
The Consumption of Amino Acids and Production of Volatile Aroma Compounds by Yarrowia lipolytica in Brewers’ Wort


Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Media
2.2. Fermentation Medium
2.3. Fermentation Conditions
2.4. Preculture and Inoculation
2.5. Analytical Methods
2.5.1. Analyses of Growth and Sugars
2.5.2. Analysis of Ethanol
2.5.3. Analysis of Amino Acids
2.5.4. Analysis of Volatile Compounds
2.6. Statistics
3. Results
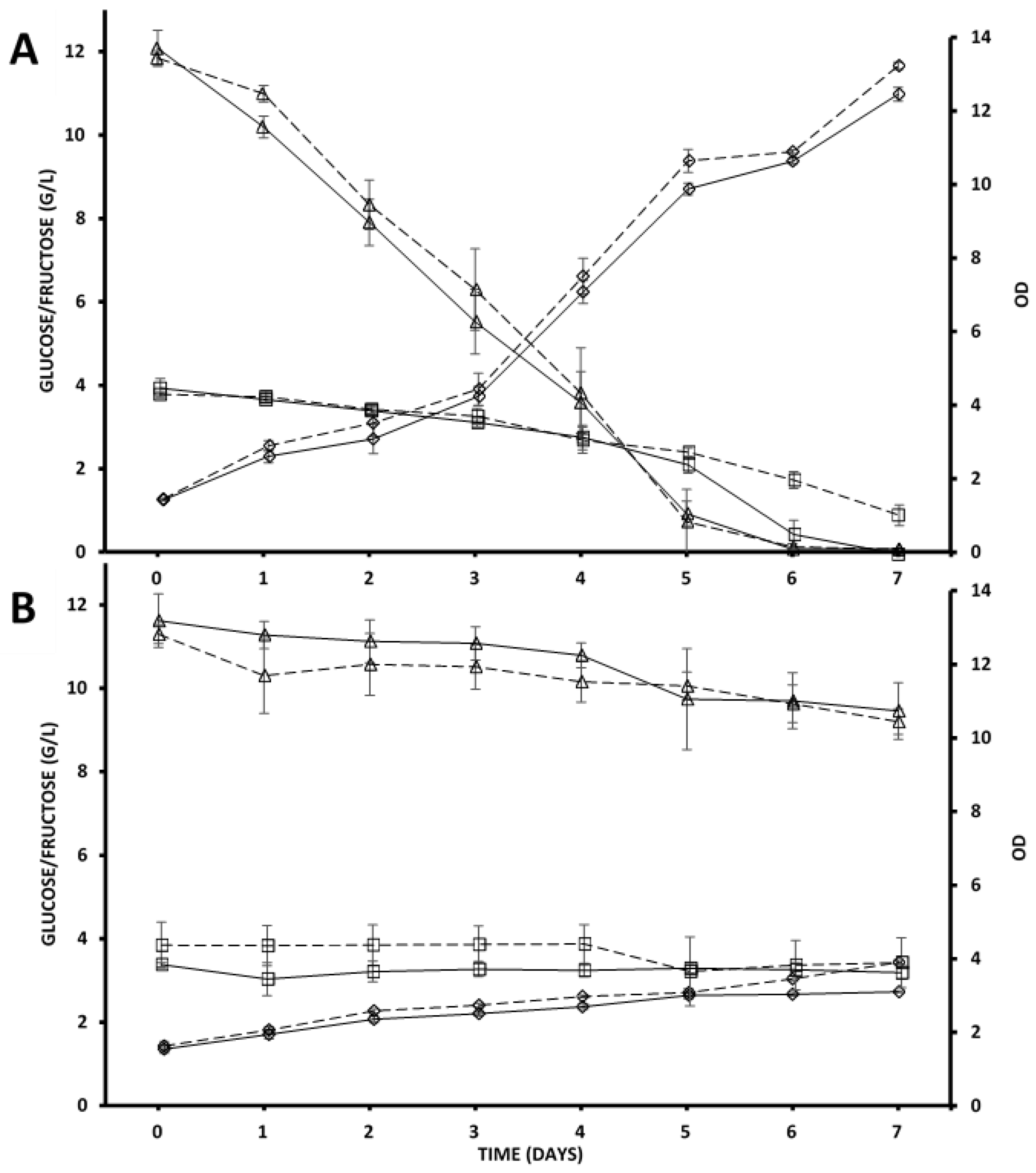
3.1. Growth, Sugar Consumption, and Ethanol Production
3.2. Consumption of Amino Acids
3.3. Production of Volatile Compounds
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yarrow, D. Four New Combinations in Yeasts. Antonie Leeuwenhoek 1972, 38, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Barth, G.; Gaillardin, C. Physiology and Genetics of the Dimorphic Fungus Yarrowia lipolytica. FEMS Microbiol. Rev. 1997, 19, 219–237. [Google Scholar] [CrossRef]
- Fickers, P.; Cheng, H.; Sze Ki Lin, C. Sugar Alcohols and Organic Acids Synthesis in Yarrowia lipolytica: Where Are We? Microorganisms 2020, 8, 574. [Google Scholar] [CrossRef] [PubMed]
- Groenewald, M.; Boekhout, T.; Neuvéglise, C.; Gaillardin, C.; Van Dijck, P.W.M.; Wyss, M. Yarrowia lipolytica: Safety Assessment of an Oleaginous Yeast with a Great Industrial Potential. Crit. Rev. Microbiol. 2014, 40, 187–206. [Google Scholar] [CrossRef] [PubMed]
- Zinjarde, S.S. Food-Related Applications of Yarrowia lipolytica. Food Chem. 2014, 152, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Coelho, M.A.; Priscilla, A.; Isabel, B. Yarrowia lipolytica: An Industrial Workhorse. In Current Research, Technology and Education Topics in Applied Microbiology and Microbial Biotechnology; Formatex: Badajoz, Spain, 2010; Volume 2. [Google Scholar]
- Nicaud, J.-M. Yarrowia lipolytica. Yeast 2012, 29, 409–418. [Google Scholar] [CrossRef]
- Sutherland, J.B.; Cornelison, C.; Crow, S.A. Yarrowia lipolytica (Candida lipolytica). In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C.A., Tortorello, M.L., Eds.; Academic Press: Cambridge, MA, USA, 2014; pp. 374–378. ISBN 978-0-12-384733-1. [Google Scholar]
- Spitaels, F.; Wieme, A.D.; Janssens, M.; Aerts, M.; Daniel, H.-M.; Van Landschoot, A.; De Vuyst, L.; Vandamme, P. The Microbial Diversity of Traditional Spontaneously Fermented Lambic Beer. PLoS ONE 2014, 9, e95384. [Google Scholar] [CrossRef]
- Spitaels, F.; Wieme, A.D.; Janssens, M.; Aerts, M.; Van Landschoot, A.; De Vuyst, L.; Vandamme, P. The Microbial Diversity of an Industrially Produced Lambic Beer Shares Members of a Traditionally Produced One and Reveals a Core Microbiota for Lambic Beer Fermentation. Food Microbiol. 2015, 49, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Dashko, S.; Zhou, N.; Compagno, C.; Piškur, J. Why, When, and How Did Yeast Evolve Alcoholic Fermentation? FEMS Yeast Res. 2014, 14, 826–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spagnuolo, M.; Shabbir Hussain, M.; Gambill, L.; Blenner, M. Alternative Substrate Metabolism in Yarrowia lipolytica. Front. Microbiol. 2018, 9, 1077. [Google Scholar] [CrossRef]
- Beopoulos, A.; Desfougeres, T.; Sabirova, J.; Zinjarde, S.; Neuvéglise, C.; Nicaud, J.M. The Hydrocarbon-Degrading Oleaginous Yeast Yarrowia lipolytica. In Handbook of Hydrocarbon and Lipid Microbiology; Springer: Berlin/Heidelberg, Germany, 2010; pp. 2111–2121. [Google Scholar] [CrossRef]
- Mansour, S.; Beckerich, J.M.; Bonnarme, P. Lactate and Amino Acid Catabolism in the Cheese-Ripening Yeast Yarrowia lipolytica. Appl. Environ. Microbiol. 2008, 74, 6505–6512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hackenschmidt, S.; Bracharz, F.; Daniel, R.; Thürmer, A.; Bruder, S.; Kabisch, J. Effects of a High-Cultivation Temperature on the Physiology of Three Different Yarrowia lipolytica Strains. FEMS Yeast Res. 2019, 19, foz068. [Google Scholar] [CrossRef]
- Liu, H.-H.; Ji, X.-J.; Huang, H. Biotechnological Applications of Yarrowia lipolytica: Past, Present and Future. Biotechnol. Adv. 2015, 33, 1522–1546. [Google Scholar] [CrossRef]
- Bankar, A.; Jadhav, L.; Phalke, V. Metagenomic Insights of Yarrowia lipolytica in Food Industry. In Metagenomic Systems Biology; Springer: Singapore, 2020; pp. 159–183. [Google Scholar] [CrossRef]
- Małajowicz, J.; Nowak, D.; Fabiszewska, A.; Iuliano, A. Comparison of Gamma-Decalactone Biosynthesis by Yeast Yarrowia lipolytica MTLY40-2p and W29 in Batch-Cultures. Biotechnol. Biotechnol. Equip. 2020, 34, 330–340. [Google Scholar] [CrossRef]
- Celińska, E.; Kubiak, P.; Białas, W.; Dziadas, M.; Grajek, W. Yarrowia lipolytica: The Novel and Promising 2-Phenylethanol Producer. J. Ind. Microbiol. Biotechnol. 2013, 40, 389–392. [Google Scholar] [CrossRef] [Green Version]
- Braga, A.; Belo, I. Production of γ-Decalactone by Yarrowia lipolytica: Insights into Experimental Conditions and Operating Mode Optimization. J. Chem. Technol. Biotechnol. 2015, 90, 559–565. [Google Scholar] [CrossRef] [Green Version]
- Kothari, S.; Vadgama, R.; Bhat, K.; Lali, A.; Anil, A. Process Optimization for Production and Purification of γ-Decalactone from Ricinoleic Acid Using Yarrowia lipolytica NCIM 3590. Biocatal. Agric. Biotechnol. 2022, 39, 102285. [Google Scholar] [CrossRef]
- Kebabci, Ö.; Cihangir, N. Comparison of Three Yarrowia lipolytica Strains for Lipase Production: NBRC 1658, IFO 1195, and a Local Strain. Turk. J. Biol. 2012, 36, 15–24. [Google Scholar] [CrossRef]
- Iucci, L.; Patrignani, F.; Belletti, N.; Ndagijimana, M.; Elisabetta Guerzoni, M.; Gardini, F.; Lanciotti, R. Role of Surface-Inoculated Debaryomyces Hansenii and Yarrowia lipolytica Strains in Dried Fermented Sausage Manufacture. Part 2: Evaluation of Their Effects on Sensory Quality and Biogenic Amine Content. Meat Sci. 2007, 75, 669–675. [Google Scholar] [CrossRef]
- Patrignani, F.; Iucci, L.; Vallicelli, M.; Guerzoni, M.E.; Gardini, F.; Lanciotti, R. Role of Surface-Inoculated Debaryomyces hansenii and Yarrowia lipolytica Strains in Dried Fermented Sausage Manufacture. Part 1: Evaluation of Their Effects on Microbial Evolution, Lipolytic and Proteolytic Patterns. Meat Sci. 2007, 75, 676–686. [Google Scholar] [CrossRef]
- Jones, M.; Pierce, J.S. Absorbtion of Amino Acids from Wort by Yeasts. J. Inst. Brew. 1964, 70, 307–315. [Google Scholar] [CrossRef]
- Liu, J.; Toldam-Andersen, T.B.; Petersen, M.A.; Zhang, S.; Arneborg, N.; Bredie, W.L.P. Instrumental and Sensory Characterisation of Solaris White Wines in Denmark. Food Chem. 2015, 166, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Johnsen, L.G.; Skou, P.B.; Khakimov, B.; Bro, R. Gas Chromatography—Mass Spectrometry Data Processing Made Easy. J. Chromatogr. A 2017, 1503, 57–64. [Google Scholar] [CrossRef]
- Methner, Y.; Hutzler, M.; Matoulková, D.; Jacob, F.; Michel, M. Screening for the Brewing Ability of Different Non-Saccharomyces Yeasts. Fermentation 2019, 5, 101. [Google Scholar] [CrossRef] [Green Version]
- Hough, J.S.; Briggs, D.E.; Stevens, R.; Young, T.W. Metabolism of Wort by Yeast. In Malting and Brewing Science: Volume II Hopped Wort and Beer; Springer: Boston, MA, USA, 1982; pp. 566–614. ISBN 978-1-4615-1799-3. [Google Scholar]
- D’Amore, T.; Russell, I.; Stewart, G.G. Sugar Utilization by Yeast during Fermentation. J. Ind. Microbiol. Biotechnol. 1989, 4, 315–323. [Google Scholar] [CrossRef]
- Moeller, L.; Zehnsdorf, A.; Aurich, A.; Bley, T.; Strehlitz, B. Substrate Utilization by Recombinant Yarrowia lipolytica Growing on Sucrose. Appl. Microbiol. Biotechnol. 2012, 93, 1695–1702. [Google Scholar] [CrossRef]
- Ferreira, I.M.; Guido, L.F. Impact of Wort Amino Acids on Beer Flavour: A Review. Fermentation 2018, 4, 23. [Google Scholar] [CrossRef] [Green Version]
- Verstrepen, K.J.; Van Laere, S.D.M.; Vanderhaegen, B.M.P.; Derdelinckx, G.; Dufour, J.-P.; Pretorius, I.S.; Winderickx, J.; Thevelein, J.M.; Delvaux, F.R. Expression Levels of the Yeast Alcohol Acetyltransferase Genes ATF1, Lg-ATF1, and ATF2 Control the Formation of a Broad Range of Volatile Esters. Appl. Environ. Microbiol. 2003, 69, 5228–5237. [Google Scholar] [CrossRef]
- Verbelen, P.J.; Saerens, S.M.; Van Mulders, S.E.; Delvaux, F.; Delvaux, F.R. The Role of Oxygen in Yeast Metabolism during High Cell Density Brewery Fermentations. Appl. Microbiol. Biotechnol. 2009, 82, 1143–1156. [Google Scholar] [CrossRef]
- Fujii, T.; Kobayashi, O.; Yoshimoto, H.; Furukawa, S.; Tamai, Y. Effect of Aeration and Unsaturated Fatty Acids on Expression of the Saccharomyces cerevisiae Alcohol Acetyltransferase Gene. Appl. Environ. Microbiol. 1997, 63, 910–915. [Google Scholar] [CrossRef] [Green Version]
- Celińska, E.; Olkowicz, M.; Grajek, W. L-Phenylalanine Catabolism and 2-Phenylethanol Synthesis in Yarrowia lipolytica—Mapping Molecular Identities through Whole-Proteome Quantitative Mass Spectrometry Analysis. FEMS Yeast Res. 2015, 15, fov041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, D.; Xu, P. Recent Advances in Biotechnological Production of 2-Phenylethanol. Biotechnol. Adv. 2011, 29, 654–660. [Google Scholar] [CrossRef] [PubMed]
- Etschmann, M.M.W.; Bluemke, W.; Sell, D.; Schrader, J. Biotechnological Production of 2-Phenylethanol. Appl. Microbiol. Biotechnol. 2002, 59, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Ma, J.; Zhu, Y.; Xu, P. Refactoring Ehrlich Pathway for High-Yield 2-Phenylethanol Production in Yarrowia lipolytica. ACS Synth. Biol. 2020, 9, 623–633. [Google Scholar] [CrossRef] [Green Version]
- Hazelwood, L.A.; Daran, J.-M.; van Maris, A.J.A.; Pronk, J.T.; Dickinson, J.R. The Ehrlich Pathway for Fusel Alcohol Production: A Century of Research on Saccharomyces cerevisiae Metabolism. Appl. Environ. Microbiol. 2008, 74, 2259–2266. [Google Scholar] [CrossRef] [Green Version]
- Dufour, J.-P.; Malcorps, P.H.; Silcock, P. Control of Ester Synthesis During Brewery Fermentation. In Brewing Yeast Fermentation Performance, 2nd ed.; Smart, K., Ed.; Blackwell Publishing: Oxford, UK, 2008; pp. 213–233. [Google Scholar] [CrossRef]
- Saerens, S.M.G.; Verbelen, P.J.; Vanbeneden, N.; Thevelein, J.M.; Delvaux, F.R. Monitoring the Influence of High-Gravity Brewing and Fermentation Temperature on Flavour Formation by Analysis of Gene Expression Levels in Brewing Yeast. Appl. Microbiol. Biotechnol. 2008, 80, 1039–1051. [Google Scholar] [CrossRef]
- Saerens, S.M.G.; Delvaux, F.; Verstrepen, K.J.; Van Dijck, P.; Thevelein, J.M.; Delvaux, F.R. Parameters Affecting Ethyl Ester Production by Saccharomyces cerevisiae during Fermentation. Appl. Environ. Microbiol. 2008, 74, 454–461. [Google Scholar] [CrossRef]
- Landaud, S.; Latrille, E.; Corrieu, G. Top Pressure and Temperature Control the Fusel Alcohol/Ester Ratio through Yeast Growth in Beer Fermentation. J. Inst. Brew. 2001, 107, 107–117. [Google Scholar] [CrossRef]
- Lehnert, R.; Kuřec, M.; Brányik, T. Effect of Oxygen Supply on Flavor Formation during Continuous Alcohol-Free Beer Production: A Model Study. J. Am. Soc. Brew. Chem. 2008, 66, 233–238. [Google Scholar] [CrossRef] [Green Version]
- Gomes, N.; Aguedo, M.; Teixeira, J.; Belo, I. Oxygen Mass Transfer in a Biphasic Medium: Influence on the Biotransformation of Methyl Ricinoleate Into γ-Decalactone by the Yeast Yarrowia lipolytica. Biochem. Eng. J. 2007, 35, 380–386. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| High Aeration | Low Aeration | |||
|---|---|---|---|---|
| 20 °C | 30 °C | 20 °C | 30 °C | |
| Day 0 | 2.40 ± 0.11 (0%) | 2.38 ± 0.03 (0%) | 2.30 ± 0.10 (0%) | 2.32 ± 0.01 (0%) |
| Day 4 | 0.93 ± 0.08 (61.1%) | 0.89 ± 0.08 (62.6%) | 2.00 ± 0.11 (13.2%) | 2.21 ± 0.06 (4.7%) |
| Day 7 | 0.60 ± 0.04 (74.9%) | 0.48 ± 0.04 (79.7%) | 1.73 ± 0.04 (24.9%) | 1.94 ± 0.05 (16.4%) |
| High Aeration | Low Aeration | |||
|---|---|---|---|---|
| 20 °C | 30 °C | 20 °C | 30 °C | |
| Group 1 Fast More than 80% on day 4 | Proline Alanine Lysine Arginine Asparagine Aspartic Acid Glutamic Acid Glycine | Proline Alanine Arginine Serine Asparagine Aspartic Acid Glutamic Acid Glycine | Aspartic Acid Glutamic Acid | Aspartic Acid |
| Group 2 Intermediate Between 50% and 80% on day 4 | Serine Methionine | Lysine Methionine | Glutamic Acid | |
| Group 3 Slow More than 10% on day 7 and less than 50% on day 4 | Leucine Phenylalanine Valine Tyrosine Isoleucine Histidine Tryptophan Glutamine | Leucine Phenylalanine Valine Tyrosine Isoleucine Threonine Histidine Tryptophan Glutamine | Proline Alanine Arginine Serine Asparagine Glutamine | Proline Alanine Arginine Serine Asparagine |
| Group 4 Little or no absorption Less than 10% on day 7 | Threonine | - | Leucine Phenylalanine Valine Lysine Tyrosine Isoleucine Threonine Histidine Tryptophan Glycine Methionine | Leucine Phenylalanine Valine Lysine Tyrosine Isoleucine Threonine Histidine Tryptophan Glycine Methionine Glutamine |
| High Aeration | Low Aeration | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 20 °C | 30 °C | 20 °C | 30 °C | |||||||||||
| Day | 0 | 4 | 7 | 0 | 4 | 7 | 0 | 4 | 7 | 0 | 4 | 7 | ||
| Esters | ||||||||||||||
| Not detected | ||||||||||||||
| Higher Alcohols | ||||||||||||||
| 1-propanol | ||||||||||||||
| 2-methyl-1-propanol | ||||||||||||||
| 3-methyl-1-butanol | ||||||||||||||
| 3-methyl-3-buten-1-ol | ||||||||||||||
| 1-pentanol | ||||||||||||||
| 1-hexanol | ||||||||||||||
| (E)-2-nonen-1-ol | ||||||||||||||
| 2-ethyl-1-hexanol | ||||||||||||||
| 1-octanol | ||||||||||||||
| propylene glycol | ||||||||||||||
| α,α-dimethyl-benzenemethanol | ||||||||||||||
| 2-phenylethanol | ||||||||||||||
| 2-dodecyl 1,3-propanediol | ||||||||||||||
| Phenolic Compounds | ||||||||||||||
| 2-methoxy-4-vinylphenol | ||||||||||||||
| 2-naphthalenol | ||||||||||||||
| phenol | ||||||||||||||
| Aldehydes | ||||||||||||||
| 3-methyl-butanal | ||||||||||||||
| 2-methyl-propanal | ||||||||||||||
| 2-methyl-butanal | ||||||||||||||
| hexanal | ||||||||||||||
| heptanal | ||||||||||||||
| 3-methyl-2-butenal | ||||||||||||||
| octanal | ||||||||||||||
| decanal | ||||||||||||||
| (E)-2-nonenal | ||||||||||||||
| Ketones | ||||||||||||||
| acetone | ||||||||||||||
| 2-butanone | ||||||||||||||
| 2,3-butanedione | ||||||||||||||
| 2,3-pentanedione | ||||||||||||||
| cyclopentanone | ||||||||||||||
| 5-methyl-2-hexanone | ||||||||||||||
| acetoin | ||||||||||||||
| 1-hydroxy-2-propanone | ||||||||||||||
| 9-oxabicyclo [6.1.0]nonan-4-one | ||||||||||||||
| 6-methyl-5-hepten-2-one | ||||||||||||||
| 4-Cyclopentene-1,3-dione | ||||||||||||||
| Acetophenone | ||||||||||||||
| β-damascenone | ||||||||||||||
| geranylacetone | ||||||||||||||
| Fatty Acids | ||||||||||||||
| acetic acid | ||||||||||||||
| formic acid | ||||||||||||||
| propanoic acid | ||||||||||||||
| 2-methyl-propanoic acid | ||||||||||||||
| butanoic acid | ||||||||||||||
| 3-methyl-butanoic acid | ||||||||||||||
| pentanoic acid | ||||||||||||||
| hexanoic acid | ||||||||||||||
| heptanoic acid | ||||||||||||||
| octanoic acid | ||||||||||||||
| nonanoic acid | ||||||||||||||
| n-decanoic acid | ||||||||||||||
| benzoic acid | ||||||||||||||
| Benzene Derivates | ||||||||||||||
| (1-methylethyl)-benzene | ||||||||||||||
| 1,2,4-trimethyl-benzene | ||||||||||||||
| 1,3-bis(1,1-dimethylethyl)-benzene | ||||||||||||||
| benzaldehyde | ||||||||||||||
| benzeneacetaldehyde | ||||||||||||||
| Furans | ||||||||||||||
| 2-pentyl-furan | ||||||||||||||
| dihydro-2-methyl-3(2H)-furanone | ||||||||||||||
| furfural | ||||||||||||||
| 1-(2-furanyl)-ethanone | ||||||||||||||
| 2-furanmethanol | ||||||||||||||
| Others | ||||||||||||||
| methanesulfonyl chloride | ||||||||||||||
| thiazole | ||||||||||||||
| α-methylstyrene | ||||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sørensen, A.B.; Petersen, M.A.; Garde, A.; Arneborg, N. The Consumption of Amino Acids and Production of Volatile Aroma Compounds by Yarrowia lipolytica in Brewers’ Wort. Fermentation 2022, 8, 579. https://doi.org/10.3390/fermentation8110579
Sørensen AB, Petersen MA, Garde A, Arneborg N. The Consumption of Amino Acids and Production of Volatile Aroma Compounds by Yarrowia lipolytica in Brewers’ Wort. Fermentation. 2022; 8(11):579. https://doi.org/10.3390/fermentation8110579
Chicago/Turabian StyleSørensen, Anders Bagger, Mikael Agerlin Petersen, Arvid Garde, and Nils Arneborg. 2022. "The Consumption of Amino Acids and Production of Volatile Aroma Compounds by Yarrowia lipolytica in Brewers’ Wort" Fermentation 8, no. 11: 579. https://doi.org/10.3390/fermentation8110579
APA StyleSørensen, A. B., Petersen, M. A., Garde, A., & Arneborg, N. (2022). The Consumption of Amino Acids and Production of Volatile Aroma Compounds by Yarrowia lipolytica in Brewers’ Wort. Fermentation, 8(11), 579. https://doi.org/10.3390/fermentation8110579