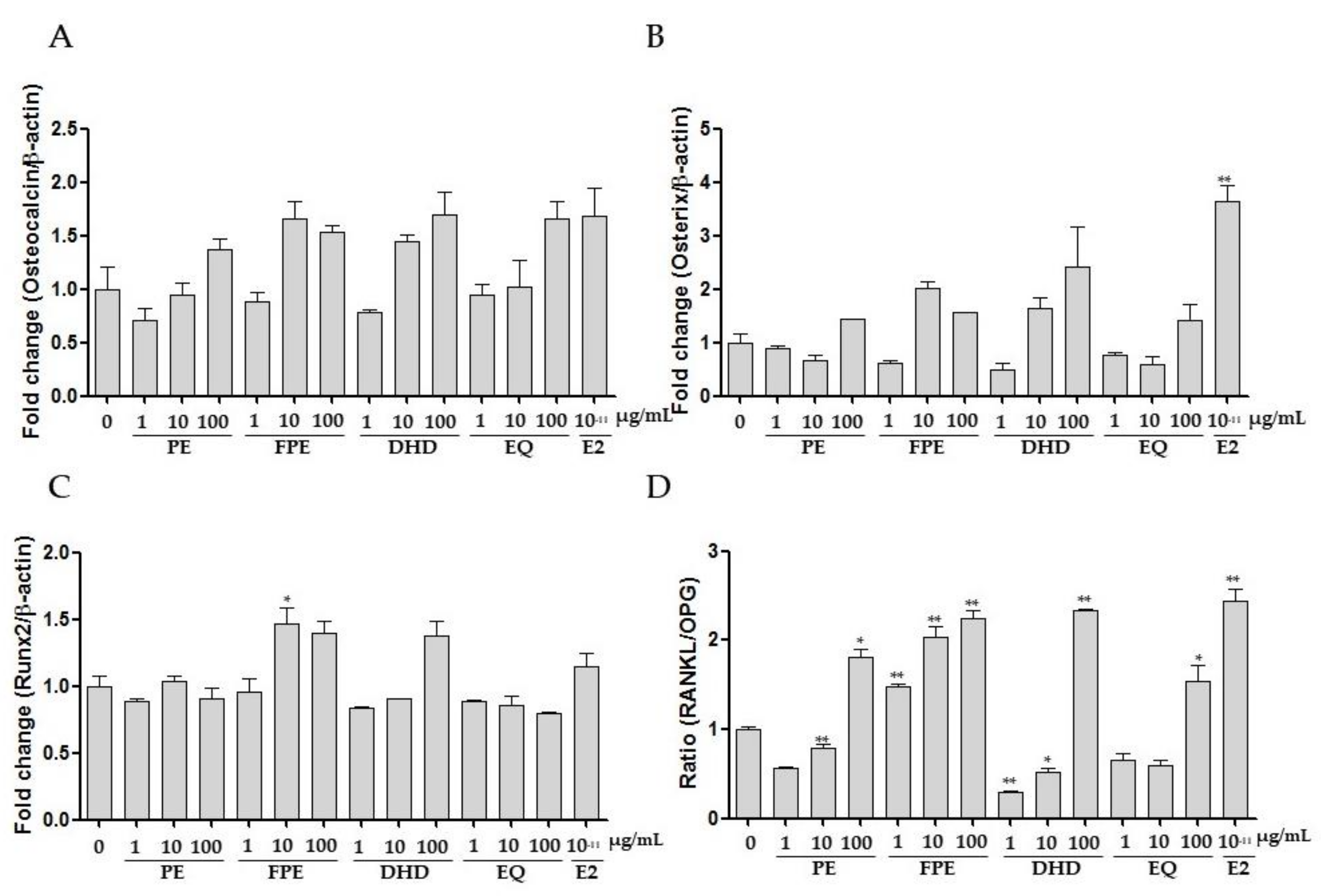
3.2. Effects of FPE on Proliferation, ALP Activities, and Gene Expression in MG 63 Cells
3.3. Effect of FPE on Osteoclast Differentiation in RAW 264.7 Cells
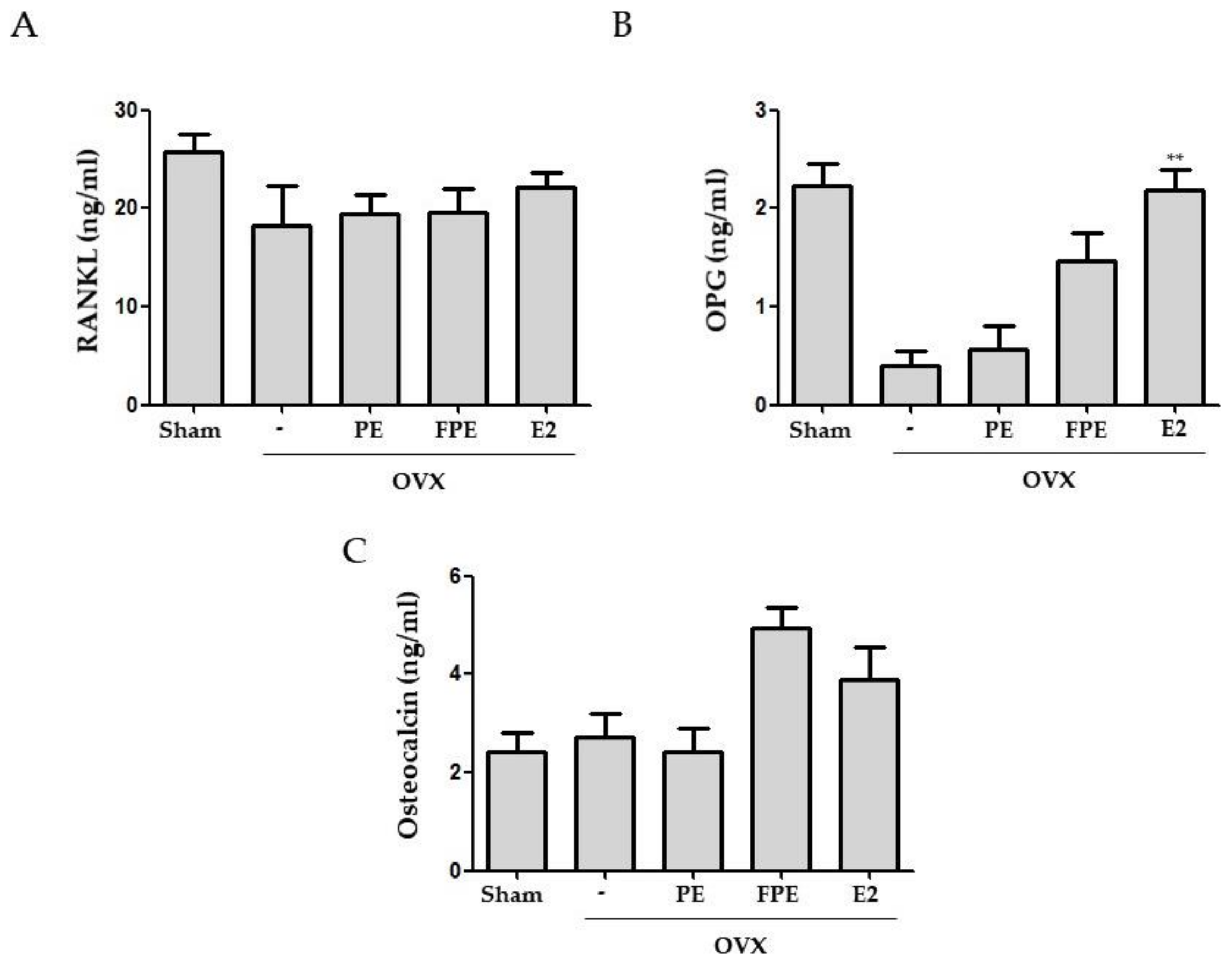
3.4. Effects of FPE in OVX-Induced Mouse Model
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Magee, P.J. Is equol production beneficial to health? Proc. Nutr. Soc. 2011, 70, 10–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, J.M.; Bab, I.; Fish, S.; Muller, R.; Uchiyama, T.; Gronowicz, G.; Nahounou, M.; Zhao, Q.; White, D.W.; Chorev, M.; et al. Human parathyroid hormone 1–34 reverses bone loss in ovariectomized mice. J. Bone Miner. Res. 2001, 16, 1665–1673. [Google Scholar] [CrossRef]
- Chung, H.J.; Cho, L.; Shin, J.S.; Lee, J.; Ha, I.H.; Park, H.J.; Lee, S.K. Effects of JSOG-6 on protection against bone loss in ovariectomized mice through regulation of osteoblast differentiation and osteoclast formation. BMC Complement. Altern. Med. 2014, 14, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raisz, L.G. Pathogenesis of osteoporosis: Concepts, conflicts, and prospects. J. Clin. Investig. 2005, 115, 3318–3325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.S.; Lu, C.H.; Chu, K.A.; Yeh, C.C.; Chiang, T.L.; Ko, T.L.; Chiu, M.M.; Chen, C.F. Xenograft of Human Umbilical Mesenchymal Stem Cells from Wharton’s Jelly Differentiating into Osteocytes and Reducing Osteoclast Activity Reverses Osteoporosis in Ovariectomized Rats. Cell Transplant. 2018, 27, 194–208. [Google Scholar] [CrossRef]
- Seeman, E. Invited Review: Pathogenesis of osteoporosis. J. Appl. Physiol. 2003, 95, 2142–2151. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.; Aryal, A.C.S.; Marini, J.C. Osteogenesis imperfecta: New genes reveal novel mechanisms in bone dysplasia. Transl. Res. 2017, 181, 27–48. [Google Scholar] [CrossRef]
- Meunier, P.J. Hyperthyroidism and osteoporosis. Ann. Endocrinol. 1995, 56, 57–59. [Google Scholar] [PubMed]
- Redlich, K.; Smolen, J.S. Inflammatory bone loss: Pathogenesis and therapeutic intervention. Nat. Rev. Drug Discov. 2012, 11, 234–250. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Qin, L.; Hung, W.Y.; Shi, Y.Y.; Leung, P.C.; Yeung, H.Y.; Leung, K.S. Flavonoids derived from herbal Epimedium Brevicornum Maxim prevent OVX-induced osteoporosis in rats independent of its enhancement in intestinal calcium absorption. Bone 2006, 38, 818–825. [Google Scholar] [CrossRef] [PubMed]
- Dionyssiotis, Y.; Skarantavos, G.; Papagelopoulos, P. Modern rehabilitation in osteoporosis, falls, and fractures. Clin. Med. Insights Arthritis Musculoskelet. Disord. 2014, 7, 33–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, S.; Crockett, J.C. Osteoporosis—A current view of pharmacological prevention and treatment. Drug Des. Dev. Ther. 2013, 7, 435–448. [Google Scholar] [CrossRef] [Green Version]
- Chow, T.H.; Lee, B.Y.; Ang, A.B.F.; Cheung, V.Y.K.; Ho, M.M.C.; Takemura, S. The effect of Chinese martial arts Tai Chi Chuan on prevention of osteoporosis: A systematic review. J. Orthop. Transl. 2018, 12, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Lee, S.K.; Chun, O.K. Soy Isoflavones and Osteoporotic Bone Loss: A Review with an Emphasis on Modulation of Bone Remodeling. J. Med. Food 2016, 19, 1–14. [Google Scholar] [CrossRef]
- Hickman, J.E.; Wu, S.; Mickley, L.J.; Lerdau, M.T. Kudzu (Pueraria montana) invasion doubles emissions of nitric oxide and increases ozone pollution. Proc. Natl. Acad. Sci. USA 2010, 107, 10115–10119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koirala, P.; Seong, S.H.; Jung, H.A.; Choi, J.S. Comparative molecular docking studies of lupeol and lupenone isolated from Pueraria lobata that inhibits BACE1: Probable remedies for Alzheimer’s disease. Asian Pac. J. Trop. Med. 2017, 10, 1117–1122. [Google Scholar] [CrossRef] [PubMed]
- Keung, W.M.; Vallee, B.L. Kudzu root: An ancient Chinese source of modern antidipsotropic agents. Phytochemistry 1998, 47, 499–506. [Google Scholar] [CrossRef]
- Gao, Y.; Wang, X.; He, C. An isoflavonoid-enriched extract from Pueraria lobata (kudzu) root protects human umbilical vein endothelial cells against oxidative stress induced apoptosis. J. Ethnopharmacol. 2016, 193, 524–530. [Google Scholar] [CrossRef]
- Jin, S.E.; Son, Y.K.; Min, B.S.; Jung, H.A.; Choi, J.S. Anti-inflammatory and antioxidant activities of constituents isolated from Pueraria lobata roots. Arch. Pharm. Res. 2012, 35, 823–837. [Google Scholar] [CrossRef]
- Seong, S.H.; Roy, A.; Jung, H.A.; Jung, H.J.; Choi, J.S. Protein tyrosine phosphatase 1B and alpha-glucosidase inhibitory activities of Pueraria lobata root and its constituents. J. Ethnopharmacol. 2016, 194, 706–716. [Google Scholar] [CrossRef]
- Cheung, D.W.; Koon, C.M.; Wat, E.; Ko, C.H.; Chan, J.Y.; Yew, D.T.; Leung, P.C.; Chan, W.Y.; Lau, C.B.; Fung, K.P. A herbal formula containing roots of Salvia miltiorrhiza (Danshen) and Pueraria lobata (Gegen) inhibits inflammatory mediators in LPS-stimulated RAW 264.7 macrophages through inhibition of nuclear factor kappaB (NFkappaB) pathway. J. Ethnopharmacol. 2013, 145, 776–783. [Google Scholar] [CrossRef] [PubMed]
- Park, K.H.; Gu, D.R.; Jin, S.H.; Yoon, C.S.; Ko, W.; Kim, Y.C.; Lee, S.H. Pueraria lobate Inhibits RANKL-Mediated Osteoclastogenesis Via Downregulation of CREB/PGC1beta/c-Fos/NFATc1 Signaling. Am. J. Chin. Med. 2017, 45, 1725–1744. [Google Scholar] [CrossRef] [PubMed]
- Huh, J.E.; Yang, H.R.; Park, D.S.; Choi, D.Y.; Baek, Y.H.; Cho, E.M.; Cho, Y.J.; Kang-Il, K.; Kim, D.Y.; Lee, J.D. Puerariae radix promotes differentiation and mineralization in human osteoblast-like SaOS-2 cells. J. Ethnopharmacol. 2006, 104, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Hessle, C.; Hanson, L.A.; Wold, A.E. Lactobacilli from human gastrointestinal mucosa are strong stimulators of IL-12 production. Clin. Exp. Immunol. 1999, 116, 276–282. [Google Scholar] [CrossRef]
- Molin, G.; Jeppsson, B.; Johansson, M.L.; Ahrne, S.; Nobaek, S.; Stahl, M.; Bengmark, S. Numerical taxonomy of Lactobacillus spp. associated with healthy and diseased mucosa of the human intestines. J. Appl. Bacteriol. 1993, 74, 314–323. [Google Scholar] [CrossRef]
- Kwon, J.E.; Lim, J.; Bang, I.; Kim, I.; Kim, D.; Kang, S.C. Fermentation product with new equol-producing Lactobacillus paracasei as a probiotic-like product candidate for prevention of skin and intestinal disorder. J. Sci. Food Agric. 2019, 99, 4200–4210. [Google Scholar] [CrossRef]
- Matthies, A.; Loh, G.; Blaut, M.; Braune, A. Daidzein and Genistein Are Converted to Equol and 5-Hydroxy-Equol by Human Intestinal Slackia isoflavoniconvertens in Gnotobiotic Rats. Nutr. J. 2012, 142, 40–46. [Google Scholar] [CrossRef]
- Xiao, L.; Zhong, M.; Huang, Y. Puerarin alleviates osteoporosis in the ovariectomy-induced mice by suppressing osteoclastogenesis via inhibition of TRAF6/ROS-dependent MAPK/NF-κB signaling pathways. Aging 2020, 12, 21706–21729. [Google Scholar] [CrossRef]
- An, J.; Yang, H.; Zhang, Q. Natural products for treatment of osteoporosis: The effects and mechanisms on promoting osteoblast-mediated bone formation. Life Sci. 2016, 147, 46–58. [Google Scholar] [CrossRef]
- Zou, S.E.; Zhang, S.F.; Zhang, R.; Zhang, J. Role of the cross-talk between estrogen receptors and peroxisome proliferator-activated receptor gamma in daidzein’s prevention and treatment of osteoporosis in ovariectomized rats. Zhonghua Yi Xue Za Zhi 2009, 89, 2972–2975. [Google Scholar] [PubMed]
- Li, N.; Tu, Y.; Shen, Y.; Qin, Y.; Lei, C.; Liu, X. Calycosin attenuates osteoporosis and regulates the expression of OPG/RANKL in ovariectomized rats via MAPK signaling. Pharmazie 2016, 71, 607–612. [Google Scholar] [CrossRef]
- Fujioka, M.; Uehara, M.; Wu, J. Equol, a metabolite of daidzein, inhibits bone loss in ovariectomized mice. J. Nutr. 2004, 134, 2623–2627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katagiri, T.; Takahashi, N. Regulatory mechanisms of osteoblast and osteoclast differentiation. Oral. Dis. 2002, 8, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Frolik, C.A.; Bryant, H.U.; Black, E.C.; Magee, D.E.; Chandrasekhar, S. Time-dependent changes in biochemical bone markers and serum cholesterol in ovariectomized rats: Effects of raloxifene HCl, tamoxifen, estrogen, and alendronate. Bone 1996, 18, 621–627. [Google Scholar] [CrossRef]
- Cano, A.; Dapia, S.; Noguera, I.; Pineda, B.; Hermenegildo, C.; del Val, R.; Caeiro, J.R.; Garcia-Perez, M.A. Comparative effects of 17beta-estradiol, raloxifene and genistein on bone 3D microarchitecture and volumetric bone mineral density in the ovariectomized mice. Osteoporos. Int. 2008, 19, 793–800. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Genes | 5′ to 3′ | Sequence |
|---|---|---|
| RANKL | Forward | ATGGCGTCCTCTCTGCTTG |
| Reverse | TGAAAGGTCAGCGTATGGCTT | |
| Runx2 | Forward | CCGGTCTCCTTCCAGGAT |
| Reverse | GGGAACTGCTGTGGCTTC | |
| Osteocalcin | Forward | CCGGTCTCCTTCCAGGAT |
| Reverse | GGGAACTGCTGTGGCTTC | |
| Osterix | Forward | ATGGCGTCCTCTCTGCTTG |
| Reverse | TGAAAGGTCAGCGTATGGCTT | |
| β-actin | Forward | CGCTGATGCATGCCTATGA |
| Reverse | AGAGGTCCACAGAGCTGATTCC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.Y.; Lee, H.-J.; Kim, T.; Lee, Y.-G.; Kwon, J.E.; Kang, S.C. Anti-Osteoporotic Activity of Pueraria lobata Fermented with Lactobacillus paracasei JS1 by Regulation of Osteoblast Differentiation and Protection against Bone Loss in Ovariectomized Mice. Fermentation 2021, 7, 186. https://doi.org/10.3390/fermentation7030186
Kim SY, Lee H-J, Kim T, Lee Y-G, Kwon JE, Kang SC. Anti-Osteoporotic Activity of Pueraria lobata Fermented with Lactobacillus paracasei JS1 by Regulation of Osteoblast Differentiation and Protection against Bone Loss in Ovariectomized Mice. Fermentation. 2021; 7(3):186. https://doi.org/10.3390/fermentation7030186
Chicago/Turabian StyleKim, Seon Yu, Hee-Ju Lee, Taehyun Kim, Yeong-Geun Lee, Jeong Eun Kwon, and Se Chan Kang. 2021. "Anti-Osteoporotic Activity of Pueraria lobata Fermented with Lactobacillus paracasei JS1 by Regulation of Osteoblast Differentiation and Protection against Bone Loss in Ovariectomized Mice" Fermentation 7, no. 3: 186. https://doi.org/10.3390/fermentation7030186
APA StyleKim, S. Y., Lee, H. -J., Kim, T., Lee, Y. -G., Kwon, J. E., & Kang, S. C. (2021). Anti-Osteoporotic Activity of Pueraria lobata Fermented with Lactobacillus paracasei JS1 by Regulation of Osteoblast Differentiation and Protection against Bone Loss in Ovariectomized Mice. Fermentation, 7(3), 186. https://doi.org/10.3390/fermentation7030186