
Bioprocess Strategy of Haematococcus lacustris for Biomass and Astaxanthin Production Keys to Commercialization: Perspective and Future Direction

 ,
,  , , ,
, , ,  , and
, and 
Abstract
:1. Introduction
2. Composition Biochemical of Microalgae Haematococcus lacustris
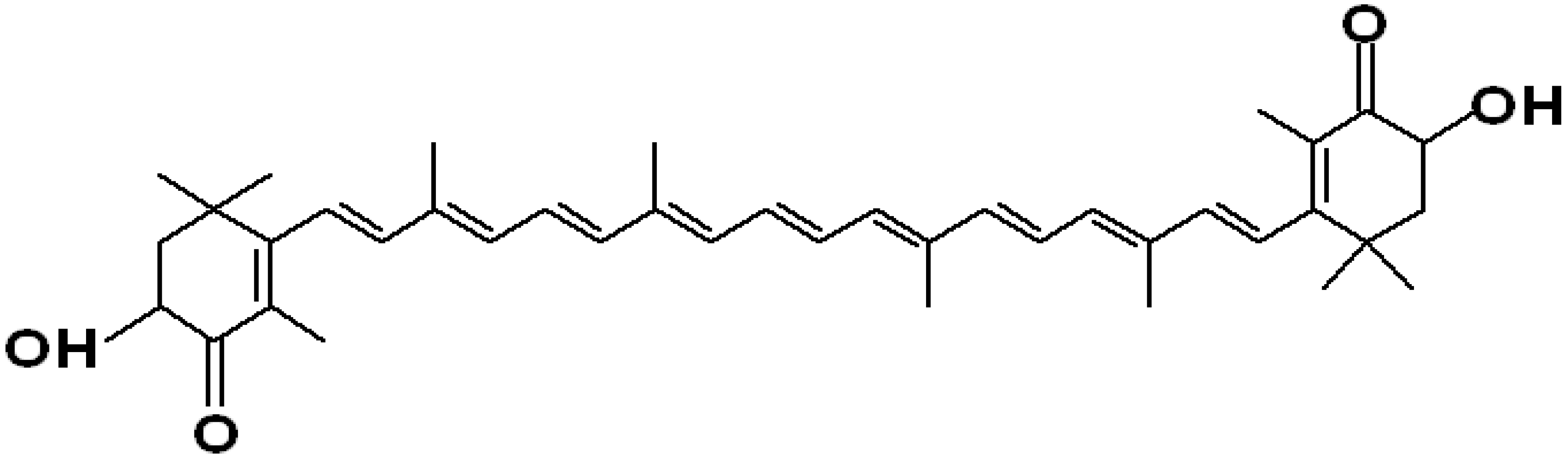
2.1. The Astaxanthin Molecules
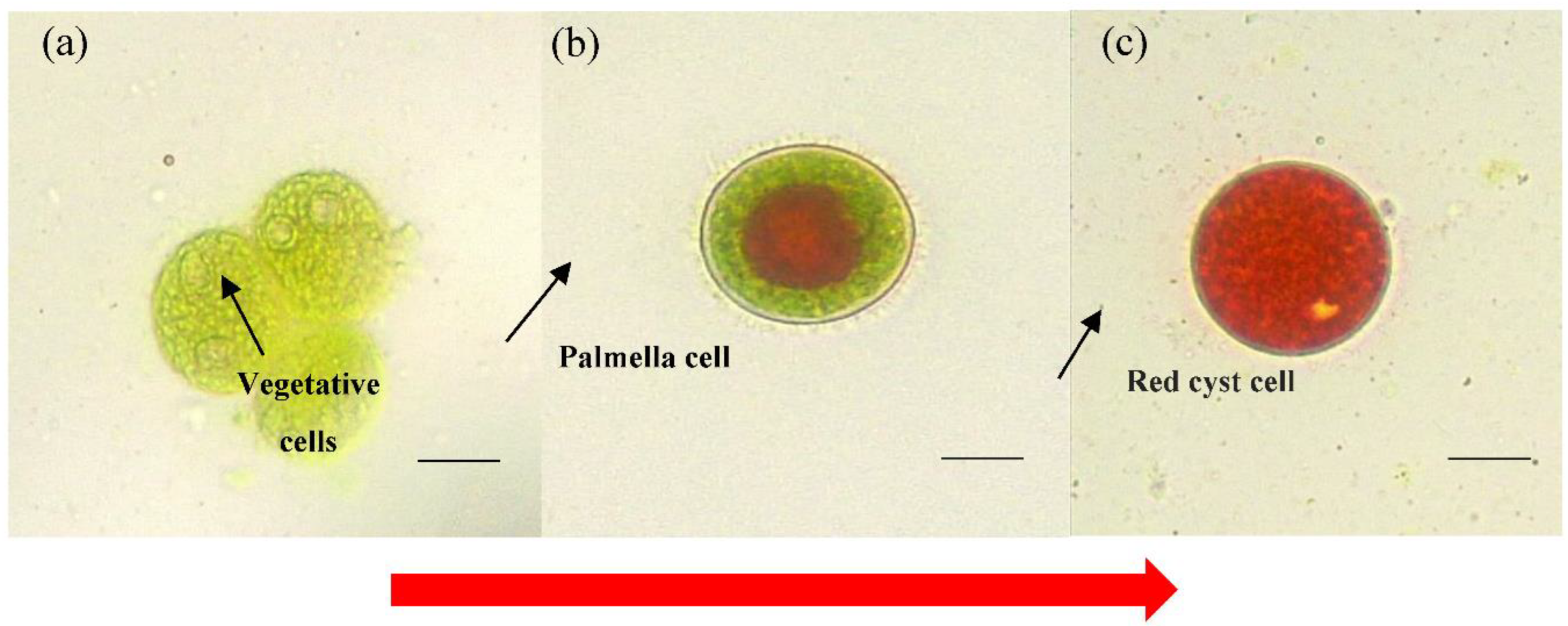
2.2. Haematococcus lacustris
3. Microalgae Cultivation Strategies for Production of H. lacustris
3.1. Open Cultivation Systems
3.2. Closed Cultivation Systems
3.3. Cultivation of H. lacustris
4. Influence of Stress Factor on Biomass Growth and Astaxanthin Accumulation of H. lacustris
4.1. Light Intensity
4.2. Temperature
4.3. Different Types of Nutrients
4.4. Initial Biomass Density
5. Perspective and Future Direction
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Oslan, S.N.H.; Shoparwe, N.F.; Yusoff, A.H.; Rahim, A.A.; Chang, C.S.; Tan, J.S.; Oslan, S.N.; Arumugam, K.; Ariff, A.B.; Sulaiman, A.Z.; et al. A Review on Haematococcus pluvialis Bioprocess Optimization of Green and Red Stage Culture Conditions for the Production of Natural Astaxanthin. Biomolecules 2021, 11, 256. [Google Scholar] [CrossRef] [PubMed]
- Khazi, M.I.; Shi, L.; Liaqat, F.; Yang, Y.; Li, X.; Yang, D.; Li, J. Sequential Continuous Mixotrophic and Phototrophic Cultivation Might Be a Cost-Effective Strategy for Astaxanthin Production from the Microalga Haematococcus lacustris. Front. Bioeng. Biotechnol. 2021, 5, 740533. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhou, X.; Zhang, Y.; Cheng, P.; Ma, R.; Cheng, W.; Chu, H. Enhancing Astaxanthin Accumulation in Haematococcus pluvialis by Coupled Light Intensity and Nitrogen Starvation in Column Photo-bioreactors. J. Microbiol. Biotechnol. 2018, 28, 2019–2028. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Chen, X.; Too, H.P. Microbial astaxanthin biosynthesis: Recent achievements, challenges, and commercialization outlook. Appl. Microbiol. Biotechnol. 2020, 104, 5725–5737. [Google Scholar] [CrossRef]
- Zhuge, F.; Ni, Y.; Wan, C.; Liu, F.; Fu, Z. Anti-diabetic effects of astaxanthin on an STZ-induced diabetic model in rats. Endocr. J. 2020, 68, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Capelli, B.; Talbott, S.; Ding, L. Astaxanthin sources: Suitability for human health and nutrition. Funct. Foods Health Dis. 2019, 9, 430. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Zhang, N.; Zhang, Y.; Lian, Q.; Qin, C.; Qian, Z.; Wu, Y.; Yang, Z.; Li, C.; Huang, X.; et al. NaCl Promotes the Efficient Formation of Haematococcus pluvialis Nonmotile Cells under Phosphorus Deficiency. Mar. Drugs 2021, 19, 337. [Google Scholar] [CrossRef]
- Li, F.; Cai, M.; Lin, M.; Huang, X.; Wang, J.; Zheng, X.; Wu, S.; An, Y. Accumulation of Astaxanthin Was Improved by the Nonmotile Cells of Haematococcus pluvialis. BioMed Res. Int. 2019, 2019, 8101762. [Google Scholar] [CrossRef] [Green Version]
- Panis, G.; Carreon, J.R. Commercial astaxanthin production derived by green alga Haematococcus pluvialis: A microalgae process model and a techno-economic assessment all through production line. Algal Res. 2016, 18, 175–190. [Google Scholar] [CrossRef] [Green Version]
- Narala, R.R.; Garg, S.; Sharma, K.K.; Thomas-Hall, S.R.; Deme, M.; Li, Y.; Schenk, P.M. Comparison of Microalgae Cultivation in Photobioreactor, Open Raceway Pond, and a Two-Stage Hybrid System. Front. Energy Res. 2016, 4, 29. [Google Scholar] [CrossRef]
- Borowiak, D.; Pstrowska, K.; Wiśniewski, M.; Grzebyk, M. Propagation of Inoculum for Haematococcus pluvialis Microalgae Scale-Up Photobioreactor Cultivation System. Appl. Sci. 2020, 10, 6283. [Google Scholar] [CrossRef]
- Mehariya, S.; Sharma, N.; Iovine, A.; Casella, P.; Marino, T.; Larocca, V.; Molino, A.; Musmarra, D. An Integrated Strategy for Nutraceuticals from Haematoccus pluvialis: From Cultivation to Extraction. Antioxidants 2020, 9, 825. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, D.S.V.; Sant’Anna, C. Impact of culture conditions on the chlorophyll content of microalgae for biotechnological applications. World J. Microbiol. Biotechnol. 2016, 33, 20. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Wang, Z.; Ding, Y.; Geng, Y.; Li, Y. Enhancing the production of astaxanthin by mixotrophic cultivation of Haematococcus pluvialis in open raceway ponds. Aquacult. Int. 2020, 28, 625–638. [Google Scholar] [CrossRef]
- Rodríguez-Sifuentes, L.; Marszalek, J.E.; Hernández-Carbajal, G.; Chuck-Hernández, C. Importance of Downstream Processing of Natural Astaxanthin for Pharmaceutical Application. Front. Chem. Eng. 2021, 2, 601483. [Google Scholar] [CrossRef]
- Jannel, S.; Caro, Y.; Bermudes, M.; Petit, T. Novel Insights into the Biotechnological Production of Haematococcus pluvialis-Derived Astaxanthin: Advances and Key Challenges to Allow Its Industrial Use as Novel Food Ingredient. J. Mar. Sci. Eng. 2020, 8, 789. [Google Scholar] [CrossRef]
- Oslan, S.N.H.; Tan, J.S.; Oslan, S.N.; Matanjun, P.; Mokhtar, R.A.M.; Shapawi, R.; Huda, N. Haematococcus pluvialis as a Potential Source of Astaxanthin with Diverse Applications in Industrial Sectors: Current Research and Future Directions. Molecules 2021, 26, 6470. [Google Scholar] [CrossRef]
- Shah, M.M.R.; Liang, Y.; Cheng, J.J.; Daroch, M. Astaxanthin-producing green microalga Haematococcus pluvialis: From singlecell to high value commercial products. Front. Plant Sci. 2016, 7, 531. [Google Scholar] [CrossRef] [Green Version]
- Bauer, A.; Minceva, M. Examination of Photo-, Mixo-, and Heterotrophic Cultivation Conditions on Haematococcus pluvialis Cyst Cell Germination. Appl. Sci. 2021, 11, 7201. [Google Scholar] [CrossRef]
- Xi, T.; Kim, D.G.; Roh, S.W.; Choi, J.-S.; Choi, Y.-E. Enhancement of astaxanthin production using Haematococcus pluvialis with novel LED wavelength shift strategy. Appl. Microbiol. Biotechnol. 2016, 100, 6231–6238. [Google Scholar] [CrossRef]
- Li, F.; Cai, M.; Lin, M.; Huang, X.; Wang, J.; Ke, H.; Wang, C.; Zheng, X.; Chen, D.; Yang, S. Enhanced Biomass and Astaxanthin Production of Haematococcus pluvialis by a Cell Transformation Strategy with Optimized Initial Biomass Density. Mar. Drugs 2020, 18, 341. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.-E.; Hwang, S.K.; Chang, W.S.; Kim, B.W.; Lee, J.; Sim, S.J. Enhanced autotrophic astaxanthin production from Haematococcus pluvialis under high temperature via heat stress-driven Haber–Weiss reaction. Appl. Microbiol. Biotechnol. 2015, 99, 5203–5215. [Google Scholar] [CrossRef] [PubMed]
- Gu, W.; Li, H.; Zhao, P.; Yu, R.; Pan, G.; Gao, S.; Xie, X.; Huang, A.; He, L.; Wang, G. Quantitative proteomic analysis of thylakoid from two microalgae (Haematococcus pluvialis and Dunaliella salina) reveals two different high light-responsive strategies. Sci. Rep. 2014, 4, 6661. [Google Scholar] [CrossRef] [PubMed]
- Butler, T.O.; McDougall, G.J.; Campbell, R.; Stanley, M.S.; Day, J.G. Media Screening for Obtaining Haematococcus pluvialis Red Motile Macrozooids Rich in Astaxanthin and Fatty Acids. Biology 2017, 7, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, C.H.; Tan, X.; Ho, S.-H.; Lam, S.S.; Show, P.L.; Nguyen, T.H.P. Conceptual design of a hybrid thin layer cascade photo-bioreactor for microalgal biodiesel synthesis. Int. J. 2020, 44, 9757–9771. [Google Scholar]
- Wan, M.; Zhang, J.; Hou, D.; Fan, J.; Li, Y.; Huang, J.; Wang, J. The effect of temperature on cell growth and astaxanthin accumulation of Haematococcus pluvialis during a light–dark cyclic cultivation. Bioresour. Technol. 2014, 167, 276–283. [Google Scholar] [CrossRef]
- Costa, J.A.V.; de Morais, M.G. An open pond system for microalgal cultivation. In Biofuels from Algae; Pandey, A., Lee, D.-J., Chisti, Y., Soccol, C.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 1–22. [Google Scholar]
- Liyanaarachchi, V.C.; Premaratne, M.; Ariyadasa, T.U.; Nimarshana, P.H.V.; Malik, A. Two-stage cultivation of microalgae for production of high-value compounds and biofuels: A review. Algal Res. 2021, 57, 102353. [Google Scholar] [CrossRef]
- Fasaei, F.; Bitter, J.H.; Slegers, P.M.; van Boxtel, A.J.B. Techno-economic evaluation of microalgae harvesting and dewatering systems. Algal Res. 2018, 31, 347–362. [Google Scholar] [CrossRef]
- Do, T.-T.; Ong, B.-N.; Nguyen Tran, M.-L.; Nguyen, D.; Melkonian, M.; Tran, H.-D. Biomass and Astaxanthin Productivities of Haematococcus pluvialis in an Angled Twin-Layer Porous Substrate Photobioreactor: Effect of Inoculum Density and Storage Time. Biology 2019, 8, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.Y.; Geng, Y.H.; Li, Z.K.; Hu, H.J.; Li, Y.G. Production of astaxanthin from Haematococcus in open pond by two-stage growth one-step process. Aquaculture 2009, 295, 275–281. [Google Scholar] [CrossRef]
- Aflalo, C.; Meshulam, Y.; Zarka, A.; Boussiba, S. On the relative efficiency of two- vs. one-stage production of astaxanthin by the green alga Haematococcus pluvialis. Biotechnol. Bioeng. 2007, 98, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Deschênes, J.-S.; Boudreau, A.; Tremblay, R. Mixotrophic production of microalgae in pilot-scale photobioreactors: Practicability and process considerations. Algal Res. 2015, 10, 80–86. [Google Scholar] [CrossRef]
- Hu, J.; Nagarajan, D.; Zhang, Q.; Chang, J.-S.; Lee, D.-J. Heterotrophic cultivation of microalgae for pigment production: A review. Biotechnol. Adv. 2018, 36, 54–67. [Google Scholar] [CrossRef] [PubMed]
- Posten, C. Design principles of photo-bioreactors for cultivation of microalgae. Eng. Life Sci. 2009, 9, 165–177. [Google Scholar] [CrossRef]
- Kiperstok, A.C.; Sebestyén, P.; Podola, B.; Melkonian, M. Biofilm cultivation of Haematococcus pluvialis enables a highly productive one-phase process for astaxanthin production using high light intensities. Algal Res. 2017, 21, 213–222. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, J.; Wang, J.; Liu, T. Attached cultivation of Haematococcus pluvialis for astaxanthin production. Bioresour. Technol. 2014, 158, 329–335. [Google Scholar] [CrossRef]
- Tran, H.D.; Do, T.T.; Le, T.L.; Tran-Nguyen, M.L.; Pham, C.H.; Melkonian, M. Cultivation of Haematococcus pluvialis for astaxanthin production on angled bench-scale and large-scale biofilm-based photo-bioreactors. Vietnam J. Sci. Technol. Eng. 2019, 61, 60–69. [Google Scholar] [CrossRef]
- Do, T.-T.; Ong, B.-N.; Le, T.-L.; Nguyen, T.-C.; Tran-Thi, B.-H.; Thu Hien, B.T.; Melkonian, M.; Tran, H.-D. Growth of Haematococcus pluvialis on a Small-Scale Angled Porous Substrate Photobioreactor for Green Stage Biomass. Appl. Sci. 2021, 11, 1788. [Google Scholar] [CrossRef]
- Wan, M.; Zhang, Z.; Wang, J.; Huang, J.; Fan, J.; Yu, A.; Wang, W.; Li, Y. Sequential Heterotrophy–Dilution–Photoinduction Cultivation of Haematococcus pluvialis for efficient production of astaxanthin. Bioresour. Technol. 2015, 198, 557–563. [Google Scholar] [CrossRef]
- Sipaúba-Tavaresa, L.H.; Tedesquea, M.G.; Scardoeli-Truzzi, B. Evaluation of the effects of sugarcane molasses as a carbon source for Ankistrodesmus gracilis and Haematococcus pluvialis (Chlorophyceae). Braz. J. Biol. 2020, 80, 594–600. [Google Scholar] [CrossRef] [Green Version]
- Park, J.C.; Choi, S.P.; Hong, M.E.; Sim, S.J. Enhanced astaxanthin production from microalga, Haematococcus pluvialis by two-stage perfusion culture with stepwise light irradiation. Bioprocess Biosyst. Eng. 2014, 37, 2039–2047. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhou, W.; Shao, H.; Liu, T. A comparative analysis of biomass and lipid content in five Tribonema sp. strains at autotrophic, heterotrophic and mixotrophic cultivation. Algal Res. 2017, 24, 284–289. [Google Scholar] [CrossRef]
- Wang, S.K.; Wang, X.; Tao, H.H.; Sun, X.S.; Tian, Y.T. Heterotrophic culture of Chlorella pyrenoidosa using sucrose as the sole carbon source by co-culture with immobilized yeast. Bioresour. Technol. 2018, 249, 425–430. [Google Scholar] [CrossRef]
- Saha, S.K.; McHugh, E.; Hayes, J.; Moane, S.; Walsh, D.; Murray, P. Effect of various stress-regulatory factors on biomass and lipid production in microalga Haematococcus pluvialis. Bioresour. Technol. 2013, 128, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.Y.; Hong, M.E.; Jin, E.S.; Woo, H.M.; Sim, S.J. Improvement in modular scalability of polymeric thin-film photo-bioreactor for autotrophic culturing of Haematococcus pluvialis using industrial flue gas. Bioresour. Technol. 2018, 249, 519–526. [Google Scholar] [CrossRef]
- Singh, S.P.; Singh, P. Effect of temperature and light on the growth of algae species: A review. Renew. Sust. Energy Rev. 2015, 50, 431–444. [Google Scholar] [CrossRef]
- Choi, Y.Y.; Joun, J.M.; Lee, J.; Hong, M.E.; Pham, H.-M.; Chang, W.S.; Sim, S.J. Development of large-scale and economic pH control system for outdoor cultivation of microalgae Haematococcus pluvialis using industrial flue gas. Bioresour. Technol. 2017, 244, 1235–1244. [Google Scholar] [CrossRef]
- Zhao, Y.; Yue, C.; Geng, S.; Ning, D.; Ma, T.; Yu, X. Role of media composition in biomass and astaxanthin production of Haematococcus pluvialis under two-stage cultivation. Bioprocess Biosyst. Eng. 2019, 42, 593–602. [Google Scholar] [CrossRef]
- Cheng, J.; Li, K.; Yang, Z.; Lu, H.; Zhou, J.; Cen, K. Gradient domestication of Haematococcus pluvialis mutant with 15% CO2 to promote biomass growth and astaxanthin yield. Bioresour. Technol. 2016, 216, 340–344. [Google Scholar] [CrossRef]
- Cheng, J.; Li, K.; Zhu, Y.; Yang, W.; Zhou, J.; Cen, K. Transcriptome sequencing and metabolic pathways of astaxanthin accumulated in Haematococcus pluvialis mutant under 15% CO2. Bioresour. Technol. 2017, 228, 99–105. [Google Scholar] [CrossRef]
- Cheah, W.Y.; Show, P.L.; Juan, J.C.; Chang, J.S.; Ling, T.C. Enhancing biomass and lipid productions of microalgae in palm oil mill effluent using carbon and nutrient supplementation. Energy Convers. Manag. 2018, 164, 188–197. [Google Scholar] [CrossRef]
- Piasecka, A.; Krzeminska, I.; Tys, J. Enrichment of Parachlorella kessleri biomass with bio-products: Oil and protein by utilization of beet molasses. J. Appl. Phycol. 2017, 29, 1735–1743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Han, D.; Sommerfeld, M.R.; Lu, C.; Hu, Q. Effect of initial biomass density on growth and astaxanthin production of Haematococcus pluvialis in an outdoor photo-bioreactor. J. Appl. Phycol. 2013, 25, 253–260. [Google Scholar] [CrossRef]
- Das, D. (Ed.) Algal Biorefinery: An Integrated Approach; Springer: Berlin, Germany, 2015. [Google Scholar]
- Hanan, N.A.; Al-Shorgani, N.K.; Shukor, H.; Abd. Rahman, N.; Kalil, M.S. Pre-Optimization Conditions for Haematococcus pluvialis Growth. Int. J. Adv. Sci. Eng. Inf. Technol. 2013, 3, 70–73. [Google Scholar] [CrossRef]
- Pham, K.T.; Nguyen, T.C.; Luong, T.H.; Dang, P.H.; Vu, D.C.; Do, T.N.; Phi, T.C.M.; Nguyen, D.B. Influence of inoculum size, CO2 concentration and LEDs on the growth of green microalgae Haematococcus pluvialis Flotow. Vietnam J. Sci. Technol. Eng. 2018, 60, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Liu, B.; Lu, X.; Cheng, K.W.; Chen, F. Staged cultivation enhances biomass accumulation in the green growth phase of Haematococcus pluvialis. Bioresour. Technol. 2017, 233, 326–331. [Google Scholar] [CrossRef]
- Witono, J.R.; Gunadi, A.; Santoso, H.; Miryanti, A.; Kumalaputri, A.J. The optimal condition on the growth of green Haematococcus pluvialis as one of the future natural resources. In Proceedings of the 26th Regional Symposium on Chemical Engineering (RSCE 2019), Kuala Lumpur, Malaysia, 30–31 October 2019. [Google Scholar]
- Li, J.; Zhu, D.; Niu, J.; Shen, S.; Wang, G. An economic assessment of astaxanthin production by large scale cultivation of Haematococcus pluvialis. Biotechnol. Adv. 2011, 29, 568–574. [Google Scholar] [CrossRef]
- Chandra, R.; Iqbal, H.M.N.; Vishal, G.; Hyung-Sool, L.; Sunil, N. Algal Biorefinery: A Sustainable Approach to Valorize Algal-based Biomass towards Multiple Product Recovery. Bioresour. Technol. 2019, 278, 346–359. [Google Scholar] [CrossRef]
- Nishshanka, G.K.S.H.; Liyanaarachchi, V.C.; Premaratne, M.; Nimarshana, P.H.V.; Ariyadasa, T.U.; Kornaros, M. Wastewater-based microalgal biorefineries for the production of astaxanthin and co-products: Current status, challenges and future perspectives. Bioresour Technol. 2021, 342, 126018. [Google Scholar] [CrossRef]
- Chew, K.W.; Yap, J.Y.; Show, P.L.; Suan, N.H.; Juan, J.C.; Ling, T.C.; Lee, D.; Chang, J. Microalgae biorefinery: High value products perspectives. Bioresour. Technol. 2017, 229, 53–62. [Google Scholar] [CrossRef]
- Fabio, R.; Paula, M.; Tatiana, M.; Paulo, C.A. Comparison of open-air and semi-enclosed cultivation system for massive microalgae production in sub-tropical and temperate latitudes. Biomass Bioenergy 2013, 59, 418–424. [Google Scholar]
- Gulzar, S.; Benjakul, S. Impact of pre-treatment and atmosphere on quality of lipids extracted from cephalothorax of Pacific white shrimp by ultrasonic assisted process. Food Chem. 2019, 309, 125732. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| System | Medium | Temperature (°C) | Light Intensities (μmol photon m−2 s−1) | Productivity | References |
|---|---|---|---|---|---|
| Closed Column Photobioreactors | BG−11 | 25 | 50, 100, 200, 400 | The highest level of astaxanthin production was achieved in nitrogen starvation and under light intensity at 200 μmol photons m−2 s−1, which productivity obtained 1.64 times higher than the control group under the photoautotrophic mode. | [3] |
| Closed air-lift photo-bioreactors | Bold’s Basal Medium | 23–25 | 190 | The system comprised of 90 dm3 volume photo-bioreactors was obtained biomass concentration in the range 1.40–1.99 g dm−3 on the eighth day of conducting experiments | [11] |
| Vertical bubble column photo-bioreactor (VBC-PBR) | BG−11 | 28 | 55–280 | High-intensity light stress increased total carotenoids (5.21 mg g−1) and FA production (19.62 mg g−1), while low light intensity stress decreased total carotenoids (5.21 mg g−1) under the different photoautotrophic conditions. | [12] |
| Outdoor raceway pond | Modified BG11 | Not Control | 300, 800, 1800 | The mixotrophic cultivation in an outdoor raceway pond of H. lacustris reached a maximum biomass productivity of 6.75 g m−2·day−1, the astaxanthin productivity reached 140 mg m−2·day−1 and about 1.2 times that of the simple phototrophic cultivation. | [14] |
| Twin-Layer porous substrate photo-bioreactor (TL-PSBR) | Modified Blue Green 11 (BG11) | 26 | 300–1000 | The TL-PSBR produced the most biomass and astaxanthin, at 8.7 g m−2 day−1 and 170 mg m−2 day−1, respectively. The longer storage times reduced productivity, but the most efficient use occurred between 300–500 μmol photon m−2·s−1. The experiment has been cultivated in the photoautotrophic mode. | [30] |
| Twin-Layer porous substrate photo-bioreactor (TL-PSBR) | Modified BG11 | 26 | 300 | In the 0.05 m2 and 2 m2 systems, dry biomass productivity reached 12 g m−2 day−1 (3% astaxanthin content in the dry biomass) and 11.25 g m−2 day−1 (2.8% astaxanthin) after 10 days of cultivation. The cultivation was conducted under photoautotrophic mode. | [38] |
| Small-scale angled (TL-PSBR) | Modified BG11 | 26 | 20–80 | The cultivation under photoautotrophic with initial biomass at 6.5 g·m−2 (inoculum) density of the biofilm was 6.5 g·m−2·day−1 dry biomass yield under 80 mol photon m−2·s−1 light intensity. | [39] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oslan, S.N.H.; Oslan, S.N.; Mohamad, R.; Tan, J.S.; Yusoff, A.H.; Matanjun, P.; Mokhtar, R.A.M.; Shapawi, R.; Huda, N. Bioprocess Strategy of Haematococcus lacustris for Biomass and Astaxanthin Production Keys to Commercialization: Perspective and Future Direction. Fermentation 2022, 8, 179. https://doi.org/10.3390/fermentation8040179
Oslan SNH, Oslan SN, Mohamad R, Tan JS, Yusoff AH, Matanjun P, Mokhtar RAM, Shapawi R, Huda N. Bioprocess Strategy of Haematococcus lacustris for Biomass and Astaxanthin Production Keys to Commercialization: Perspective and Future Direction. Fermentation. 2022; 8(4):179. https://doi.org/10.3390/fermentation8040179
Chicago/Turabian StyleOslan, Siti Nur Hazwani, Siti Nurbaya Oslan, Rosfarizan Mohamad, Joo Shun Tan, Abdul Hafidz Yusoff, Patricia Matanjun, Ruzaidi Azli Mohd Mokhtar, Rossita Shapawi, and Nurul Huda. 2022. "Bioprocess Strategy of Haematococcus lacustris for Biomass and Astaxanthin Production Keys to Commercialization: Perspective and Future Direction" Fermentation 8, no. 4: 179. https://doi.org/10.3390/fermentation8040179
APA StyleOslan, S. N. H., Oslan, S. N., Mohamad, R., Tan, J. S., Yusoff, A. H., Matanjun, P., Mokhtar, R. A. M., Shapawi, R., & Huda, N. (2022). Bioprocess Strategy of Haematococcus lacustris for Biomass and Astaxanthin Production Keys to Commercialization: Perspective and Future Direction. Fermentation, 8(4), 179. https://doi.org/10.3390/fermentation8040179