

Integrating 1G with 2G Bioethanol Production by Using Distillers’ Dried Grains with Solubles (DDGS) as the Feedstock for Lignocellulolytic Enzyme Production



Abstract
:1. Introduction

2. The Main Differences in 1G and 2G Bioethanol Production
3. Mechanism of Enzymatic Hydrolysis of 1G Feedstock
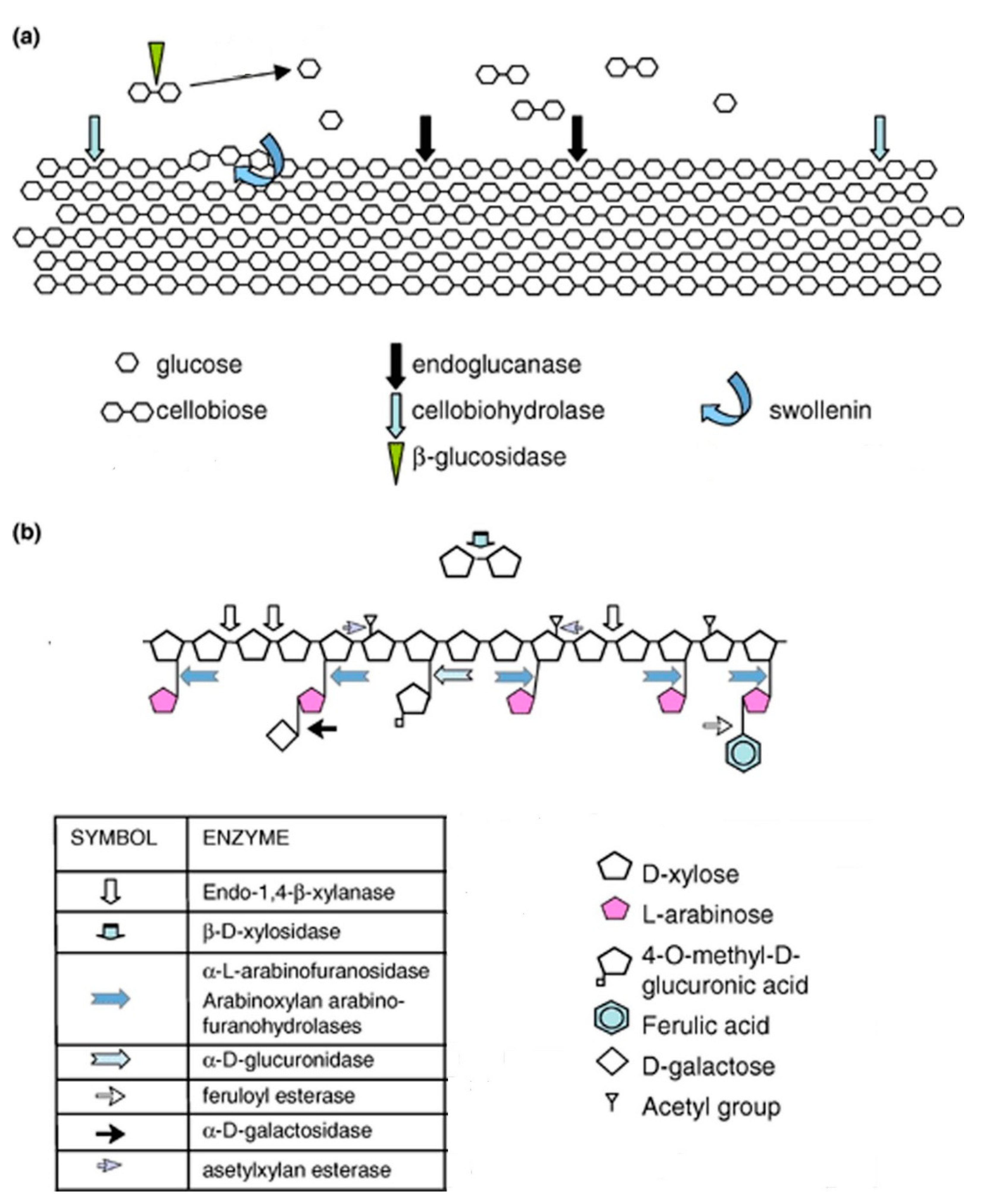
4. Mechanism of Enzymatic Degradation of Lignocellulosic Biomass

5. Lignocellulolytic Enzyme Productions
6. Pretreatment for Enzyme Production
7. Optimization of Enzyme Production
8. On-Site Lignocellulolytic Enzyme Production for 2G Bioethanol Production
9. Integration of 1G and 2G Bioethanol Refineries
10. Concluding Remarks and Future Trends
Funding
Conflicts of Interest
References
- Aui, A.; Wang, Y.; Mba-Wright, M. Evaluating the economic feasibility of cellulosic ethanol: A meta-analysis of techno-economic analysis studies. Renew. Sustain. Energy Rev. 2021, 145, 111098. [Google Scholar] [CrossRef]
- Miklautsch, P.; Woschank, M. A framework of measures to mitigate greenhouse gas emissions in freight transport: Systematic literature review from a Manufacturer’s perspective. J. Clean. Prod. 2022, 366, 132883. [Google Scholar] [CrossRef]
- Edenhofer, O.; Pichs-Madruga, R.; Sokona, Y.; Agrawala, S.; Bashmakov, I.A.; Blanco, G.; Broome, J.; Bruckner, T.; Brunner, S.; Bustamante, M. Summary for policymakers. In Climate Change 2014: Mitigation of Climate Change; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Hoang, T.-D.; Nghiem, N. Recent developments and current status of commercial production of fuel ethanol. Fermentation 2021, 7, 314. [Google Scholar] [CrossRef]
- Raj, T.; Chandrasekhar, K.; Naresh Kumar, A.; Rajesh Banu, J.; Yoon, J.-J.; Kant Bhatia, S.; Yang, Y.-H.; Varjani, S.; Kim, S.-H. Recent advances in commercial biorefineries for lignocellulosic ethanol production: Current status, challenges and future perspectives. Bioresour. Technol. 2022, 344, 126292. [Google Scholar] [CrossRef]
- Niphadkar, S.; Bagade, P.; Ahmed, S. Bioethanol production: Insight into past, present and future perspectives. Biofuels 2018, 9, 229–238. [Google Scholar] [CrossRef]
- Bertrand, E.; Vandenberghe, L.P.S.; Soccol, C.R.; Sigoillot, J.-C.; Faulds, C. First generation bioethanol. In Green Fuels Technology; Springer: Berlin/Heidelberg, Germany, 2016; pp. 175–212. [Google Scholar]
- Wang, M.; Wu, M.; Huo, H. Life-cycle energy and greenhouse gas emission impacts of different corn ethanol plant types. Environ. Res. Lett. 2007, 2, 24001. [Google Scholar] [CrossRef]
- Qaisar, K.; Nawaz, A.; Akram, F.; Mukhtar, H.; Zohu, X.; Xu, Y.; Mumtaz, M.W.; Rashid, U.; Ghani, W.A.W.A.K.; Choong, T.S.Y. Advances in valorization of lignocellulosic biomass towards energy generation. Catalysts 2021, 11, 309. [Google Scholar]
- Wu, X.; Luo, N.; Xie, S.; Zhang, H.; Zhang, Q.; Wang, F.; Wang, Y. Photocatalytic transformations of lignocellulosic biomass into chemicals. Chem. Soc. Rev. 2020, 49, 6198–6223. [Google Scholar] [CrossRef]
- Iram, A.; Cekmecelioglu, D.; Demirci, A. Ideal Feedstock and Fermentation Process Improvements for the Production of Lignocellulolytic Enzymes. Processes 2021, 9, 38. [Google Scholar] [CrossRef]
- Iram, A.; Berenjian, A.; Demirci, A. A review on the utilization of lignin as a fermentation substrate to produce lignin-modifying enzymes and other value-added products. Molecules 2021, 26, 2960. [Google Scholar] [CrossRef]
- Fan, W.; Sun, X.; Cui, G.; Li, Q.; Xu, Y.; Wang, L.; Li, X.; Hu, B.; Chi, Z. A strategy of co-fermentation of distillers dried grains with solubles (DDGS) and lignocellulosic feedstocks as swine feed. Crit. Rev. Biotechnol. 2022, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Iram, A.; Cekmecelioglu, D.; Demirci, A. Distillers’ Dried Grains with Solubles (DDGS) and its Potential as the Fermentation Feedstock. Appl. Microbiol. Biotechnol. 2020, 104, 6115–6128. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.V.; Hamid, S.B.A.; Zain, S.K. Conversion of lignocellulosic biomass to nanocellulose: Structure and chemical process. Sci. World J. 2014, 2014, 631013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Susmozas, A.; Martín-Sampedro, R.; Ibarra, D.; Eugenio, M.E.; Iglesias, R.; Manzanares, P.; Moreno, A.D. Process strategies for the transition of 1G to advanced bioethanol production. Processes 2020, 8, 1310. [Google Scholar] [CrossRef]
- Kwiatkowski, J.R.; McAloon, A.J.; Taylor, F.; Johnston, D.B. Modeling the process and costs of fuel ethanol production by the corn dry-grind process. Ind. Crops Prod. 2006, 23, 288–296. [Google Scholar] [CrossRef]
- Guo, Y.; Liu, G.; Ning, Y.; Li, X.; Hu, S.; Zhao, J.; Qu, Y. Production of cellulosic ethanol and value-added products from corn fiber. Bioresour. Bioprocess. 2022, 9, 1–18. [Google Scholar] [CrossRef]
- Gurunath, R.B.; Gobinath, R.; Giridhar, P.V. Bioethanol Production from Lignocellulosic Biomass: Past, Present and Future Trends. Res. J. Biotechnol. 2022, 17, 10. [Google Scholar]
- Kumar, B.; Bhardwaj, N.; Agrawal, K.; Verma, P. Bioethanol production: Generation-based comparative status measurements. In Biofuel Production Technologies: Critical Analysis for Sustainability; Springer: Berlin/Heidelberg, Germany, 2020; pp. 155–201. [Google Scholar]
- Badiei, M.; Asim, N.; Jahim, J.M.; Sopian, K. Comparison of chemical pretreatment methods for cellulosic biomass. APCBEE Procedia 2014, 9, 170–174. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.S.; Lee, Y.Y.; Kim, T.H. A review on alkaline pretreatment technology for bioconversion of lignocellulosic biomass. Bioresour. Technol. 2016, 199, 42–48. [Google Scholar] [CrossRef]
- McAloon, A.; Taylor, F.; Yee, W.; Ibsen, K.; Wooley, R. Determining the Cost of Producing Ethanol from Corn Starch and Lignocellulosic Feedstocks; National Renewable Energy Lab.(NREL): Golden, CO, USA, 2000. [Google Scholar]
- Hii, S.L.; Tan, J.S.; Ling, T.C.; Ariff, A. Bin Pullulanase: Role in starch hydrolysis and potential industrial applications. Enzym. Res. 2012, 2012, 921362. [Google Scholar] [CrossRef] [Green Version]
- Zabed, H.; Sahu, J.N.; Boyce, A.N.; Faruq, G. Fuel ethanol production from lignocellulosic biomass: An overview on feedstocks and technological approaches. Renew. Sustain. Energy Rev. 2016, 66, 751–774. [Google Scholar] [CrossRef]
- Barbosa, F.C.; Silvello, M.A.; Goldbeck, R. Cellulase and oxidative enzymes: New approaches, challenges and perspectives on cellulose degradation for bioethanol production. Biotechnol. Lett. 2020, 42, 875–884. [Google Scholar] [CrossRef]
- Balat, M.; Balat, H.; Öz, C. Progress in bioethanol processing. Prog. Energy Combust. Sci. 2008, 34, 551–573. [Google Scholar] [CrossRef]
- Aro, N.; Pakula, T.; Penttilä, M. Transcriptional regulation of plant cell wall degradation by filamentous fungi. FEMS Microbiol. Rev. 2005, 29, 719–739. [Google Scholar] [CrossRef] [Green Version]
- Houfani, A.A.; Anders, N.; Spiess, A.C.; Baldrian, P.; Benallaoua, S. Insights from enzymatic degradation of cellulose and hemicellulose to fermentable sugars–a review. Biomass Bioenergy 2020, 134, 105481. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, Y.; Cheng, H.; Zhou, H. Hemicellulose degradation: An overlooked issue in acidic deep eutectic solvents pretreatment of lignocellulosic biomass. Ind. Crops Prod. 2022, 187, 115335. [Google Scholar] [CrossRef]
- Sukumaran, R.K.; Singhania, R.R.; Pandey, A. Microbial cellulases-production, applications and challenges. J. Sci. Ind. Res. 2005, 64, 890–896. [Google Scholar]
- Pant, S.; Nag, P.; Ghati, A.; Chakraborty, D.; Maximiano, M.R.; Franco, O.L.; Mandal, A.K.; Kuila, A. Employment of the CRISPR/Cas9 system to improve cellulase production in Trichoderma reesei. Biotechnol. Adv. 2022, 60, 108022. [Google Scholar] [CrossRef]
- Srivastava, N.; Hussain, A.; Kushwaha, D.; Haque, S.; Mishra, P.K.; Gupta, V.K.; Srivastava, M. Nickel ferrite nanoparticles induced improved fungal cellulase production using residual algal biomass and subsequent hydrogen production following dark fermentation. Fuel 2021, 304, 121391. [Google Scholar] [CrossRef]
- Lodha, A.; Pawar, S.; Rathod, V. Optimised cellulase production from fungal co-culture of Trichoderma reesei NCIM 1186 and Penicillium citrinum NCIM 768 under solid state fermentation. J. Environ. Chem. Eng. 2020, 8, 103958. [Google Scholar] [CrossRef]
- Singhal, A.; Kumari, N.; Ghosh, P.; Singh, Y.; Garg, S.; Shah, M.P.; Jha, P.K.; Chauhan, D.K. Optimizing cellulase production from Aspergillus flavus using response surface methodology and machine learning models. Environ. Technol. Innov. 2022, 27, 102805. [Google Scholar] [CrossRef]
- Vázquez-Montoya, E.L.; Castro-Ochoa, L.D.; Maldonado-Mendoza, I.E.; Luna-Suárez, S.; Castro-Martínez, C. Moringa straw as cellulase production inducer and cellulolytic fungi source. Rev. Argent. De Microbiol. 2020, 52, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Xiang, J.; Wang, X.; Sang, T. Cellulase production from Trichoderma reesei RUT C30 induced by continuous feeding of steam-exploded Miscanthus lutarioriparius. Ind. Crops Prod. 2021, 160, 113129. [Google Scholar] [CrossRef]
- Manglekar, R.R.; Geng, A. CRISPR-Cas9-mediated seb1 disruption in Talaromyces pinophilus EMU for its enhanced cellulase production. Enzym. Microb. Technol. 2020, 140, 109646. [Google Scholar] [CrossRef]
- Han, L.; Tan, Y.; Ma, W.; Niu, K.; Hou, S.; Guo, W.; Liu, Y.; Fang, X. Precision engineering of the transcription factor Cre1 in Hypocrea jecorina (Trichoderma reesei) for efficient cellulase production in the presence of glucose. Front. Bioeng. Biotechnol. 2020, 8, 852. [Google Scholar] [CrossRef]
- Goukanapalle, P.K.R.; Kanderi, D.K.; Rajoji, G.; Kumari, S.; Bontha, R.R. Optimization of cellulase production by a novel endophytic fungus Pestalotiopsis microspora TKBRR isolated from Thalakona forest. Cellulose 2020, 27, 6299–6316. [Google Scholar] [CrossRef]
- Raheja, Y.; Kaur, B.; Falco, M.; Tsang, A.; Chadha, B.S. Secretome analysis of Talaromyces emersonii reveals distinct CAZymes profile and enhanced cellulase production through response surface methodology. Ind. Crops Prod. 2020, 152, 112554. [Google Scholar] [CrossRef]
- Xue, Y.; Han, J.; Li, Y.; Liu, J.; Gan, L.; Long, M. Promoting cellulase and hemicellulase production from Trichoderma orientalis EU7-22 by overexpression of transcription factors Xyr1 and Ace3. Bioresour. Technol. 2020, 296, 122355. [Google Scholar] [CrossRef]
- Luo, Y.; Valkonen, M.; Jackson, R.E.; Palmer, J.M.; Bhalla, A.; Nikolaev, I.; Saloheimo, M.; Ward, M. Modification of transcriptional factor ACE3 enhances protein production in Trichoderma reesei in the absence of cellulase gene inducer. Biotechnol. Biofuels 2020, 13, 137. [Google Scholar] [CrossRef]
- Kaur, J.; Chugh, P.; Soni, R.; Soni, S.K. A low-cost approach for the generation of enhanced sugars and ethanol from rice straw using in-house produced cellulase-hemicellulase consortium from A. niger P-19. Bioresour. Technol. Rep. 2020, 11, 100469. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, J.; Luo, L.; Wang, E.; Wang, R.; Liu, L.; Liu, J.; Yuan, H. Low-cost cellulase-hemicellulase mixture secreted by Trichoderma harzianum EM0925 with complete saccharification efficacy of lignocellulose. Int. J. Mol. Sci. 2020, 21, 371. [Google Scholar] [CrossRef] [Green Version]
- Faria, S.P.; de Melo, G.R.; Cintra, L.C.; Ramos, L.P.; Jesuino, R.S.A.; Ulhoa, C.J.; de Faria, F.P. Production of cellulases and xylanases by Humicola grisea var. thermoidea and application in sugarcane bagasse arabinoxylan hydrolysis. Ind. Crops Prod. 2020, 158, 112968. [Google Scholar] [CrossRef]
- Pramanik, S.K.; Mahmud, S.; Paul, G.K.; Jabin, T.; Naher, K.; Uddin, M.S.; Zaman, S.; Saleh, M.A. Fermentation optimization of cellulase production from sugarcane bagasse by Bacillus pseudomycoides and molecular modeling study of cellulase. Curr. Res. Microb. Sci. 2021, 2, 100013. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Xie, Y.; Gao, X.; Shan, M.; Sun, C.; Niu, Y.D.; Shan, A. Screening of cellulose degradation bacteria from Min pigs and optimization of its cellulase production. Electron. J. Biotechnol. 2020, 48, 29–35. [Google Scholar] [CrossRef]
- Pham, V.H.T.; Kim, J.; Shim, J.; Chang, S.; Chung, W. Coconut mesocarp-based lignocellulosic waste as a substrate for cellulase production from high promising multienzyme-producing Bacillus amyloliquefaciens FW2 without pretreatments. Microorganisms 2022, 10, 327. [Google Scholar] [CrossRef]
- Mohamad Zabidi, N.A.; Foo, H.L.; Loh, T.C.; Mohamad, R.; Abdul Rahim, R. Enhancement of versatile extracellular cellulolytic and hemicellulolytic enzyme productions by Lactobacillus plantarum RI 11 isolated from Malaysian food using renewable natural polymers. Molecules 2020, 25, 2607. [Google Scholar] [CrossRef]
- Beladhadi, R.V.; Shankar, K.; Jayalakshmi, S.K.; Sreeramulu, K. Production of Cocktail of Lignolytic, Cellulolytic and Hemicellulolytic Enzymes by the Novel Bacterium Burkholderia sp SMB1 Utilizing Rice Bran and Straw: Application in the Saccharification of Untreated Agro-wastes for Bioethanol Production. Waste Biomass Valoriz. 2022, 13, 1565–1577. [Google Scholar] [CrossRef]
- Fujii, T.; Inoue, H.; Ishikawa, K. Enhancing cellulase and hemicellulase production by genetic modification of the carbon catabolite repressor gene, creA, in Acremonium cellulolyticus. AMB Express 2013, 3, 73. [Google Scholar] [CrossRef] [Green Version]
- Satari, B.; Karimi, K.; Kumar, R. Cellulose solvent-based pretreatment for enhanced second-generation biofuel production: A review. Sustain. Energy Fuels 2019, 3, 11–62. [Google Scholar] [CrossRef]
- Ghosh, S.; Chowdhury, R.; Bhattacharya, P. Sustainability of cereal straws for the fermentative production of second generation biofuels: A review of the efficiency and economics of biochemical pretreatment processes. Appl. Energy 2017, 198, 284–298. [Google Scholar] [CrossRef]
- Vancov, T.; McIntosh, S. Alkali pretreatment of cereal crop residues for second-generation biofuels. Energy Fuels 2011, 25, 2754–2763. [Google Scholar] [CrossRef]
- Wagner, A.O.; Lackner, N.; Mutschlechner, M.; Prem, E.M.; Markt, R.; Illmer, P. Biological pretreatment strategies for second-generation lignocellulosic resources to enhance biogas production. Energies 2018, 11, 1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rastogi, M.; Shrivastava, S. Recent advances in second generation bioethanol production: An insight to pretreatment, saccharification and fermentation processes. Renew. Sustain. Energy Rev. 2017, 80, 330–340. [Google Scholar] [CrossRef]
- Iram, A.; Cekmecelioglu, D.; Demirci, A. Optimization of dilute sulfuric acid, aqueous ammonia, and steam explosion as the pretreatments steps for distillers’ dried grains with solubles as a potential fermentation feedstock. Bioresour. Technol. 2019, 282, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Sharma, H.K.; Xu, C.; Qin, W. Biological pretreatment of lignocellulosic biomass for biofuels and bioproducts: An overview. Waste Biomass Valoriz. 2019, 10, 235–251. [Google Scholar] [CrossRef]
- Sindhu, R.; Binod, P.; Pandey, A. Biological pretreatment of lignocellulosic biomass - An overview. Bioresour. Technol. 2016, 199, 76–82. [Google Scholar] [CrossRef]
- Binod, P.; Gnansounou, E.; Sindhu, R.; Pandey, A. Enzymes for second generation biofuels: Recent developments and future perspectives. Bioresour. Technol. Rep. 2019, 5, 317–325. [Google Scholar] [CrossRef]
- Juhasz, T.; Szengyel, Z.; Reczey, K.; Siika-Aho, M.; Viikari, L. Characterization of cellulases and hemicellulases produced by Trichodermareesei on various carbon sources. Process Biochem. 2005, 40, 3519–3525. [Google Scholar] [CrossRef]
- Xin, F.; Geng, A. Horticultural waste as the substrate for cellulase and hemicellulase production by Trichoderma reesei under solid-state fermentation. Appl. Biochem. Biotechnol. 2010, 162, 295–306. [Google Scholar] [CrossRef]
- Kang, S.W.; Park, Y.S.; Lee, J.S.; Hong, S.I.; Kim, S.W. Production of cellulases and hemicellulases by Aspergillus niger KK2 from lignocellulosic biomass. Bioresour. Technol. 2004, 91, 153–156. [Google Scholar] [CrossRef]
- Rajoka, M.I.; Malik, K.A. Cellulase and Hemicellulase production by Cellulomonas flavigena NIAB 441. Biotechnol. Lett. 1984, 6, 597–600. [Google Scholar] [CrossRef]
- Iram, A.; Cekmecelioglu, D.; Demirci, A. Screening of bacterial and fungal strains for cellulase and xylanase production using distillers’ dried grains with solubles (DDGS) as the main feedstock. Biomass Convers. Biorefin. 2020, 11, 1955–1964. [Google Scholar] [CrossRef]
- Jampala, P.; Tadikamalla, S.; Preethi, M.; Ramanujam, S.; Uppuluri, K.B. Concurrent production of cellulase and xylanase from Trichoderma reesei NCIM 1186: Enhancement of production by desirability-based multi-objective method. 3 Biotech 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ahamed, A.; Vermette, P. Culture-based strategies to enhance cellulase enzyme production from Trichoderma reesei RUT-C30 in bioreactor culture conditions. Biochem. Eng. J. 2008, 40, 399–407. [Google Scholar] [CrossRef]
- Ahamed, A.; Vermette, P. Effect of culture medium composition on Trichoderma reesei’s morphology and cellulase production. Bioresour. Technol. 2009, 100, 5979–5987. [Google Scholar] [CrossRef]
- Abdullah, J.J.; Greetham, D.; Pensupa, N.; Tucker, G.A.; Du, C. Optimizing cellulase production from municipal solid waste (MSW) using solid state fermentation (SSF). J. Fundam. Renew. Energy Appl. 2016, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Saratale, G.D.; Kshirsagar, S.D.; Sampange, V.T.; Saratale, R.G.; Oh, S.-E.; Govindwar, S.P.; Oh, M.-K. Cellulolytic enzymes production by utilizing agricultural wastes under solid state fermentation and its application for biohydrogen production. Appl. Biochem. Biotechnol. 2014, 174, 2801–2817. [Google Scholar] [CrossRef]
- Iram, A.; Cekmecelioglu, D.; Demirci, A. Salt and nitrogen amendment and optimization for cellulase and xylanase production using dilute acid hydrolysate of distillers’ dried grains with solubles (DDGS) as the feedstock. Bioprocess Biosyst. Eng. 2022, 45, 527–540. [Google Scholar] [CrossRef]
- Kachlishvili, E.; Penninckx, M.J.; Tsiklauri, N.; Elisashvili, V. Effect of nitrogen source on lignocellulolytic enzyme production by white-rot basidiomycetes under solid-state cultivation. World J. Microbiol. Biotechnol. 2006, 22, 391–397. [Google Scholar] [CrossRef]
- Iram, A.; Cekmecelioglu, D.; Demirci, A. Optimization of the fermentation parameters to maximize the production of cellulases and xylanases using DDGS as the main feedstock in stirred tank bioreactors. Biocatal. Agric. Biotechnol. 2022, 45, 102514. [Google Scholar] [CrossRef]
- Limkar, M.B.; Pawar, S. V; Rathod, V.K. Statistical optimization of xylanase and alkaline protease co-production by Bacillus spp using Box-Behnken Design under submerged fermentation using wheat bran as a substrate. Biocatal. Agric. Biotechnol. 2019, 17, 455–464. [Google Scholar] [CrossRef]
- Soni, R.; Nazir, A.; Chadha, B.S. Optimization of cellulase production by a versatile Aspergillus fumigatus fresenius strain (AMA) capable of efficient deinking and enzymatic hydrolysis of Solka floc and bagasse. Ind. Crops Prod. 2010, 31, 277–283. [Google Scholar] [CrossRef]
- Slade, R.; Bauen, A.; Shah, N. The greenhouse gas emissions performance of cellulosic ethanol supply chains in Europe. Biotechnol. Biofuels 2009, 2, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Kazi, F.K.; Fortman, J.A.; Anex, R.P.; Hsu, D.D.; Aden, A.; Dutta, A.; Kothandaraman, G. Techno-economic comparison of process technologies for biochemical ethanol production from corn stover. Fuel 2010, 89, S20–S28. [Google Scholar] [CrossRef]
- Sørensen, A.; Teller, P.J.; Lübeck, P.S.; Ahring, B.K. Onsite enzyme production during bioethanol production from biomass: Screening for suitable fungal strains. Appl. Biochem. Biotechnol. 2011, 164, 1058–1070. [Google Scholar] [CrossRef] [PubMed]
- de Souza, M.F.; da Silva Bon, E.P.; da Silva, A.S. Production of cellulases and β-glucosidases by Trichoderma reesei Rut C30 using steam-pretreated sugarcane bagasse: An integrated approach for onsite enzyme production. Braz. J. Chem. Eng. 2021, 38, 435–442. [Google Scholar] [CrossRef]
- Singh, A.; Bishnoi, N.R. Optimization of enzymatic hydrolysis of pretreated rice straw and ethanol production. Appl. Microbiol. Biotechnol. 2012, 93, 1785–1793. [Google Scholar] [CrossRef]
- Cavka, A.; Alriksson, B.; Rose, S.H.; Van Zyl, W.H.; Jönsson, L.J. Production of cellulosic ethanol and enzyme from waste fiber sludge using SSF, recycling of hydrolytic enzymes and yeast, and recombinant cellulase-producing Aspergillus niger. J. Ind. Microbiol. Biotechnol. 2014, 41, 1191–1200. [Google Scholar] [CrossRef]
- Cheng, M.-H.; Wang, Z.; Dien, B.S.; Slininger, P.J.W.; Singh, V. Economic analysis of cellulosic ethanol production from sugarcane bagasse using a sequential deacetylation, hot water and disk-refining pretreatment. Processes 2019, 7, 642. [Google Scholar] [CrossRef] [Green Version]
- Idris, A.S.O.; Pandey, A.; Rao, S.S.; Sukumaran, R.K. Cellulase production through solid-state tray fermentation, and its use for bioethanol from sorghum stover. Bioresour. Technol. 2017, 242, 265–271. [Google Scholar] [CrossRef]
- Maeda, R.N.; Barcelos, C.A.; Santa Anna, L.M.M.; Pereira Jr, N. Cellulase production by Penicillium funiculosum and its application in the hydrolysis of sugar cane bagasse for second generation ethanol production by fed batch operation. J. Biotechnol. 2013, 163, 38–44. [Google Scholar] [CrossRef] [Green Version]
- Johnson, E. Integrated enzyme production lowers the cost of cellulosic ethanol. Biofuels Bioprod. Biorefin. 2016, 10, 164–174. [Google Scholar] [CrossRef] [Green Version]
- Sharma, B.; Larroche, C.; Dussap, C.-G. Comprehensive assessment of 2G bioethanol production. Bioresour. Technol. 2020, 313, 123630. [Google Scholar] [CrossRef] [PubMed]
- Qiao, J.; Cui, H.; Wang, M.; Fu, X.; Wang, X.; Li, X.; Huang, H. Integrated biorefinery approaches for the industrialization of cellulosic ethanol fuel. Bioresour. Technol. 2022, 360, 127516. [Google Scholar] [CrossRef] [PubMed]
- Macrelli, S.; Mogensen, J.; Zacchi, G. Techno-economic evaluation of 2nd generation bioethanol production from sugar cane bagasse and leaves integrated with the sugar-based ethanol process. Biotechnol. Biofuels 2012, 5, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Ayodele, B.V.; Alsaffar, M.A.; Mustapa, S.I. An overview of integration opportunities for sustainable bioethanol production from first-and second-generation sugar-based feedstocks. J. Clean. Prod. 2020, 245, 118857. [Google Scholar] [CrossRef]
- Lawford, H.G.; Rousseau, J.D. Cellulosic fuel ethanol. In Biotechnology for Fuels and Chemicals; Springer: Berlin/Heidelberg, Germany, 2003; pp. 457–469. [Google Scholar]
- Erdei, B.; Hancz, D.; Galbe, M.; Zacchi, G. SSF of steam-pretreated wheat straw with the addition of saccharified or fermented wheat meal in integrated bioethanol production. Biotechnol. Biofuels 2013, 6, 1–14. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Parameter | 1G | 2G | Advantage of 1G over 2G or Vice-Versa | References |
|---|---|---|---|---|
| Feedstock | Sugar or starch based: corn, wheat, sugarcane, beet, etc. | Lignocellulosic biomass: inedible parts of the plant, straw, wood, and sawdust, etc. | Sugar- or starch-based feedstocks are easy to breakdown into simple monosaccharides which are then easily fermented into ethanol. | [19] |
| Pretreatment | Liquefaction | Dilute acid, steam, AFEX, etc. | Liquefaction requires low to medium process conditions as compared to lignocellulosic pretreatment strategies. | [2,3] |
| Enzymatic hydrolysis | Saccharification with amylases and glucoamylases | Lignocellulolytic process with cellulases and hemicellulases | Saccharification has high conversion yield as compared to the cellulose and hemicellulose hydrolysis | [16] |
| Cost of feedstock | 40–70% | 30% | 2G is advantageous as the feedstock is inexpensive. | [16] |
| Pretreatment cost | Low | High | 1G is advantageous as low capital investment is needed. | [16] |
| Reduction in GHG emissions | 39–52% | 86% | 2G ethanol is more sustainable as compared to 1G ethanol. | [1] |
| Food vs. fuel issue | Yes | No | 2G ethanol is advantageous over 1G for food security. | [20] |
| Microorganism | Strain | Reference |
|---|---|---|
| Fungi | Trichoderma reesei | [32] |
| Cladosporium cladosporioides NS2 | [33] | |
| Trichoderma reesei NCIM 1186 and Penicillium citrinum NCIM 768 | [34] | |
| Aspergillus flavus | [35] | |
| Penicillium sp., Cladosporium sp., Fusarium sp. | [36] | |
| Trichoderma reesei RUT C30 | [37] | |
| Talaromyces pinophilus EMU | [38] | |
| Trichoderma reesei | [39] | |
| Pestalotiopsis microspora TKBRR | [40] | |
| Talaromyces emersonii | [41] | |
| Trichoderma orientalis EU7-22 | [42] | |
| Trichoderma reesei | [43] | |
| Aspergillus niger | [44] | |
| Trichoderma harzianum EM0925 | [45] | |
| Humicola grisea var. thermoidea | [46] | |
| Bacteria | Bacillus pseudomycoides | [47] |
| Bacillus velezensis | [48] | |
| Bacillus amyloliquefaciens FW2 | [49] | |
| Lactobacillus plantarum RI 11 | [50] | |
| Burkholderia sp. | [51] |
| Research Aspect | Feedstock | Main Results | References |
|---|---|---|---|
| Screening for fungal strains | Wet oxidized wheat straw and filter cake | Twenty-five out of sixty-four fungal strains were selected for their cellulolytic activities. | [79] |
| Enzyme activities on the substrate and glucose yield | Steam-pretreated sugarcane bagasse | Cellulase activity (1.93 FPU/mL) and β-glucosidase activity (0.37 BGU/mL) were obtained. Glucose yield was 80%. | [80] |
| Extent of enzyme production | Microwave alkali-pretreated rice straw | Cellulase activity (24 FPU/gds), xylanase activity (258 IU/gds) and β-glucosidase activity (3.8 IU/gds) were obtained. | [81] |
| Extent of enzyme production | Spent fiber sludge hydrolysates | Cellulase activity of 2700 to 2900 nkat/mL was obtained. | [82] |
| Production cost and energy analysis of 2G ethanol | Sugarcane bagasse | The minimum selling price of ethanol varies from 4.91 USD to 4.52 USD/gal. | [83] |
| Greenhouse gas emissions reductions | N/A | On-site enzyme production further decreases the greenhouse gas emissions. | [77] |
| Extent of enzyme production and hydrolysis | Wheat bran and cellulose | The produced cellulase hydrolyzed alkali pretreated sorghum stover which was fermented to ethanol with approximately 80% efficiency. | [84] |
| Extent of enzyme production and hydrolysis | Sugar cane bagasse | The fermentation efficiency to ethanol of 78% was achieved with the on-site enzyme blends. | [85] |
| Technoeconomic analysis | Same as 2G ethanol | Production cost of 2G ethanol decreased by 19% with on-site enzyme production scenario. | [86] |
| Technoeconomic analysis | Corn stover | The product value (PV) of 2G ethanol was estimated to be 1.42 USD/LGE. | [78] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iram, A.; Cekmecelioglu, D.; Demirci, A. Integrating 1G with 2G Bioethanol Production by Using Distillers’ Dried Grains with Solubles (DDGS) as the Feedstock for Lignocellulolytic Enzyme Production. Fermentation 2022, 8, 705. https://doi.org/10.3390/fermentation8120705
Iram A, Cekmecelioglu D, Demirci A. Integrating 1G with 2G Bioethanol Production by Using Distillers’ Dried Grains with Solubles (DDGS) as the Feedstock for Lignocellulolytic Enzyme Production. Fermentation. 2022; 8(12):705. https://doi.org/10.3390/fermentation8120705
Chicago/Turabian StyleIram, Attia, Deniz Cekmecelioglu, and Ali Demirci. 2022. "Integrating 1G with 2G Bioethanol Production by Using Distillers’ Dried Grains with Solubles (DDGS) as the Feedstock for Lignocellulolytic Enzyme Production" Fermentation 8, no. 12: 705. https://doi.org/10.3390/fermentation8120705
APA StyleIram, A., Cekmecelioglu, D., & Demirci, A. (2022). Integrating 1G with 2G Bioethanol Production by Using Distillers’ Dried Grains with Solubles (DDGS) as the Feedstock for Lignocellulolytic Enzyme Production. Fermentation, 8(12), 705. https://doi.org/10.3390/fermentation8120705