
Plant Proteins as an Alternative Nitrogen Source for Chiral Purity L-Lactic Acid Fermentation from Lignocellulose Feedstock



Abstract
:1. Introduction
2. Materials and Methods
2.1. Raw Feedstock
2.2. Reagents and Enzymes
2.3. Microorganisms and Medium
2.4. Pretreatment and Biodetoxification
2.5. Preparation of Cottonseed Meal Hydrolysate
2.6. Cellulosic L-Lactic Acid SSCF
2.7. Analysis
3. Results and Discussions
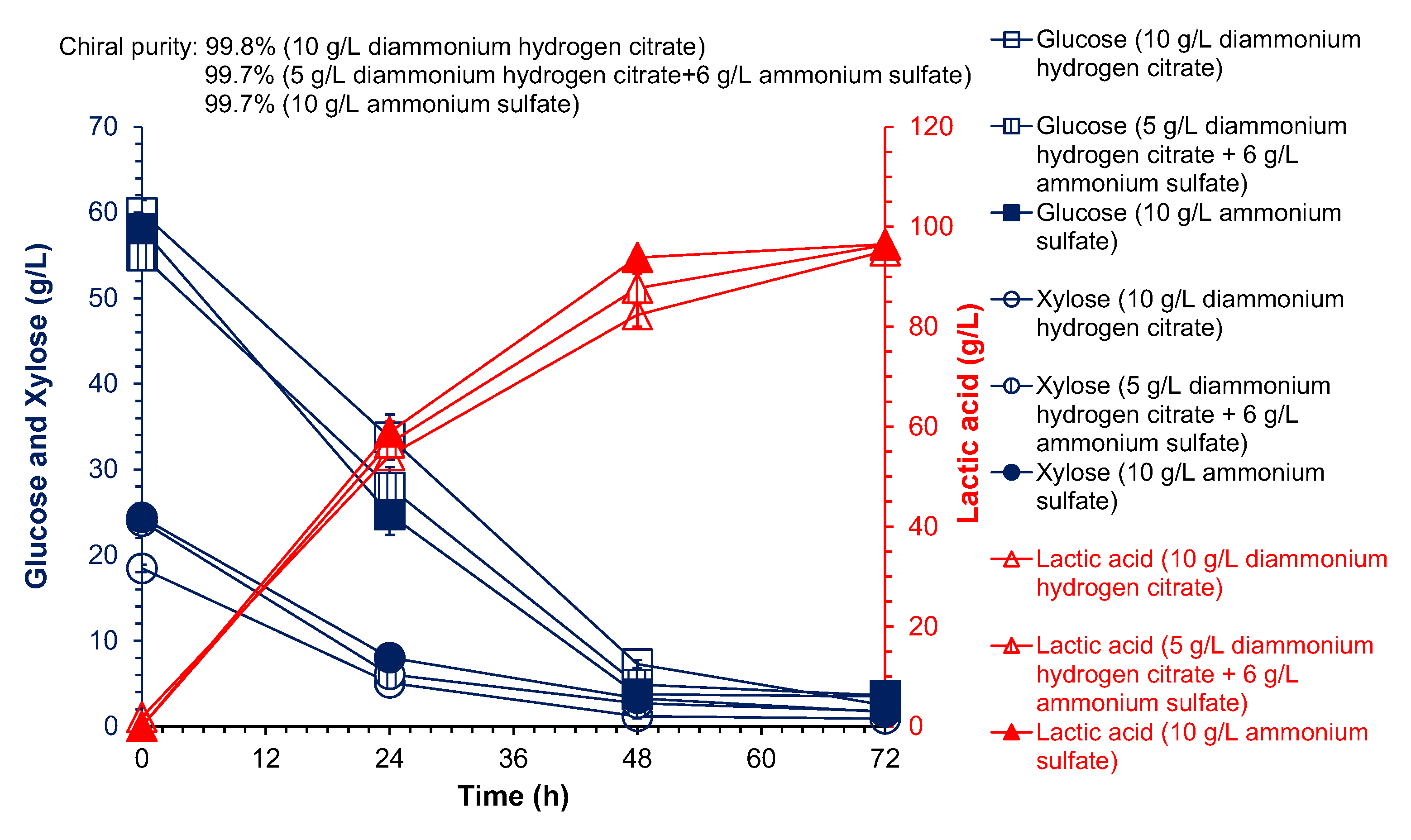
3.1. Screening Alternative Nitrogen Sources for High Chiral Purity Lactic Acid Fermentation
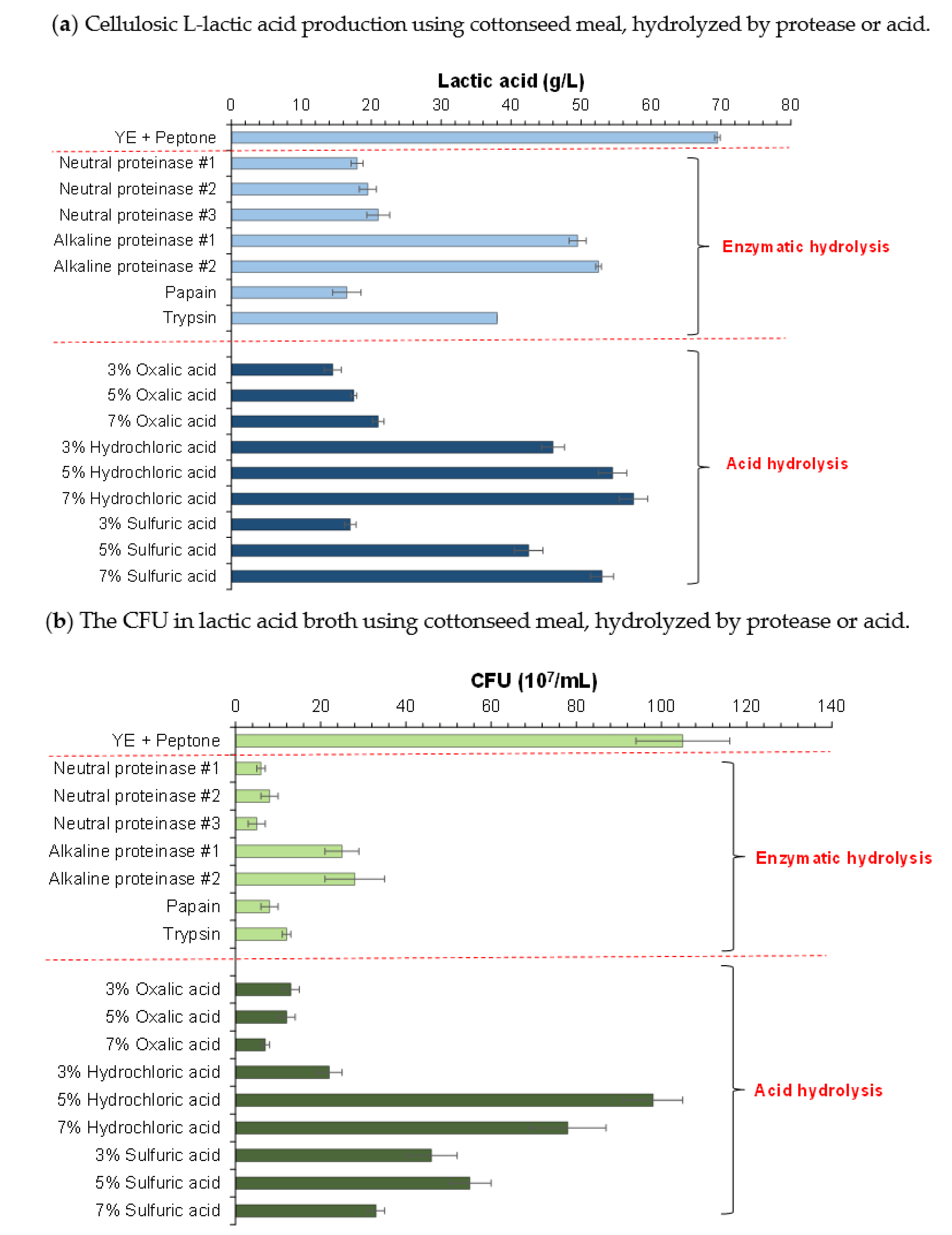
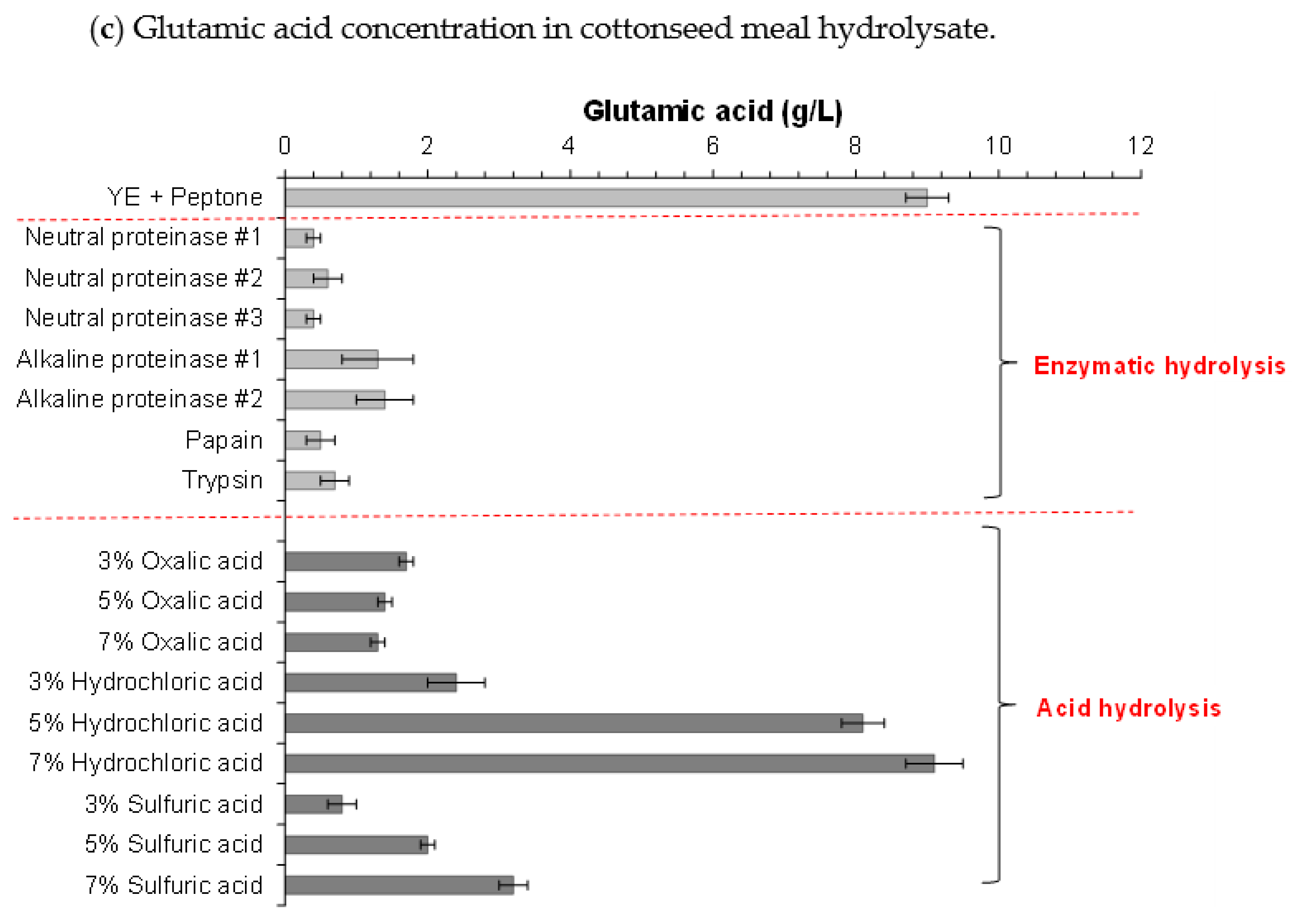
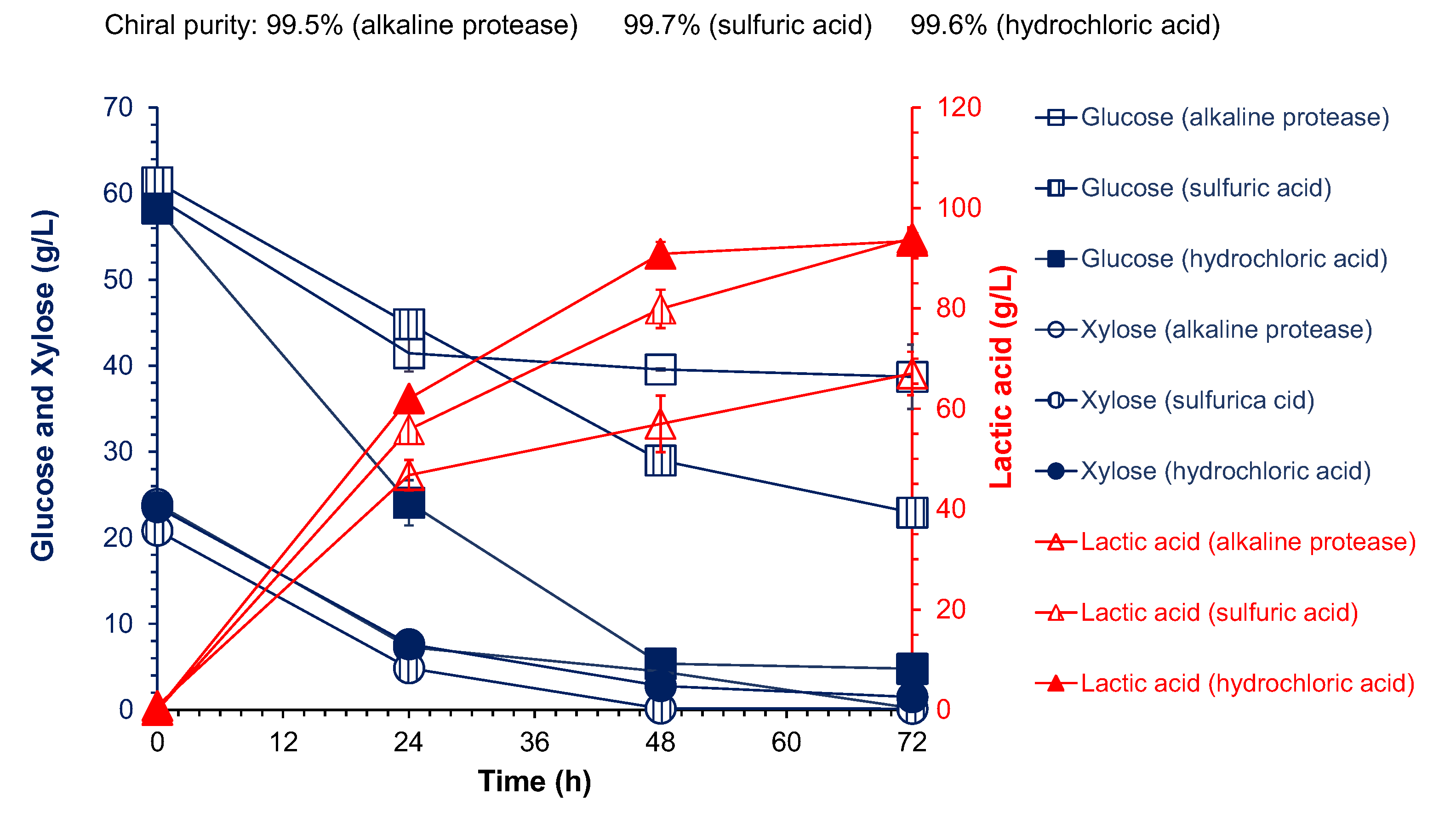
3.2. Hydrolysis of Cottonseed Meal and Consequent Cellulosic L-Lactic Acid Fermentation
3.3. Cellulose L-Lactic Acid Production by SSCF
3.4. Preliminary Techno-Economic Evaluations
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lu, Z.; He, F.; Shi, Y.; Lu, M.; Yu, L. Fermentative production of L(+)-lactic acid using hydrolyzed acorn starch, persimmon juice and wheat bran hydrolysate as nutrients. Bioresour. Technol. 2010, 101, 3642–3648. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Rahman, M.A.; Xiao, Y.; Tashiro, Y.; Wang, Y.; Zendo, T.; Sakai, K.; Sonomoto, K. Fed-batch fermentation for enhanced lactic acid production from glucose/xylose mixture without carbon catabolite repression. J. Biosci. Bioeng. 2015, 119, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Inkinen, S.; Hakkarainen, M.; Albertsson, A.C.; Sodergard, A. From lactic acid to poly(lactic acid) (PLA): Characterization and analysis of PLA and its precursors. Biomacromolecules 2011, 12, 523–532. [Google Scholar] [CrossRef]
- Ahmad, A.; Banat, F.; Taher, H. A review on the lactic acid fermentation from low-cost renewable materials: Recent developments and challenges. Environ. Technol. Innov. 2020, 20, 101138. [Google Scholar] [CrossRef]
- Tejayadi, S.; Cheryan, M. Lactic acid from cheese whey permeate. Productivity and economics of a continuous membrane bioreactor. Appl. Microbio. Biotechnol. 1995, 43, 242–248. [Google Scholar] [CrossRef]
- Altaf, M.; Venkateshwar, M.; Srijana, M.; Reddy, G. An economic approach for L-(+) lactic acid fermentation by Lactobacillus amylophilus GV6 using inexpensive carbon and nitrogen sources. J. Appl. Microbiol. 2007, 103, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Juturu, V.; Wu, J.C. Microbial production of lactic acid: The latest development. Crit. Rev. Biotechnol. 2016, 36, 967–977. [Google Scholar] [CrossRef]
- Zhang, Y.; Yoshida, M.; Vadlani, P.V. Biosynthesis of D-lactic acid from lignocellulosic biomass. Biotechnol. Lett. 2018, 40, 1167–1179. [Google Scholar] [CrossRef]
- Pauli, T.; Fitzpatrick, J.J. Malt combing nuts as a nutrient supplement to whey permeate for producing lactic by fermentation with Lactobacillus casei. Process Biochem. 2002, 38, 1–6. [Google Scholar] [CrossRef]
- Wee, Y.J.; Ryu, H.W. Lactic acid production by Lactobacillus sp. RKY2 in a cell-recycle continuous fermentation using lignocellulosic hydrolyzates as inexpensive raw materials. Bioresour. Technol. 2009, 100, 4262–4270. [Google Scholar] [CrossRef]
- Sikder, J.; Roy, M.; Dey, P.; Pal, P. Techno-economic analysis of a membrane-integrated bioreactor system for production of lactic acid from sugarcane juice. Biochem. Eng. J. 2012, 63, 81–87. [Google Scholar] [CrossRef]
- Rivas, B.; Moldes, A.B.; Domínguez, J.M.; Parajó, J.C. Development of culture media containing spent yeast cells of Debaryomyces hansenii and corn steep liquor for lactic acid production with Lactobacillus rhamnosus. Int. J. Food Microbiol. 2004, 97, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Lima, C.; Fontes Coelho, L.; Blanco, K.; Contiero, J. Response surface optimization of D(-)-lactic acid production by Lactobacillus SMI 8 using corn steep liquor and yeast autolysate as an alternative nitrogen source. Afr. J. Biotechnol. 2009, 8, 5842–5846. [Google Scholar]
- Yu, L.; Lei, T.; Ren, X.; Pei, X.; Feng, Y. Response surface optimization of L-(+)-lactic acid production using corn steep liquor as an alternative nitrogen source by Lactobacillus rhamnosus CGMCC 1466. Biochem. Eng. J. 2008, 39, 496–502. [Google Scholar] [CrossRef]
- Pejin, J.; Radosavljevic, M.; Pribic, M.; Kocic-Tanackov, S.; Mladenovic, D.; Djukic-Vukovic, A.; Mojovic, L. Possibility of L-(+)-lactic acid fermentation using malting, brewing, and oil production by-products. Waste Manage. 2018, 79, 153–163. [Google Scholar] [CrossRef]
- Li, Y.; Wang, L.; Ju, J.; Yu, B.; Ma, Y. Efficient production of polymer-grade D-lactate by Sporolactobacillus laevolacticus DSM442 with agricultural waste cottonseed as the sole nitrogen source. Bioresour. Technol. 2013, 142, 186–191. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, B.; Li, F.; Xu, K.; Ma, C.; Tao, F.; Li, Q.; Xu, P. Highly efficient production of D-lactate by Sporolactobacillus sp. CASD with simultaneous enzymatic hydrolysis of peanut meal. Appl. Microbiol. Biotechnol. 2011, 89, 1009–1017. [Google Scholar] [CrossRef]
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C. Determination of Sugars, Byproducts, and Degradation Products in Liquid Fraction Process Samples; NREL/TP-510–42623; National Renewable Energy Laboratory: Golden, CO, USA, 2008.
- Sluiter, A.; Hames, B.; Scarlata, C.; Sluiter, J.; Templeton, D. Determination of Structural Carbohydrates and Lignin in Biomass National Renewable; NREL/TP-510–42623; National Renewable Energy Laboratory: Golden, CO, USA, 2012.
- Zhang, B.; Khushik, F.A.; Zhan, B.; Bao, J. Transformation of lignocellulose to starch-like carbohydrates by organic acid-catalyzed pretreatment and biological detoxification. Biotechnol. Bioeng. 2021, 118, 4105–4118. [Google Scholar] [CrossRef]
- Qiu, Z.; Gao, Q.; Bao, J. Engineering Pediococcus acidilactici with xylose assimilation pathway for high titer cellulosic l-lactic acid fermentation. Bioresour. Technol. 2018, 249, 9–15. [Google Scholar] [CrossRef]
- Liu, G.; Zhang, Q.; Li, H.; Qureshi, A.S.; Zhang, J.; Bao, X.; Bao, J. Dry biorefining maximizes the potentials of simultaneous saccharification and co-fermentation for cellulosic ethanol production. Biotechnol. Bioeng. 2018, 115, 60–69. [Google Scholar] [CrossRef]
- Liu, G.; Sun, J.; Zhang, J.; Tu, Y.; Bao, J. High titer L-lactic acid production from corn stover with minimum wastewater generation and techno-economic evaluation based on Aspen plus modeling. Bioresour. Technol. 2015, 198, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Ding, S.; Li, Z.; Tan, T. L-lactic acid production by Lactobacillus casei fermentation with corn steep liquor-supplemented acid-hydrolysate of soybean meal. Biotechnol. J. 2006, 1, 1453–1458. [Google Scholar] [CrossRef] [PubMed]
- John, R.P.; Nampoothiri, K.M.; Pandey, A. Fermentative production of lactic acid from biomass: An overview on process developments and future perspectives. Appl. Microbiol. Biotechnol. 2007, 74, 524–534. [Google Scholar] [CrossRef] [PubMed]
- Alves de Oliveira, R.; Komesu, A.; Vaz Rossell, C.E.; Maciel Filho, R. Challenges and opportunities in lactic acid bioprocess design—From economic to production aspects. Biochem. Eng. J. 2018, 133, 219–239. [Google Scholar] [CrossRef]
- Edwinoliver, N.G.; Thirunavukarasu, K.; Purushothaman, S.; Rose, C.; Gowthaman, M.K.; Kamini, N.R. Corn steep liquor as a nutrition adjunct for the production of Aspergillus niger lipase and hydrolysis of oils thereof. J. Agric. Food Chem. 2009, 57, 10658–10663. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Yuan, Y.J. Comprehensive quality evaluation of corn steep liquor in 2-keto-L-gluonic acid fermentation. J. Agric. Food Chem. 2011, 59, 9845–9853. [Google Scholar] [CrossRef]
- Selling, G.W.; Hojilla-Evangelista, M.P.; Hay, W.T.; Utt, K.D.; Grose, G.D. Preparation and properties of solution cast films from pilot-scale cottonseed protein isolate. Ind. Crop. Prod. 2022, 178, 114615. [Google Scholar] [CrossRef]
- Brock, S.; Kuenz, A.; Prüße, U. Impact of hydrolysis methods on the utilization of agricultural residues as nutrient source for D-lactic acid production by Sporolactobacillus inulinus. Fermentation 2019, 5, 12. [Google Scholar] [CrossRef] [Green Version]
- Tanksley, T.D., Jr.; Knabe, D.A.; Purser, K.; Zebrowska, T.; Corley, J.R. Apparent digestibility of amino acids and nitrogen in three cottonseed meals and one soybean meal. J. Anim. Sci. 1981, 52, 769–777. [Google Scholar] [CrossRef] [Green Version]
- Gao, M.T.; Hirata, M.; Toorisaka, E.; Hano, T. Study on acid-hydrolysis of spent cells for lactic acid fermentation. Biochem. Eng. J. 2006, 28, 87–91. [Google Scholar] [CrossRef]
- Nielsen, H.P.; Frandsen, F.J.; Dam-Johansen, K.; Baxter, L.L. The implications of chlorine-associated corrosion on the operation of biomass-fired boilers. Prog. Energy Combust. Sci. 2000, 26, 283–298. [Google Scholar] [CrossRef]
- He, N.; Jia, J.; Qiu, Z.; Fang, C.; Liden, G.; Liu, X.; Bao, J. Cyclic L-lactide synthesis from lignocellulose biomass by biorefining with complete inhibitor removal and highly simultaneous sugars assimilation. Biotechnol. Bioeng. 2022, 119, 1903–1915. [Google Scholar] [CrossRef] [PubMed]
- Åkerberg, C.; Zacchi, G. An economic evaluation of the fermentative production of lactic acid from wheat flour. Bioresour. Technol. 2000, 75, 119–126. [Google Scholar] [CrossRef]
- González, M.I.; Álvarez, S.; Riera, F.; Álvarez, R. Economic evaluation of an integrated process for lactic acid production from ultrafiltered whey. J. Food Eng. 2007, 80, 553–561. [Google Scholar] [CrossRef]
- Humbird, D.; Davis, R.; Tao, L.; Kinchin, C.; Hsu, D.; Aden, A.; Schoen, P.; Lukas, J.; Olthof, B.; Worley, M.; et al. Process Design and Economics for Biochemical Conversion of Lignocellulosic Biomass to Ethanol: Dilute-Acid Pretreatment and Enzymatic Hydrolysis of Corn Stover; NREL/TP-5100–47764; National Renewable Energy Laboratory: Golden, CO, USA, 2011. [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nitrogen Sources a | Dosage (g/L) | Lactic Acid Titer (g/L) | L-Lactic Acid Purity (%) | L-Lactic Acid Yield (g/g DM) b |
|---|---|---|---|---|
| YE + peptone, reagent grade | 15 for YE; 10 for peptone | 105.0 ± 0.5 | 99.5 ± 0.1 | 0.33 ± 0.01 |
| YE + peptone, industrial grade | 15 for YE; 10 for peptone | 102.0 ± 0.8 | 99.6 ± 0.1 | 0.32 ± 0.02 |
| DCSLP #1 | 20 | 87.2 ± 1.4 | 94.9 ± 0.1 | 0.26 ± 0.01 |
| DCSLP #2 | 20 | 85.4 ± 1.0 | 95.3 ± 0.1 | 0.25 ± 0.01 |
| Nitrogen a | Price b (USD/kg) | Protein (mg/g DM) | Lactic Acid (mg/g DM) | L-Lactic Acid (mg/g DM) | D-Lactic Acid (mg/g DM) |
|---|---|---|---|---|---|
| YE, reagent grade | 34.92 | 658.6 ± 1.6 | 3.3 ± 0.2 | 1.8 ± 0.1 | 1.5 ± 0.1 |
| YE, industrial grade | 17.46 | 769.3 ± 9.5 | 2.1 ± 0.1 | 0.9 ± 0.1 | 1.2 ± 0.2 |
| Peptone, reagent grade | 80.00 | 760.2 ± 14.3 | 2.0 ± 0.1 | 1.1 ± 0.1 | 0.9 ± 0.1 |
| Peptone, industrial grade | 11.11 | 728.4 ± 12.5 | ND c | ND c | ND c |
| DCSLP #1 | 1.43 | 451.2 ± 5.3 | 215.8 ± 3.5 | 85.6 ± 1.4 | 130.2 ± 2.1 |
| DCSLP #2 | 0.32 | 467.5 ± 6.1 | 196.3 ± 2.7 | 85.1 ± 0.4 | 111.2 ± 2.3 |
| Nitrogen | Price (USD/kg) | Protein (mg/g DM) | Lactic Acid (mg/g DM) | L-Lactic Acid (mg/g DM) | D-Lactic Acid (mg/g DM) |
|---|---|---|---|---|---|
| Soybean meal | 0.32 | 456.3 ± 9.5 | ND * | ND * | ND * |
| Cottonseed meal | 1.59 | 545.3 ± 11.3 | ND * | ND * | ND * |
| Case 1 [23] | Case 2 [This Study] | Case 3 [This Study] | |
|---|---|---|---|
| Strain | P. acidilactici TY112 | P. acidilactici ZY271 | P. acidilactici ZY271 |
| Xylose utilization | No | Yes | Yes |
| Raw feedstock | Corn stover | Wheat straw | Wheat straw |
| Pretreatment acid dosage | 5.0%, w/w (dry matter) | 4.1%, w/w (dry matter) | 4.1%, w/w (dry matter) |
| Fermentation solids loading | 30% (w/w) | 25% (w/w) | 25% (w/w) |
| Nitrogen sources | |||
| Complex nitrogen source a | 20 g/L DCSLP | 15 g/L YE + 10 g/L Peptone | 20 g/L cottonseed hydrolysate |
| Available nitrogen source | 2 g/L diammonium phosphate | 2 g/L diammonium hydrogen citrate | 10 g/L ammonium sulfate |
| SSCF period (h) | 72 | 72 | 72 |
| Titer (g/L) | 104.5 | 102.0 | 96.5 |
| Yield (g/g dry raw feedstock) | 0.27 | 0.33 | 0.31 |
| Productivity (g/L/h) | 1.45 | 1.46 | 1.32 |
| Chiral purity (%) | 95.3 | 99.5 | 99.7 |
| Material | USD Price (2022) |
|---|---|
| Feedstock (wheat straw) | 71.24/ton |
| Sulfuric acid, 98% | 125.06/ton |
| Lime | 99.69/ton |
| Diammonium hydrogen citrate | 3166.11/ton |
| Ammonium sulfate | 87.07/ton |
| Manganese sulfate | 443.26/ton |
| Yeast extract (YE) | 17,413.60/ton |
| Peptone | 11,081.38/ton |
| Cottonseed meal | 1266.44/ton |
| Case 1 | Case 2 | Case 3 | |
|---|---|---|---|
| Feedstock handling rate | 300,000 metric tons/year | 300,000 metric tons/year | 300,000 metric tons/year |
| Total capital investment a | USD 186 million | USD 193 million | USD 195 million |
| Lactic acid yield | 269 kg/ton corn stover | 330 kg/ton wheat straw | 310 kg/ton wheat straw |
| (95.3% L-purity) | (99.6% L-purity) | (99.7% L-purity) | |
| Plant water usage | 6. 10 kg/kg lactic acid product | 7.29 kg/kg lactic acid product | 8.10 kg/kg lactic acid product |
| Minimum lactic acid selling price (USD/kg lactic acid product) | 0.584 | 3.241 | 0.813 |
| Feedstock | 0.237 | 0.276 | 0.286 |
| Enzyme b | 0.130 | 0.101 | 0.104 |
| Complex nitrogen | 0.009 | 2.624 | 0.193 |
| Available nitrogen | 0.016 | 0.045 | 0.007 |
| Sulfuric acid c | / | / | 0.005 |
| CaCO3 d | / | / | 0.004 |
| None-enzyme conversion | 0.192 | 0.195 | 0.214 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, B.; Wu, L.; Liu, X.; Bao, J. Plant Proteins as an Alternative Nitrogen Source for Chiral Purity L-Lactic Acid Fermentation from Lignocellulose Feedstock. Fermentation 2022, 8, 546. https://doi.org/10.3390/fermentation8100546
Zhang B, Wu L, Liu X, Bao J. Plant Proteins as an Alternative Nitrogen Source for Chiral Purity L-Lactic Acid Fermentation from Lignocellulose Feedstock. Fermentation. 2022; 8(10):546. https://doi.org/10.3390/fermentation8100546
Chicago/Turabian StyleZhang, Bin, Lei Wu, Xiucai Liu, and Jie Bao. 2022. "Plant Proteins as an Alternative Nitrogen Source for Chiral Purity L-Lactic Acid Fermentation from Lignocellulose Feedstock" Fermentation 8, no. 10: 546. https://doi.org/10.3390/fermentation8100546
APA StyleZhang, B., Wu, L., Liu, X., & Bao, J. (2022). Plant Proteins as an Alternative Nitrogen Source for Chiral Purity L-Lactic Acid Fermentation from Lignocellulose Feedstock. Fermentation, 8(10), 546. https://doi.org/10.3390/fermentation8100546