
Agro-Industrial Wastes: A Substrate for Multi-Enzymes Production by Cryphonectria parasitica


Abstract
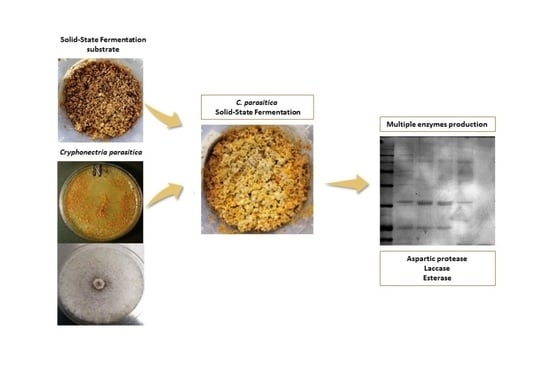
:
1. Introduction
2. Materials and Methods
2.1. Microorganisms and Their Molecular Characterization
2.2. Agro-Food Waste Based Substrates and Solid State Fermentation
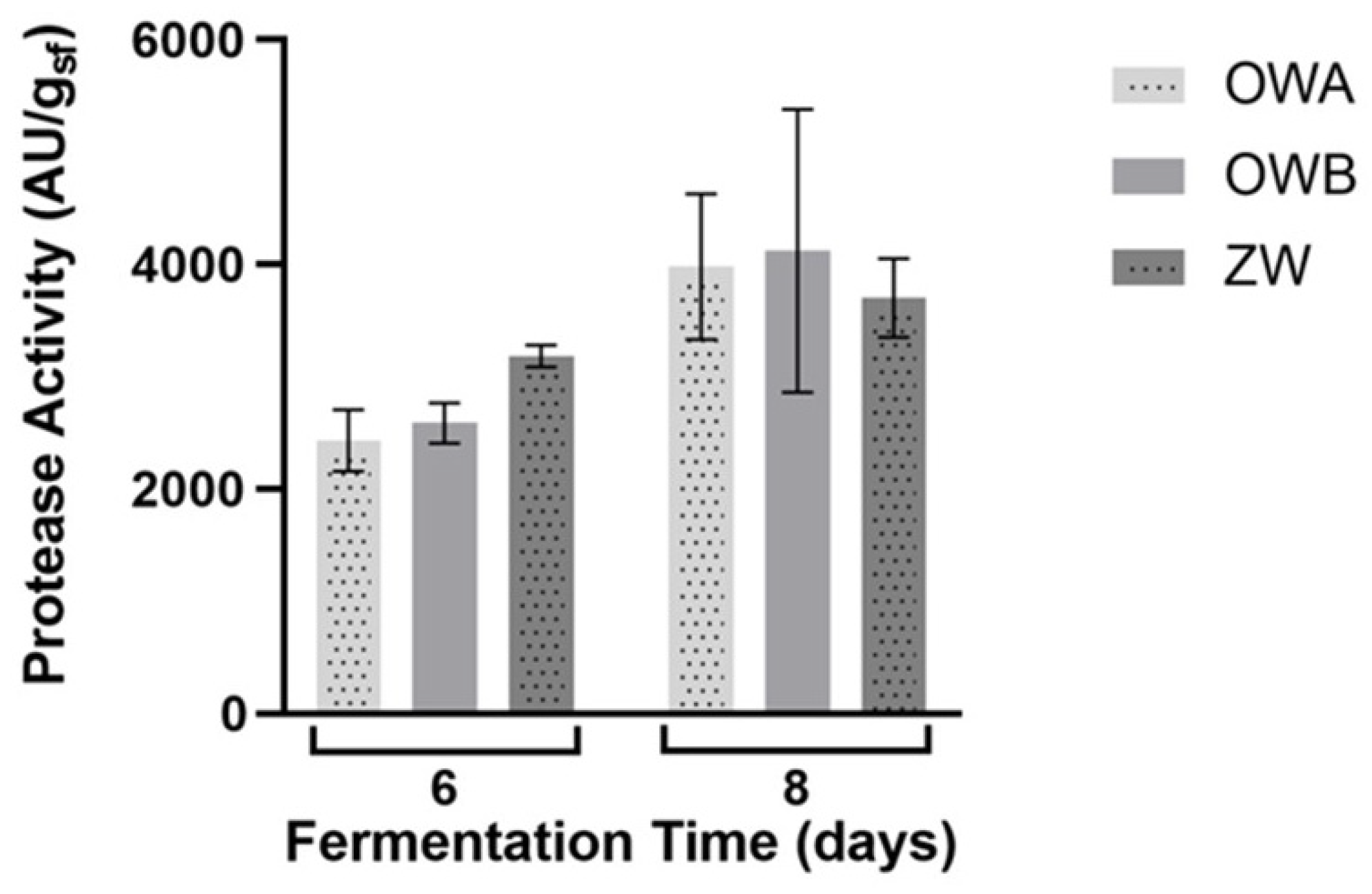
2.2.1. Effect of the Carbon Source on the Fungal Growth and Protease Activity
2.2.2. Effect of Water Content on the Fungal Growth and Protease Activity
2.2.3. Effect of Substrate Density on the Fungal Growth and Protease Activity
2.3. SSF Protocol Set Up
2.4. Extraction of Crude Enzyme from the Fermented Media
2.5. Protease Activity Assay
2.6. TBA Assay
2.7. Esterase Activity Assay
2.8. Laccase Activity Assay
2.9. SDS-PAGE
2.10. Protein Determination
2.11. Statistical Analysis
3. Results and Discussion
3.1. SSF Protocol Set Up
3.1.1. Fungal Growth on Different Raw Material
3.1.2. Protease Production under SSF
3.1.3. Enzymes Putatively Identification
3.2. Multi-Enzymes Production under SSF
3.2.1. Strains Characterization
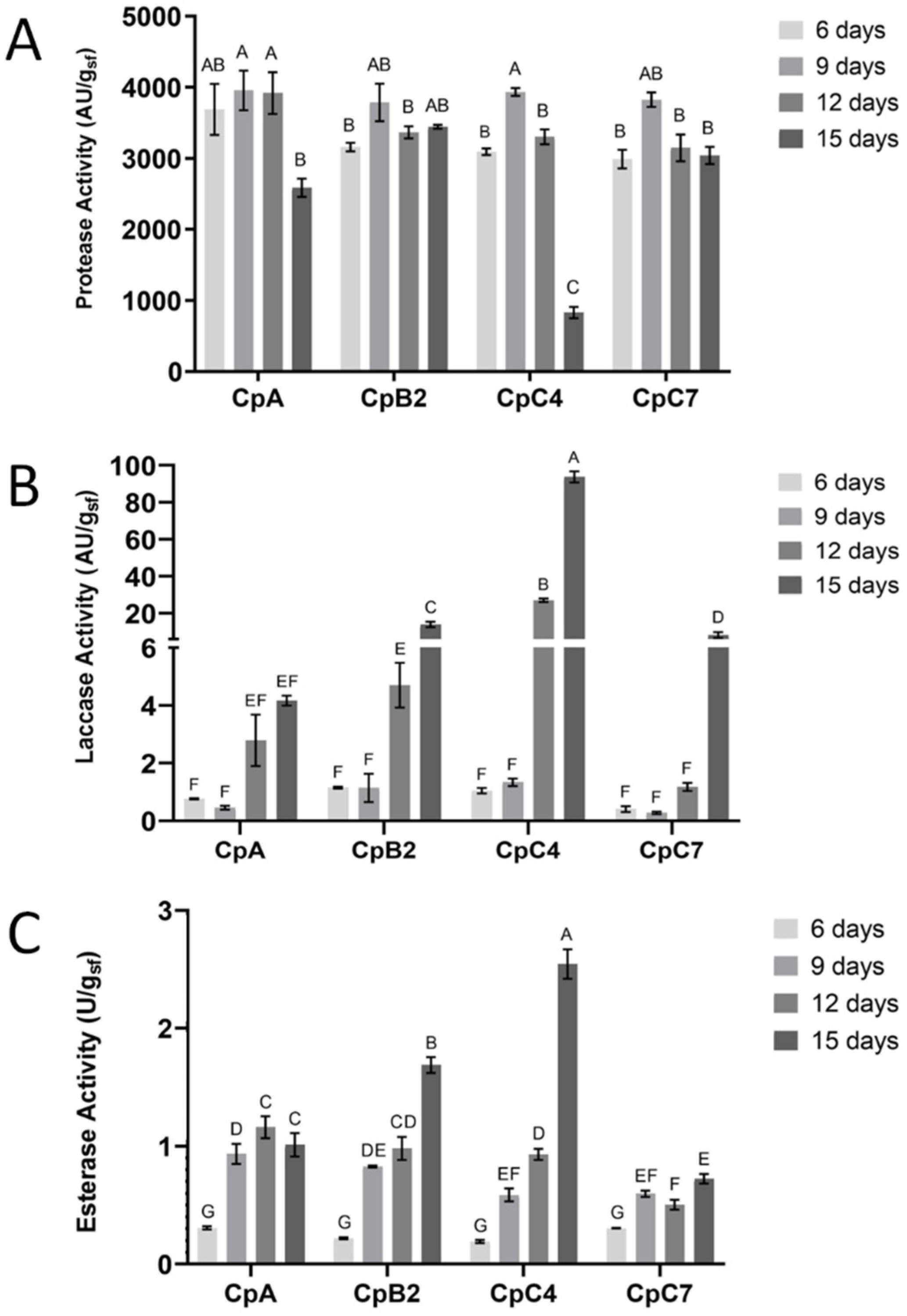
3.2.2. Multiple Enzymes Activities
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Bringezu, S.B.R. Sustainable Resource Management: Global Trends, Visions and Policies, 1st ed.; Taylor & Francis: New York, NY, USA, 2009. [Google Scholar]
- Ravindran, R.; Jaiswal, A.K. Exploitation of Food Industry Waste for High-Value Products. Trends Biotechnol. 2016, 34, 58–69. [Google Scholar] [CrossRef] [Green Version]
- Belewu, M.A.; Babalola, F.T. Nutrient enrichment of waste agricultural residues after solid state fermentation using Rhizopus oligosporus. J. Appl. Biosci. 2009, 13, 695–699. [Google Scholar]
- Sadh, P.K.; Duhan, S.; Duhan, J.S. Agro-industrial wastes and their utilization using solid state fermentation: A review. Bioresour. Bioprocess 2018, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Bajaj, B.K. Potential application spectrum of microbial proteases for clean and green industrial production. Energy Ecol. Environ. 2017, 2, 370–386. [Google Scholar] [CrossRef] [Green Version]
- Marzo, C.; Díaz, A.B.; Caro, I.; Blandino, A. Valorization of agro-industrial wastes to produce hydrolytic enzymes by fungal solid-state fermentation. Waste Manag. Res. 2018, 37, 149–156. [Google Scholar] [CrossRef]
- Alias, C.; Bulgari, D.; Bilo, F.; Borgese, L.; Gianoncelli, A.; Ribaudo, G.; Gobbi, E.; Alessandri, I. Food waste-assisted metal extraction from printed circuit boards: The aspergillus niger route. Microorganisms 2021, 9, 895. [Google Scholar] [CrossRef] [PubMed]
- Razzaq, A.; Shamsi, S.; Ali, A.; Ali, Q.; Sajjad, M.; Malik, A.; Ashraf, M. Microbial Proteases Applications. Front. Bioeng. Biotechnol. 2019, 7, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarnthima, R.; Khammuang, S.; Svasti, J. Extracellular ligninolytic enzymes by Lentinus polychrous Lév. under solid-state fermentation of potential agro-industrial wastes and their effectiveness in decolorization of synthetic dyes. Biotechnol. Bioprocess Eng. 2009, 14, 513–522. [Google Scholar] [CrossRef]
- Arregui, L.; Ayala, M.; Gómez-Gil, X.; Gutiérrez-Soto, G.; Hernández-Luna, C.E.; de los Santos, H.M.; Levin, L.; Rojo-Domínguez, A.; Romero-Martínez, D.; Saparrat, M.C.N.; et al. Laccases: Structure, function, and potential application in water bioremediation. Microb. Cell Fact. 2019, 18, 200. [Google Scholar] [CrossRef] [PubMed]
- Barriuso, J.; Prieto, A.; Martínez, M.J. Fungal genomes mining to discover novel sterol esterases and lipases as catalysts. BMC Genom. 2013, 14, 712. [Google Scholar] [CrossRef] [Green Version]
- Chandra, P.; Enespa; Singh, R.; Arora, P.K. Microbial lipases and their industrial applications: A comprehensive review. Microb. Cell Fact. 2020, 19, 169. [Google Scholar] [CrossRef]
- Diaz, A.B.; Blandino, A.; Caro, I. Value added products from fermentation of sugars derived from agro-food residues. Trends Food Sci. Technol. 2018, 71, 52–64. [Google Scholar] [CrossRef]
- Anitha, T.S.; Palanivelu, P. Purification and characterization of an extracellular keratinolytic protease from a new isolate of Aspergillus parasiticus. Protein Expr. Purif. 2013, 88, 214–220. [Google Scholar] [CrossRef]
- Ramos, O.S.; Malcata, F.X. 3.48—Food-Grade Enzymes. In Comprehensive Biotechnology, 2nd ed.; Moo-Young, M., Ed.; Academic Press: Burlington, VT, USA, 2011; pp. 555–569. ISBN 978-0-08-088504-9. [Google Scholar]
- Vandenberghe, L.P.S.; Pandey, A.; Carvalho, J.C.; Letti, L.A.J.; Woiciechowski, A.L.; Karp, S.G.; Thomaz-Soccol, V.; Martínez-Burgos, W.J.; Penha, R.O.; Herrmann, L.W.; et al. Solid-state fermentation technology and innovation for the production of agricultural and animal feed bioproducts. Syst. Microbiol. Biomanuf. 2021, 1, 142–165. [Google Scholar] [CrossRef]
- Soccol, C.R.; da Costa, E.S.F.; Letti, L.A.J.; Karp, S.G.; Woiciechowski, A.L.; de Souza, V.L.P. Recent developments and innovations in solid state fermentation. Biotechnol. Res. Innov. 2017, 1, 52–71. [Google Scholar] [CrossRef]
- de Castro, R.J.S.; Sato, H.H. Enzyme Production by Solid State Fermentation: General Aspects and an Analysis of the Physicochemical Characteristics of Substrates for Agro-industrial Wastes Valorization. Waste Biomass Valorization 2015, 6, 1085–1093. [Google Scholar] [CrossRef]
- Yazid, N.A.; Barrena, R.; Komilis, D.; Sánchez, A. Solid-state fermentation as a novel paradigm for organic waste valorization: A review. Sustainability 2017, 9, 224. [Google Scholar] [CrossRef] [Green Version]
- López-Gómez, J.P.; Venus, J. Potential role of sequential solid-state and submerged-liquid fermentations in a circular bioeconomy. Fermentation 2021, 7, 76. [Google Scholar] [CrossRef]
- Webb, C. Design Aspects of Solid State Fermentation as Applied to Microbial Bioprocessing. J. Appl. Biotechnol. Bioeng. 2017, 4, 511–532. [Google Scholar] [CrossRef] [Green Version]
- Carboué, Q.; Tranier, M.-S.; Perraud-Gaime, I.; Roussos, S. Production of Microbial Enzymes by Solid-state Fermentation for Food Applications. In Microbial Enzyme Technology in Food Applications; Ramesh, C., Ray, C.M.R., Eds.; CRC Press: London, UK; Taylor & Francis Group Boca Raton: New York, NY, USA, 2017; ISBN 9781498749831. [Google Scholar]
- Kumla, J.; Suwannarach, N.; Sujarit, K.; Penkhrue, W.; Kakumyan, P.; Jatuwong, K.; Vadthanarat, S.; Lumyong, S. Cultivation of Mushrooms and Their Lignocellulolytic Enzyme Production Through the Utilization of Agro-Industrial Waste. Molecules 2020, 25, 2811. [Google Scholar] [CrossRef]
- Jatuwong, K.; Kumla, J.; Suwannarach, N.; Matsui, K.; Lumyong, S. Bioprocessing of Agricultural Residues as Substrates and Optimal Conditions for Phytase Production of Chestnut Mushroom, Pholiota adiposa, in Solid State Fermentation. J. Fungi 2020, 6, 384. [Google Scholar] [CrossRef]
- Ramírez, C.M.G.; Rivera-Ríos, J.M.; Téllez-Jurado, A.; Maqueda Gálvez, A.P.; Mercado-Flores, Y.; Arana-Cuenca, A. Screening for thermotolerant ligninolytic fungi with laccase, lipase, and protease activity isolated in Mexico. J. Environ. Manage. 2012, 95, S256–S259. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, R.S.; Sánchez, G.I.; Arévalo-Villena, M.; Pérez, B.A. Production and immobilization of enzymes by solid-state fermentation of agroindustrial waste. Bioprocess Biosyst. Eng. 2015, 38, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Viayaraghavan, P.; Kumar, J.S.; Arasu, V.M.; Al-Dhabi, N.A. Simultaneous production of commercial enzymes using agro industrial residues by statistical approach. J. Sci. Food Agric. 2019, 99, 2685–2696. [Google Scholar] [CrossRef] [PubMed]
- McCarroll, D.R.; Thor, E. Pectolytic, cellulytic and proteolytic activities expressed by cultures of Endothia parasitica, and inhibition of these activities by components extracted from Chinese and American chestnut inner bark. Physiol. Plant Pathol. 1985, 26, 367–378. [Google Scholar] [CrossRef]
- Varley, D.A.; Podila, G.K.; Hiremath, S.T. Cutinase in Cryphonectria parasitica, the chestnut blight fungus: Suppression of cutinase gene expression in isogenic hypovirulent strains containing double-stranded RNAs. Mol. Cell. Biol. 1992, 12, 4539–4544. [Google Scholar] [CrossRef] [PubMed]
- Farias, G.M.; Gorbea, C.; Elkins, J.R.; Griffin, G.J. Purification, characterization, and substrate relationships of the tannase from Cryphonectria parasitica. Physiol. Mol. Plant Pathol. 1994, 44, 51–63. [Google Scholar] [CrossRef]
- Chung, H.-J.; Kwon, B.-R.; Kim, J.-M.; Park, S.-M.; Park, J.-K.; Cha, B.-J.; Yang, M.-S.; Kim, D.-H. A tannic acid-inducible and hypoviral-regulated Laccase3 contributes to the virulence of the chestnut blight fungus Cryphonectria parasitica. Mol. Plant. Microbe. Interact. 2008, 21, 1582–1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FDA Microorganisms & Microbial-Derived Ingredients Used in Food (Partial List). Available online: https://www.fda.gov/food/generally-recognized-safe-gras/microorganisms-microbial-derived-ingredients-used-food-partial-list (accessed on 20 November 2021).
- Rigling, D.; Prospero, S. Cryphonectria parasitica, the causal agent of chestnut blight: Invasion history, population biology and disease control. Mol. Plant Pathol. 2018, 19, 7–20. [Google Scholar] [CrossRef] [Green Version]
- Hillman, B.I.; Suzuki, N. Viruses of the chestnut blight fungus, Cryphonectria parasitica. Adv. Virus Res. 2004, 63, 423–472. [Google Scholar] [CrossRef]
- Kazmierczak, P.; Pfeiffer, P.; Zhang, L.; Van Alfen, N.K. Transcriptional repression of specific host genes by the mycovirus Cryphonectria hypovirus 1. J. Virol. 1996, 70, 1137–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Lu, L.; Yang, Y.; Chen, Q.; Chen, B. Proteomic analysis of Cryphonectria parasitica infected by a virulence-attenuating hypovirus. Wei Sheng Wu Xue Bao 2014, 54, 803–812. [Google Scholar] [PubMed]
- Gobbi, E.; Rekab, D.; Locci, R. Mitochondrial plasmids of the pCp family are spread worldwide in Cryphonectria parasitica populations. Mycol. Res. 2002, 106, 1408–1416. [Google Scholar] [CrossRef]
- Lione, G.; Giordano, L.; Turina, M.; Gonthier, P. Hail-Induced Infections of the Chestnut Blight Pathogen Cryphonectria parasitica Depend on Wound Size and May Lead to Severe Diebacks. Phytopathology 2020, 110, 1280–1293. [Google Scholar] [CrossRef]
- Anson, M.L. The estimation of pepsin, trypsin, papain, and cathepsin with hemoglobin. J. Gen. Physiol. 1938, 22, 79–89. [Google Scholar] [CrossRef]
- Tello-Solís, S.R.; Rodríguez-Romero, A.; Hernández-Arana, A. Circular dichroism studies of acid proteinases from Aspergillus niger and Aspergillus awamori. Biochem. Mol. Biol. Int. 1994, 33, 759–768. [Google Scholar]
- Vishwanatha, K.S.; Rao, A.G.A.; Singh, S.A. Acid protease production by solid-state fermentation using Aspergillus oryzae MTCC 5341: Optimization of process parameters. J. Ind. Microbiol. Biotechnol. 2010, 37, 129–138. [Google Scholar] [CrossRef]
- Meitner, P.A.; Kassell, B. Bovine pepsinogens and pepsins. A series of zymogens and enzymes that differ in organic phosphate content. Biochem. J. 1971, 121, 249–256. [Google Scholar] [CrossRef] [Green Version]
- Stoops, J.K.; Horgan, D.J.; Runnegar, M.T.C.; De Jersey, J.; Webb, E.C.; Zerner, B. Carboxylesterases (EC 3.1.1). Kinetic studies on carboxylesterases. Biochemistry 1969, 8, 2026–2033. [Google Scholar] [CrossRef]
- Kumar, D.; Kumar, L.; Nagar, S.; Raina, C.; Parshad, R.; Gupta, V.K. Screening, isolation and production of lipase/esterase producing Bacillus sp. strain DVL2 and its potential evaluation in esterification and resolution reactions. Arch. Appl. Sci. Res. 2012, 4, 1763–1770. [Google Scholar]
- Rigling, D.; Heiniger, U.; Hohl, H.R. Reduction of Laccase Activity in dsRNA-Containing Hypovirulent Strains of Cryphonectria (Endothia) Parasitica. Phytopathology 1989, 79, 219–2231. [Google Scholar] [CrossRef]
- Rigling, D.; Van Alfen, N.K. Extra- and Intracellular Laccases of the Chestnut Blight Fungus, Cryphonectria parasitica. Appl. Environ. Microbiol. 1993, 59, 3634–3639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Whitaker, J.R. Protease of Endothia parasitica. In Proteolytic Enzymes; Methods in Enzymology; Academic Press: New York, NY, USA, 1970; Volume 19, pp. 436–445. [Google Scholar]
- Gao, S.; Shain, L. Characterization of an endopolygalacturonase produced by the chestnut blight fungus. Physiol. Mol. Plant Pathol. 1994, 45, 169–179. [Google Scholar] [CrossRef]
- Wu, B.; Gaskell, J.; Held, B.W.; Toapanta, C.; Vuong, T.V.; Ahrendt, S.; Lipzen, A.; Zhang, J.; Schilling, J.S.; Master, E.; et al. Retracted and Republished from: “Substrate-Specific Differential Gene Expression and RNA Editing in the Brown Rot Fungus Fomitopsis pinicola”. Appl. Environ. Microbiol. 2021, 87, e00329-21. [Google Scholar] [CrossRef]
- Shang, J.; Wu, X.; Lan, X.; Fan, Y.; Dong, H.; Deng, Y.; Nuss, D.L.; Chen, B. Large-scale expressed sequence tag analysis for the chestnut blight fungus Cryphonectria parasitica. Fungal Genet. Biol. 2008, 45, 319–327. [Google Scholar] [CrossRef]
- Parker, M.L.; Ng, A.; Waldron, K.W. The phenolic acid and polysaccharide composition of cell walls of bran layers of mature wheat (Triticum aestivum L. cv. Avalon) grains. J. Sci. Food Agric. 2005, 85, 2539–2547. [Google Scholar] [CrossRef]
- Cantero, D.A.; Martínez, C.; Bermejo, M.D.; Cocero, M.J. Simultaneous and selective recovery of cellulose and hemicellulose fractions from wheat bran by supercritical water hydrolysis. Green Chem. 2015, 17, 610–618. [Google Scholar] [CrossRef]
- Pelizer, L.H.; de Moraes, I.O. Influence of water activity on chemical reactivity. Water Act. Theory Appl. Food 2017, 2, 27–54. [Google Scholar] [CrossRef]
- Abbas, A.; Ansumali, S. Global Potential of Rice Husk as a Renewable Feedstock for Ethanol Biofuel Production. BioEnergy Res. 2010, 3, 328–334. [Google Scholar] [CrossRef]
- Ballesteros, L.F.; Teixeira, J.A.; Mussatto, S.I. Chemical, Functional, and Structural Properties of Spent Coffee Grounds and Coffee Silverskin. Food Bioprocess Technol. 2014, 7, 3493–3503. [Google Scholar] [CrossRef] [Green Version]
- Chutmanop, J.; Chuichulcherm, S.; Chisti, Y.; Srinophakun, P. Protease production by Aspergillus oryzae in solid-state fermentation using agroindustrial substrates. J. Chem. Technol. Biotechnol. 2008, 83, 1012–1018. [Google Scholar] [CrossRef]
- Rodríguez Couto, S. Exploitation of biological wastes for the production of value-added products under solid-state fermentation conditions. Biotechnol. J. 2008, 3, 859–870. [Google Scholar] [CrossRef]
- Macchione, M.M.; Merheb, C.W.; Gomes, E.; da Silva, R. Protease Production by Different Thermophilic Fungi. Appl. Biochem. Biotechnol. 2008, 146, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Basheer, S.M.; Chellappan, S.; Beena, P.S.; Sukumaran, R.K.; Elyas, K.K.; Chandrasekaran, M. Lipase from marine Aspergillus awamori BTMFW032: Production, partial purification and application in oil effluent treatment. N. Biotechnol. 2011, 28, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Thakur, S.; Gupte, A. Optimization and hyper production of laccase from novel agaricomycete Pseudolagarobasidium acaciicola AGST3 and its application in in vitro decolorization of dyes. Ann. Microbiol. 2015, 65, 185–196. [Google Scholar] [CrossRef]
- Pitol, L.O.; Finkler, A.T.J.; Dias, G.S.; Machado, A.S.; Zanin, G.M.; Mitchell, D.A.; Krieger, N. Optimization studies to develop a low-cost medium for production of the lipases of Rhizopus microsporus by solid-state fermentation and scale-up of the process to a pilot packed-bed bioreactor. Process. Biochem. 2017, 62, 37–47. [Google Scholar] [CrossRef]
- Wang, J.; Shi, L.; He, X.; Lu, L.; Li, X.; Chen, B. Comparative Secretome Analysis Reveals Perturbation of Host Secretion Pathways by a Hypovirus. Sci. Rep. 2016, 6, 34308. [Google Scholar] [CrossRef] [Green Version]
- Van Alfen, N.K.; Jaynes, R.A.; Anagnostakis, S.L.; Day, P.R. Chestnut Blight: Biological Control by Transmissible Hypovirulence in Endothia parasitica. Science 1975, 12, 890–891. [Google Scholar] [CrossRef]
- Shapira, R.; Choi, G.H.; Nuss, D.L. Virus-like genetic organization and expression strategy for a double-stranded RNA genetic element associated with biological control of chestnut blight. EMBO J. 1991, 10, 731–739. [Google Scholar] [CrossRef]
- Allen, T.D.; Dawe, A.L.; Nuss, D.L. Use of cDNA Microarrays To Monitor Transcriptional Responses of the Chestnut Blight Fungus Cryphonectria parasitica to Infection by Virulence-Attenuating Hypoviruses. Eukaryot. Cell 2003, 2, 1253–1265. [Google Scholar] [CrossRef] [Green Version]
- Ikasari, L.; Mitchell, D.A. Protease production by Rhizopus oligosporus in solid-state fermentation. World J. Microbiol. Biotechnol. 1994, 10, 320–324. [Google Scholar] [CrossRef]
- Rivera-Hoyos, C.; Morales-Álvarez, E.; Poutou-Piñales, R.; Pedroza-Rodríguez, A.; Vazquez, R.R.; Delgado-Boada, J. Fungal Laccases. Fungal Biol. Rev. 2013, 27, 67–82. [Google Scholar] [CrossRef]
- Majeau, J.-A.; Brar, S.K.; Tyagi, R.D. Laccases for removal of recalcitrant and emerging pollutants. Bioresour. Technol. 2010, 101, 2331–2350. [Google Scholar] [CrossRef] [PubMed]
- Masutti, D.; Borgognone, A.; Setti, L. Production of Enzymes from Rice Husks and Wheat Straw in Solid State Fermentation. Chem. Eng. Trans. 2012, 27, 133–138. [Google Scholar] [CrossRef]
- Parsley, T.B.; Chen, B.; Geletka, L.M.; Nuss, D.L. Differential Modulation of Cellular Signaling Pathways by Mild and Severe Hypovirus Strains. Eukaryot. Cell 2002, 1, 401–413. [Google Scholar] [CrossRef] [Green Version]
- Geisseler, D.; Horwath, W.R. Regulation of extracellular protease activity in soil in response to different sources and concentrations of nitrogen and carbon. Soil Biol. Biochem. 2008, 40, 3040–3048. [Google Scholar] [CrossRef]
- Tzanov, T.; Basto, C.; Gübitz, G.M.; Cavaco-Paulo, A. Laccases to Improve the Whiteness in a Conventional Bleaching of Cotton. Macromol. Mater. Eng. 2003, 288, 807–810. [Google Scholar] [CrossRef] [Green Version]
- Naganthran, A.; Masomian, M.; Rahman, R.N.Z.R.A.; Ali, M.S.M.; Nooh, H.M. Improving the Efficiency of New Automatic Dishwashing Detergent Formulation by Addition of Thermostable Lipase, Protease and Amylase. Molecules 2017, 22, 1577. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Origin | Canker Type | CHV1 |
|---|---|---|---|
| CpA | Tuscany | evolutive | − |
| CpB2 | Tuscany | evolutive | − |
| CpC4 | Lombardy | superficial | + |
| CpC7 | Lombardy | evolutive | − |
| Substrate for SSF | |||
|---|---|---|---|
| Chemical Components | Wheat Bran (ZW *) | Rice Husk (RH) | Spent Coffee Grounds (CG) |
| Cellulose | 32 | 33.4 | 12.5 |
| Hemicellulose | 21 | 21 | 39 |
| Lignin | 22 | 18.3 | 24 |
| Carbohydrates | 26.6 | 1.1 | 30 |
| Protein | 14.1 | 2 | 17.5 |
| Lipids | 5.5 | 0.8 | 2.3 |
| Ash | 0.5 | 15.5 | 1.3 |
| Agro-Food Waste | Abbreviation | Ratio Water/Waste | Milled a | C. parasitica Growth b |
|---|---|---|---|---|
| Organic wheat bran | OWA | 1:1 | − | ++ |
| OWB | 1.5:1 | − | ++ | |
| Zootechnical wheat bran | ZW | 1:1 | − | ++ |
| Rice husk | RHM | 1:1 | + | + |
| RHU | 1:1 | − | −/+ | |
| Spent coffee grounds | CGA | 2:1 | − | −/+ |
| CGB | 1:1 | − | −/+ |
| Enzyme | Microorganism | SSF Substrate | Fermentation Period | Enzyme Substate | Unit Definition | Enzyme Activity | Reference |
|---|---|---|---|---|---|---|---|
| Protease | A. oryzae MTCC 5341 | Wheat bran with supplements | 120 h | Hemoglobin | 1 U = 0.001 increase in A280 nm/min | 864,000 U/gds a | Vishwanatha et al., 2010 |
| C. parasitica CpA | Wheat bran | 9 days | Hemoglobin | 1 AU = 0.01 increase in A280 nm/min | 3955.6 U/gsf b | Current study | |
| A. oryzae (Ozykat-1) | Wheat bran and rice bran | 4 days | Casein | 1 U = release 1 μg tyrosine/min | 1200 U/gds | Chutmanop et al., 2008 | |
| Laccase | P. acaciicola AGST3 | Wheat bran with supplements | 12 days | ABTS * | 1 U = oxidation 1 μmol ABTS/min | 535,000 U/gds | Thakur and Gupte 2015 |
| T. versicolor JSRK13 | Parthenium sp. with supplements | 17 days | Guaiacol | 1 U = 0.01 increase A470 nm/min | 185 U/gsf | Singh et al., 2019 | |
| C. parasitica CpC4 | Wheat bran | 15 days | 2,6-Dimethoxyphenol | 1 AU = 1.0 increase A468 nm/min | 93.8 U/gsf | Current study | |
| Esterase /Lipase | R. microsporus CPQBA 312-07 DRM | Wheat bran and sugarcane bagasse with supplements | 18 h | Olive oil | 1 U = release 1 μmol fatty acid/min | 265 U/gsf | Pitol et al., 2017 |
| A. ibericus MUM 03.49 | Olive pomace and wheat bran with supplements | 7 days | pNP-butyrate ** | 1 U = release 1 μmol di pNP/min | 223 U/gds | Oliveira et al., 2017 | |
| C. parasitica CpC4 | Wheat bran | 15 days | pNP-butyrate | 1 U = release 1 μmol di pNP/min | 2.55 U/gsf | Current study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Savino, S.; Bulgari, D.; Monti, E.; Gobbi, E. Agro-Industrial Wastes: A Substrate for Multi-Enzymes Production by Cryphonectria parasitica. Fermentation 2021, 7, 279. https://doi.org/10.3390/fermentation7040279
Savino S, Bulgari D, Monti E, Gobbi E. Agro-Industrial Wastes: A Substrate for Multi-Enzymes Production by Cryphonectria parasitica. Fermentation. 2021; 7(4):279. https://doi.org/10.3390/fermentation7040279
Chicago/Turabian StyleSavino, Salvatore, Daniela Bulgari, Eugenio Monti, and Emanuela Gobbi. 2021. "Agro-Industrial Wastes: A Substrate for Multi-Enzymes Production by Cryphonectria parasitica" Fermentation 7, no. 4: 279. https://doi.org/10.3390/fermentation7040279
APA StyleSavino, S., Bulgari, D., Monti, E., & Gobbi, E. (2021). Agro-Industrial Wastes: A Substrate for Multi-Enzymes Production by Cryphonectria parasitica. Fermentation, 7(4), 279. https://doi.org/10.3390/fermentation7040279