

Integration of Dark Fermentation with Microbial Electrolysis Cells for Biohydrogen and Methane Production from Distillery Wastewater and Glycerol Waste Co-Digestion

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Substrates and Inoculum
2.2. Optimization of DW and GW Proportions in Dark Fermentative Hydrogen Production
2.3. Continuous Biohydrogen Production from DW and GW Co-Digestion in a CSTR
2.4. Methane Production from Dark Fermentation Effluent Using a Single-Chamber MEC

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Dark Fermentation Effluent |
|---|---|
| pH | 5.1 |
| TS (g/L) | 56.6 ± 3.2 |
| VS (g/L) | 31.7 ± 3.1 |
| Ash (g/L) | 24.9 ± 0.2 |
| Alkalinity (g/L) | 5.8 ± 0.1 |
| Total volatile fatty acid (g/L) | 5.0 ± 3.0 |
| COD (g/L) | 78.1 ± 7.3 |
2.5. Microbial Community Analysis Using 16S rRNA Amplicon Sequencing on an Illumination Platform
2.6. Analytical Methods
3. Results and Discussion
3.1. Biohydrogen Production from a Dark Fermentative of DW and GW Co-Digestion at Various Ratios
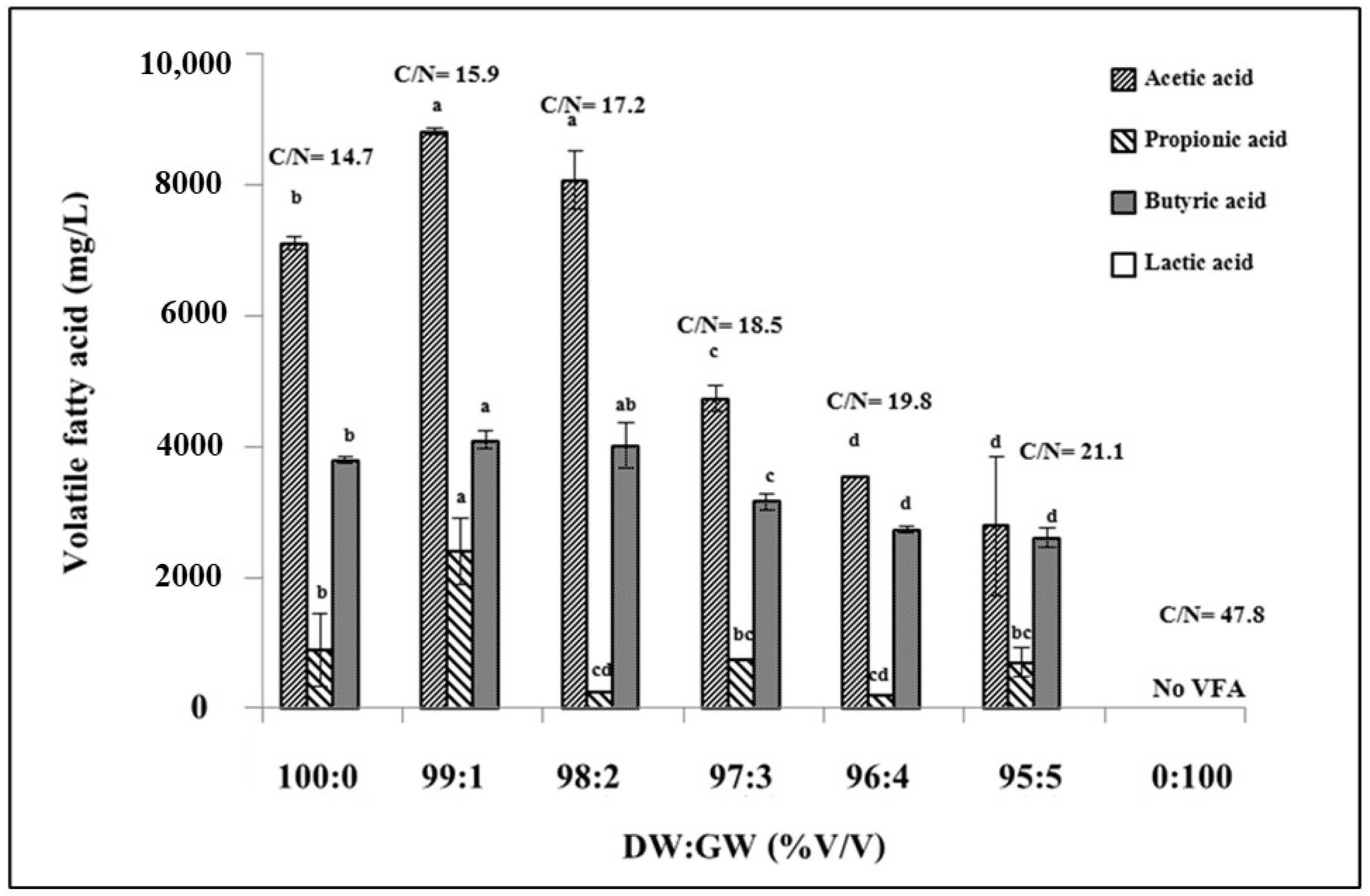
3.2. VFA Production from a Dark Fermentative of DW and GW Co-Digestion at Various Ratios
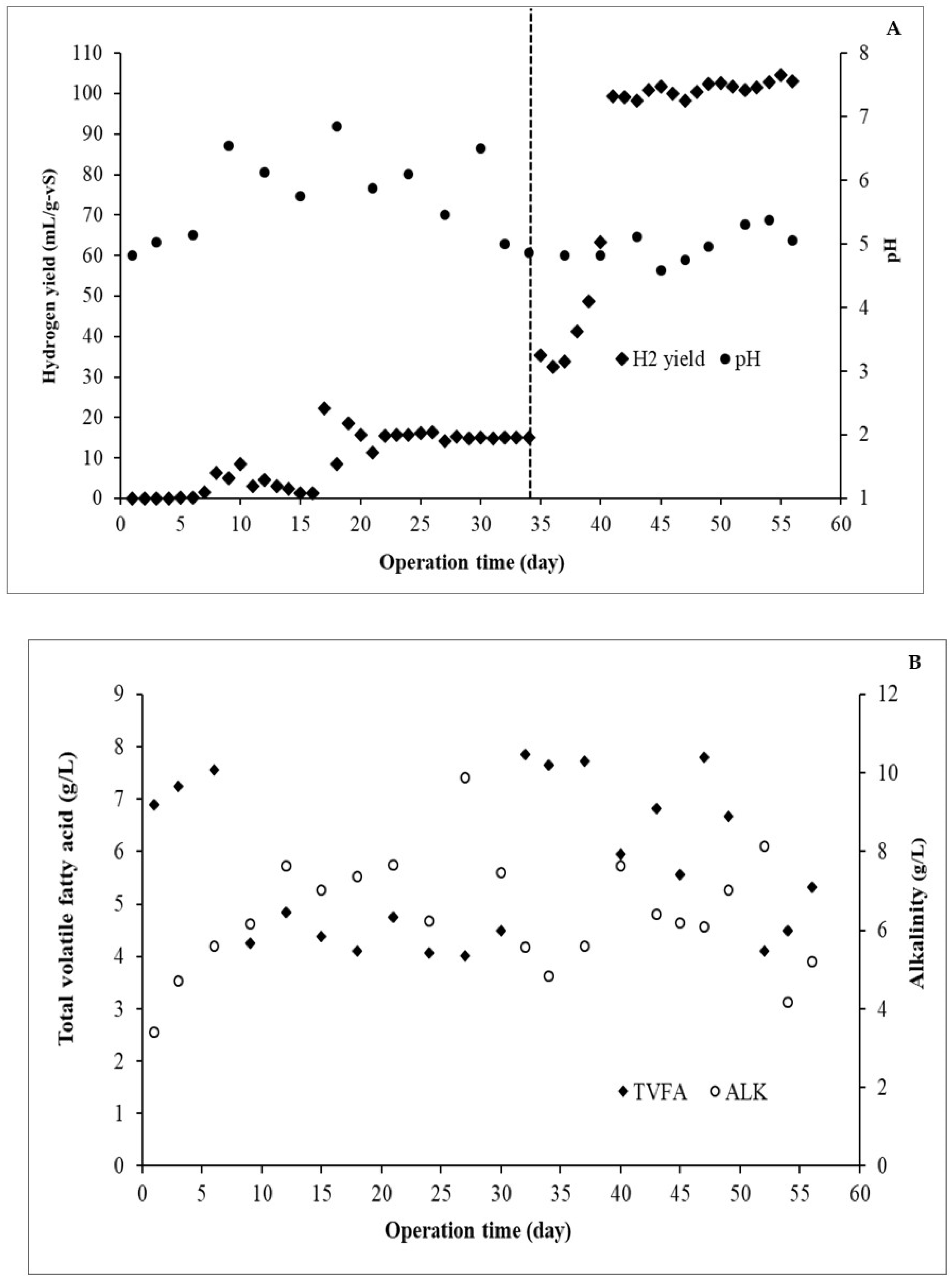
3.3. Continuous Biohydrogen Production from DW and GW Co-Digestion in a CSTR
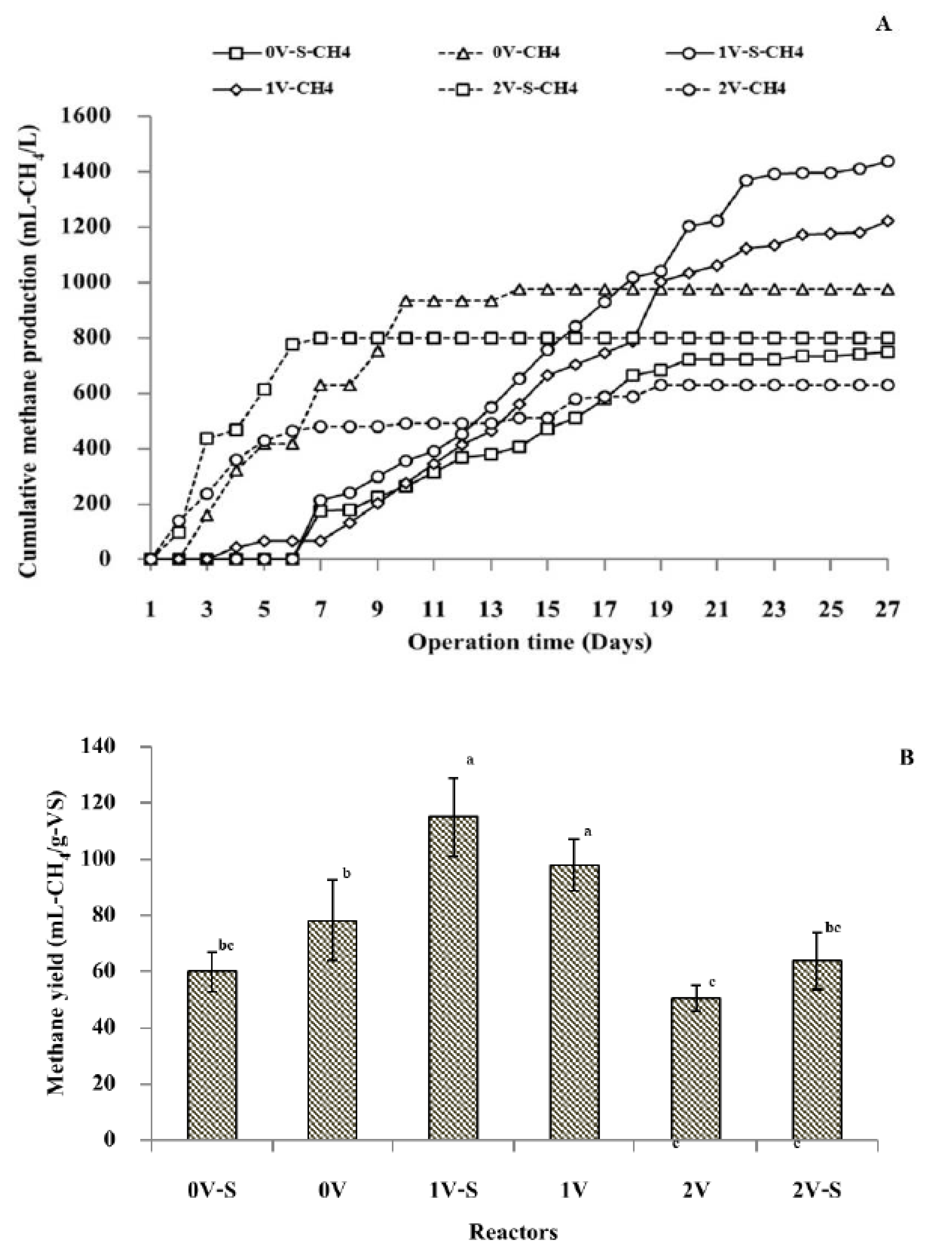
3.4. Methane Production from the Dark Fermentation Effluent Using a Single-Chamber MEC
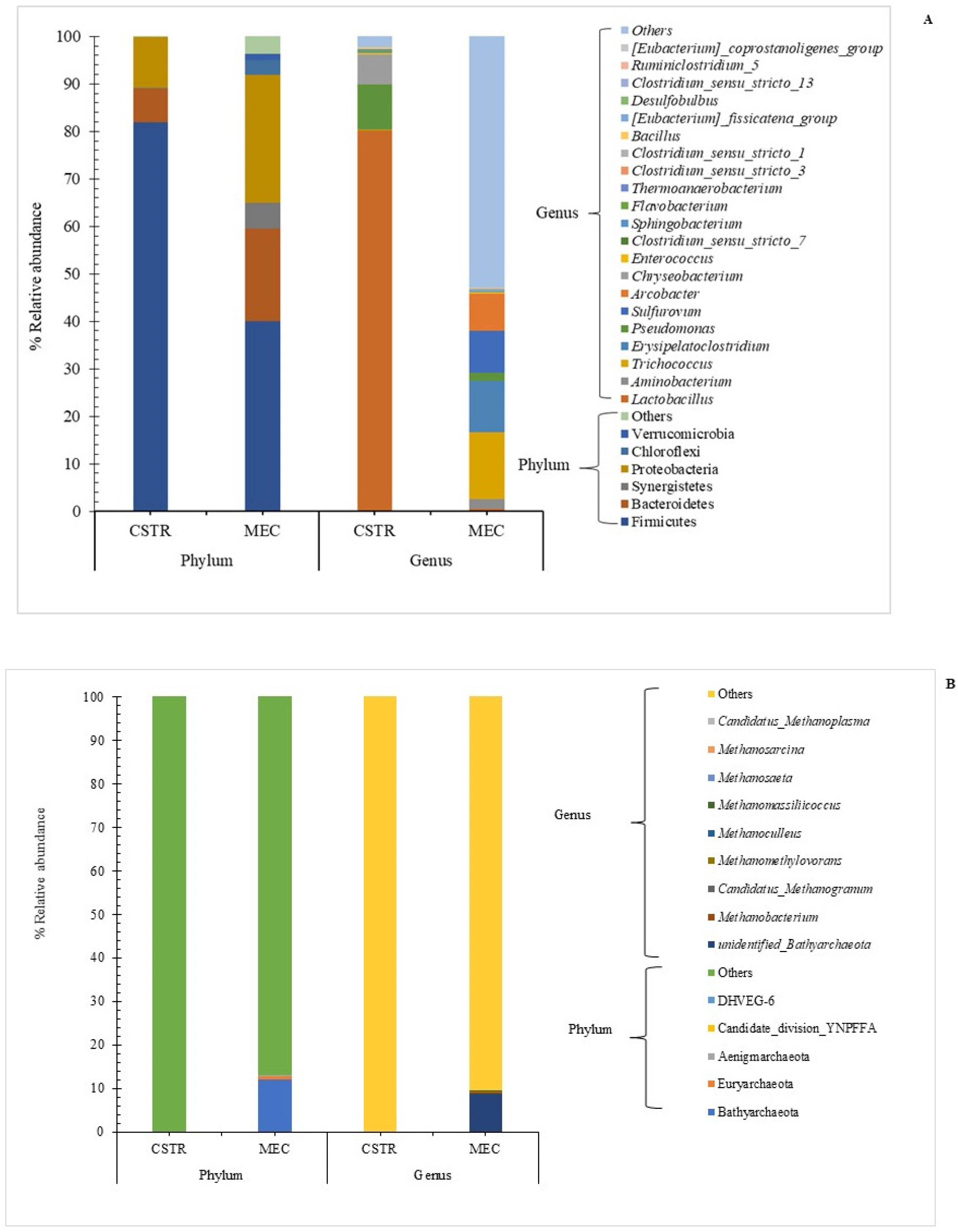
3.5. Microbial Community Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Babadi, A.A.; Rahmati, S.; Fakhlaei, R.; Barati, B.; Wang, S.; Doherty, W.; Ostrikov, K. Emerging technologies for biodiesel production: Processes, challenges, and opportunities. Biomass Bioenergy 2022, 163, 106521. [Google Scholar] [CrossRef]
- Sittijunda, S.; Reungsang, A. Valorization of crude glycerol into hydrogen, 1,3-propanediol, and ethanol in an up-flow anaerobic sludge blanket (UASB) reactor under thermophilic conditions. Renew. Energy 2020, 161, 361–372. [Google Scholar] [CrossRef]
- Decarpigny, C.; Aljawish, A.; His, C.; Fertin, B.; Bigan, M.; Dhulster, P.; Millares, M.; Froidevaux, R. Bioprocesses for the Biodiesel Production from Waste Oils and Valorization of Glycerol. Energies 2022, 15, 3381. [Google Scholar] [CrossRef]
- Luo, X.; Ge, X.; Cui, S.; Li, Y. Value-added processing of crude glycerol into chemicals and polymers. Bioresour. Technol. 2016, 215, 144–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kongjan, P.; Jariyaboon, R.; Reungsang, A.; Sittijunda, S. Co-fermentation of 1,3-propanediol and 2,3-butanediol from crude glycerol derived from the biodiesel production process by newly isolated Enterobacter sp.: Optimization factors affecting. Bioresour. Technol. Rep. 2021, 13, 100616. [Google Scholar] [CrossRef]
- Akhbari, A.; Ibrahim, S. Bioenergy recovery from food waste through dark fermentation direction. Res. Sq. 2022. preprint. [Google Scholar]
- Policastro, G.; Cesaro, A.; Fabbricino, M. Photo-fermentative hydrogen production from cheese whey: Engineering of a mixed culture process in a semi-continuous, tubular photo-bioreactor. Int. J. Hydrogen Energy 2022, in press. [Google Scholar] [CrossRef]
- Sun, M.; Sheng, G.-P.; Zhang, L.; Xia, C.-R.; Mu, Z.-X.; Liu, X.-W.; Wang, H.-L.; Yu, H.-Q.; Qi, R.; Yu, T.; et al. An MEC-MFC-Coupled System for Biohydrogen Production from Acetate. Environ. Sci. Technol. 2008, 42, 8095–8100. [Google Scholar] [CrossRef] [PubMed]
- Amrut Pawar, A.; Karthic, A.; Lee, S.; Pandit, S.; Jung, S.P. Microbial electrolysis cells for electromethanogenesis: Materials, configurations and operations. Environ. Eng. Res. 2022, 27, 200484. [Google Scholar] [CrossRef]
- Nelabhotla, A.B.T.; Dinamarca, C. Bioelectrochemical CO2 Reduction to Methane: MES Integration in Biogas Production Processes. Appl. Sci. 2019, 9, 1056. [Google Scholar] [CrossRef] [Green Version]
- Cheng, D.; Ngo, H.H.; Guo, W.; Chang, S.W.; Nguyen, D.D.; Deng, L.; Chen, Z.; Ye, Y.; Bui, X.T.; Hoang, N.B. Advanced strategies for enhancing dark fermentative biohydrogen production from biowaste towards sustainable environment. Bioresour. Technol. 2022, 351, 127045. [Google Scholar] [CrossRef]
- Badia-Fabregat, M.; Rago, L.; Baeza, J.A.; Guisasola, A. Hydrogen production from crude glycerol in an alkaline microbial electrolysis cell. Int. J. Hydrogen Energy 2019, 44, 17204–17213. [Google Scholar] [CrossRef]
- Sarma, S.J.; Brar, S.K.; Sydney, E.B.; Le Bihan, Y.; Buelna, G.; Soccol, C.R. Microbial hydrogen production by bioconversion of crude glycerol: A review. Int. J. Hydrogen Energy 2012, 37, 6473–6490. [Google Scholar] [CrossRef]
- Nguyen, P.K.T.; Das, G.; Kim, J.; Yoon, H.H. Hydrogen production from macroalgae by simultaneous dark fermentation and microbial electrolysis cell. Bioresour. Technol. 2020, 315, 123795. [Google Scholar] [CrossRef]
- Thu Ha Tran, T.; Khanh Thinh Nguyen, P. Enhanced hydrogen production from water hyacinth by a combination of ultrasonic-assisted alkaline pretreatment, dark fermentation, and microbial electrolysis cell. Bioresour. Technol. 2022, 357, 127340. [Google Scholar] [CrossRef] [PubMed]
- Im, S.; Yun, Y.-M.; Song, Y.-C.; Kim, D.-H. Enhanced anaerobic digestion of glycerol by promoting DIET reaction. Biochem. Eng. J. 2019, 142, 18–26. [Google Scholar] [CrossRef]
- Zeppilli, M.; Pavesi, D.; Gottardo, M.; Micolucci, F.; Villano, M.; Majone, M. Using effluents from two-phase anaerobic digestion to feed a methane-producing microbial electrolysis. Chem. Eng. J. 2017, 328, 428–433. [Google Scholar] [CrossRef]
- Prasertsan, P.; Leamdum, C.; Chantong, S.; Mamimin, C.; Kongjan, P.; O-Thong, S. Enhanced biogas production by co-digestion of crude glycerol and ethanol with palm oil mill effluent and microbial community analysis. Biomass Bioenergy 2021, 148, 106037. [Google Scholar] [CrossRef]
- Sittijunda, S.; Reungsang, A. Methane Production from the Co-digestion of Algal Biomass with Crude Glycerol by Anaerobic Mixed Cultures. Waste Biomass Valorization 2020, 11, 1873–1881. [Google Scholar] [CrossRef]
- Wongarmat, W.; Sittijunda, S.; Mamimin, C.; Reungsang, A. Acidogenic phase anaerobic digestion of pretreated sugarcane filter cake for co-digestion with biogas effluent to enhance the methane production. Fuel 2022, 310, 122466. [Google Scholar] [CrossRef]
- Hoarau, J.; Grondin, I.; Caro, Y.; Petit, T. Sugarcane Distillery Spent Wash, a New Resource for Third-Generation Biodiesel Production. Water 2018, 10, 1623. [Google Scholar] [CrossRef]
- Kongjan, P.; O-Thong, S.; Angelidaki, I. Biohydrogen production from desugared molasses (DM) using thermophilic mixed cultures immobilized on heat treated anaerobic sludge granules. Int. J. Hydrogen Energy 2011, 36, 14261–14269. [Google Scholar] [CrossRef]
- Thong, S.; Prasertsan, P.; Birkeland, N.K. Evaluation of methods for preparing hydrogen-producing seed inocula under thermophilic condition by process performance and microbial community analysis. Bioresour. Technol. 2009, 100, 909–918. [Google Scholar] [CrossRef]
- Angelidaki, I.; Sanders, W. Assessment of the anaerobic biodegradability of macropollutants. Re Views Environ. Sci. Bio Technol. 2004, 3, 117–129. [Google Scholar] [CrossRef]
- Zhou, J.; Bruns, M.A.; Tiedje, J.M. DNA recovery from soils of diverse composition. Appl. Environ. Microbiol. 1996, 62, 316–322. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Clesceri, L.S. Standard Methods for Examination of Water and Wastewater; American Public Health Association: Washington, DC, USA, 1998; Volume 9. [Google Scholar]
- Official Methods of Analysis of AOAC International, 19th ed.; AOAC International: Gaithersburg, MD, USA, 2012.
- Sani, K.; Jariyaboon, R.; O-Thong, S.; Cheirsilp, B.; Kaparaju, P.; Raketh, M.; Kongjan, P. Deploying two-stage anaerobic process to co-digest greasy sludge and waste activated sludge for effective waste treatment and biogas recovery. J. Environ. Manag. 2022, 316, 115307. [Google Scholar] [CrossRef] [PubMed]
- Mata-Alvarez, J.; Dosta, J.; Romero-Güiza, M.S.; Fonoll, X.; Peces, M.; Astals, S. A critical review on anaerobic co-digestion achievements between 2010 and 2013. Renew. Sustain. Energy Rev. 2014, 36, 412–427. [Google Scholar] [CrossRef]
- Hartmann, H.; Angelidaki, I.; Ahring, B.K. Co-digestion of the organic fraction of municipal waste with other waste types. In Biomethanization of the Organic Fraction of Municipal Solid Wastes; IWA Publishing: London UK, 2002; pp. 181–200. [Google Scholar]
- Venkataramanan, K.P.; Boatman, J.J.; Kurniawan, Y.; Taconi, K.A.; Bothun, G.D.; Scholz, C. Impact of impurities in biodiesel-derived crude glycerol on the fermentation by Clostridium pasteurianum ATCC 6013. Appl. Microbiol. Biotechnol. 2012, 93, 1325–1335. [Google Scholar] [CrossRef]
- Sittijunda, S.; Reungsang, A. Media optimization for biohydrogen production from waste glycerol by anaerobic thermophilic mixed cultures. Int. J. Hydrogen Energy 2012, 37, 15473–15482. [Google Scholar] [CrossRef]
- Hamawand, I.; Baillie, C. Anaerobic Digestion and Biogas Potential: Simulation of Lab and Industrial-Scale Processes. Energies 2015, 8, 454–474. [Google Scholar] [CrossRef]
- Issah, A.A.; Kabera, T. Impact of volatile fatty acids to alkalinity ratio and volatile solids on biogas production under thermophilic conditions. Waste Manag. Res. J. Int. Solid Wastes Public Clean. Assoc. ISWA 2021, 39, 871–878. [Google Scholar] [CrossRef] [PubMed]
- Ding, A.; Yang, Y.; Sun, G.; Wu, D. Impact of applied voltage on methane generation and microbial activities in an anaerobic microbial electrolysis cell (MEC). Chem. Eng. J. 2016, 283, 260–265. [Google Scholar] [CrossRef]
- Jayabalan, T.; Matheswaran, M.; Naina Mohammed, S. Biohydrogen production from sugar industry effluents using nickel based electrode materials in microbial electrolysis cell. Int. J. Hydrogen Energy 2019, 44, 17381–17388. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, R.; Chen, C.; Liu, G.; He, Y.; Liu, X. Biogas production from co-digestion of corn stover and chicken manure under anaerobic wet, hemi-solid, and solid state conditions. Bioresour. Technol. 2013, 149, 406–412. [Google Scholar] [CrossRef] [PubMed]
- Świątek, M.; Lewicki, A.; Szymanowska, D.; Kubiak, P. The effect of introduction of chicken manure on the biodiversity and performance of an anaerobic digester. Electron. J. Biotechnol. 2019, 37, 25–33. [Google Scholar] [CrossRef]
- Wang, H.; Zhu, X.; Yan, Q.; Zhang, Y.; Angelidaki, I. Microbial community response to ammonia levels in hydrogen assisted biogas production and upgrading process. Bioresour. Technol. 2020, 296, 122276. [Google Scholar] [CrossRef]
- Vanwonterghem, I.; Evans, P.N.; Parks, D.H.; Jensen, P.D.; Woodcroft, B.J.; Hugenholtz, P.; Tyson, G.W. Methylotrophic methanogenesis discovered in the archaeal phylum Verstraetearchaeota. Nat. Microbiol. 2016, 1, 16170. [Google Scholar] [CrossRef] [Green Version]
- Florou-Paneri, P.; Christaki, E.; Bonos, E. Lactic acid bacteria as source of functional ingredients. In Lactic Acid Bacteria—R & D for Food, Health and Livestock Purposes; Kongo, M., Ed.; Intech Open: Rijeka, Croatia, 2012; pp. 589–614. [Google Scholar]
- Tsapekos, P.; Kougias, P.G.; Treu, L.; Campanaro, S.; Angelidaki, I. Process performance and comparative metagenomic analysis during co-digestion of manure and lignocellulosic biomass for biogas production. Appl. Energy 2017, 185, 126–135. [Google Scholar] [CrossRef]
- Zhang, J.; Zhao, W.; Yang, J.; Li, Z.; Zhang, J.; Zang, L.J.B.T. Comparison of mesophilic and thermophilic dark fermentation with nickel ferrite nanoparticles supplementation for biohydrogen production. Bioresour. Technol. 2021, 329, 124853. [Google Scholar] [CrossRef]
- Silva, V.; Ratti, R.; Sakamoto, I.; Andrade, M.; Varesche, M.J.R.E. Biotechnological products in batch reactors obtained from cellulose, glucose and xylose using thermophilic anaerobic consortium. Renew. Energy 2018, 125, 537–545. [Google Scholar] [CrossRef]
- Strazzera, G.; Battista, F.; Tonanzi, B.; Rossetti, S.; Bolzonella, D. Optimization of short chain volatile fatty acids production from household food waste for biorefinery applications. Environ. Technol. Innov. 2021, 23, 101562. [Google Scholar] [CrossRef]
- Enzmann, F.; Mayer, F.; Rother, M.; Holtmann, D. Methanogens: Biochemical background and biotechnological applications. AMB Express 2018, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Pope, P.B.; Eijsink, V.G.; Schnürer, A. Characterization of microbial community structure during continuous anaerobic digestion of straw and cow manure. Microb. Biotechnol. 2015, 8, 815–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lomans, B.P.; Maas, R.; Luderer, R.; Op den Camp, H.J.; Pol, A.; van der Drift, C.; Vogels, G.D. Isolation and characterization of Methanomethylovorans hollandica gen. nov., sp. nov., isolated from freshwater sediment, a methylotrophic methanogen able to grow on dimethyl sulfide and methanethiol. Appl. Environ. Microbiol. 1999, 65, 3641–3650. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Wang, L.; Gao, L.; Wang, A.-J. Chapter 5.10—Hydrogen and Methane Production in Bioelectrochemical System: Biocathode Structure and Material Upgrading. In Microbial Electrochemical Technology; Mohan, S.V., Varjani, S., Pandey, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 921–953. [Google Scholar]





| Parameter | DW | GW |
|---|---|---|
| pH | 4.8 | 10.0 |
| Total solid (TS, g/L) | 220.9 ± 53.0 | 769.1 ± 0.5 |
| Volatile solid (VS, g/L) | 160.6 ± 33.0 | 688.7 ± 0.1 |
| Ash (g/L) | 60.3 ± 49.7 | 80.4 ± 0.6 |
| Chemical oxygen demand (COD, g/L) | 106.8 ± 12.9 | 622.9 ± 93.1 |
| Total Kjeldahl nitrogen (g/L) | 4.3 ± 0.2 | 0.5 ± 0.3 |
| Nitrogen content (%wt) | 0.4 ± 0.0 | ND |
| Carbon content (%wt) | 5.7 ± 0.0 | 47.8 ± 0.0 |
| C/N ratio | 14.7 | 47.8 |
| Hydrogen content (%wt) | 10.2 ± 0.0 | 10.0 ± 0.0 |
| Oxygen content (%wt) | 74.1 ± 0.3 | 39.3 ± 0.3 |
| Parameter | INH | INM |
|---|---|---|
| pH | 6.0 | 7.3 |
| TS (g/L) | 23.6 ± 2.6 | 50.5 ± 2.5 |
| Total dissolved solid (g/L) | 16.0 ± 1.5 | 4.5 ± 1.3 |
| Total suspended solid (g/L) | 7.7 ± 2.9 | 46.0 ± 1.2 |
| VS (g/L) | 14.3 ± 2.1 | 37.8 ± 2.6 |
| Volatile suspended solid (g/L) | 7.7 ± 2.9 | 33.3 ± 1.6 |
| Volatile dissolved solid (g/L) | 6.7 ± 1.0 | 4.5 ± 1.3 |
| Ash (g/L) | 9.3 ± 0.7 | 12.7 ± 0.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sittijunda, S.; Baka, S.; Jariyaboon, R.; Reungsang, A.; Imai, T.; Kongjan, P. Integration of Dark Fermentation with Microbial Electrolysis Cells for Biohydrogen and Methane Production from Distillery Wastewater and Glycerol Waste Co-Digestion. Fermentation 2022, 8, 537. https://doi.org/10.3390/fermentation8100537
Sittijunda S, Baka S, Jariyaboon R, Reungsang A, Imai T, Kongjan P. Integration of Dark Fermentation with Microbial Electrolysis Cells for Biohydrogen and Methane Production from Distillery Wastewater and Glycerol Waste Co-Digestion. Fermentation. 2022; 8(10):537. https://doi.org/10.3390/fermentation8100537
Chicago/Turabian StyleSittijunda, Sureewan, Sulfan Baka, Rattana Jariyaboon, Alissara Reungsang, Tsuyoshi Imai, and Prawit Kongjan. 2022. "Integration of Dark Fermentation with Microbial Electrolysis Cells for Biohydrogen and Methane Production from Distillery Wastewater and Glycerol Waste Co-Digestion" Fermentation 8, no. 10: 537. https://doi.org/10.3390/fermentation8100537
APA StyleSittijunda, S., Baka, S., Jariyaboon, R., Reungsang, A., Imai, T., & Kongjan, P. (2022). Integration of Dark Fermentation with Microbial Electrolysis Cells for Biohydrogen and Methane Production from Distillery Wastewater and Glycerol Waste Co-Digestion. Fermentation, 8(10), 537. https://doi.org/10.3390/fermentation8100537